Protein Identification of Venoms of the African Spitting Cobras, Naja mossambica and Naja nigricincta nigricincta

 , , , , ,
, , , , , 
Abstract
:1. Introduction
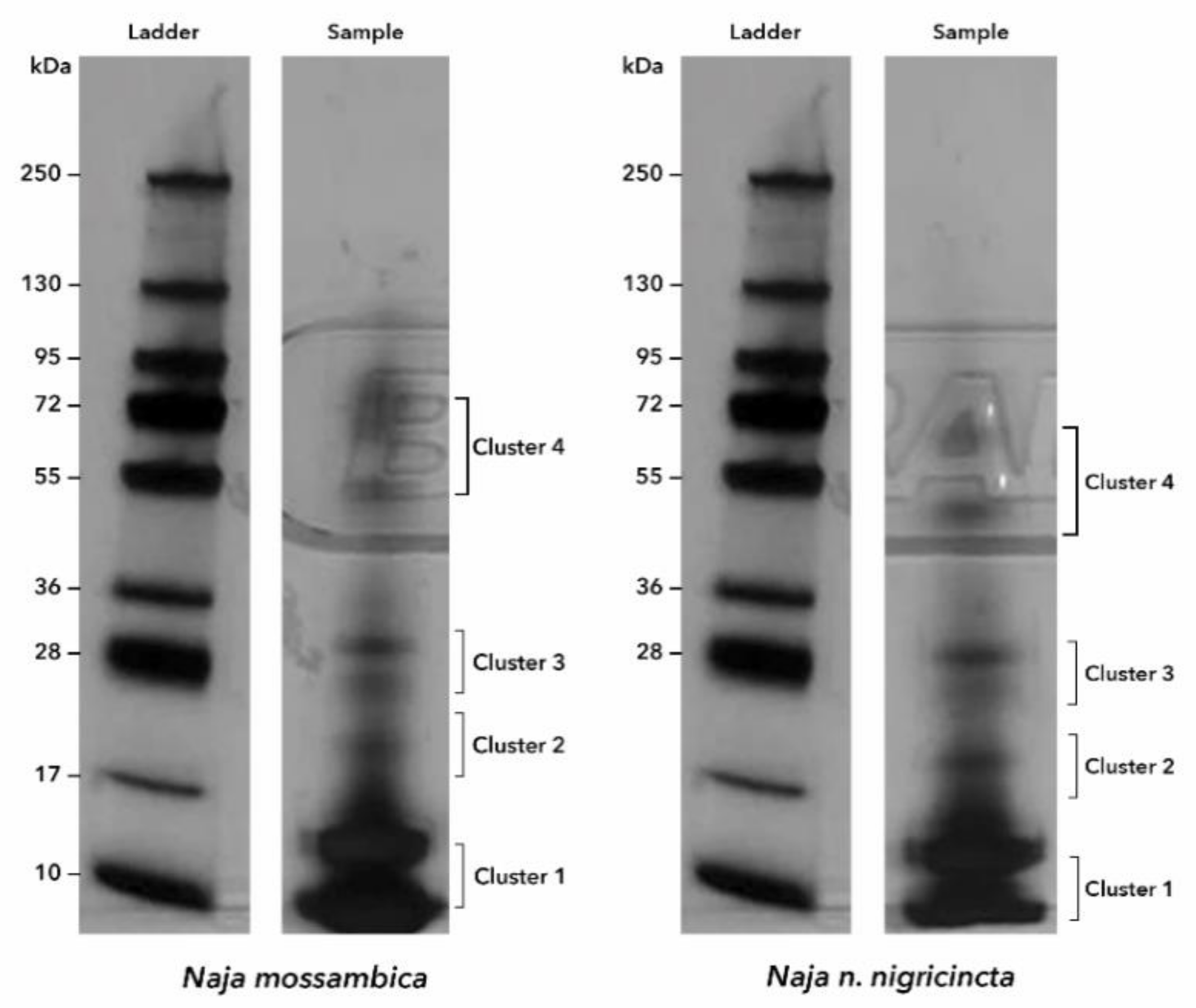
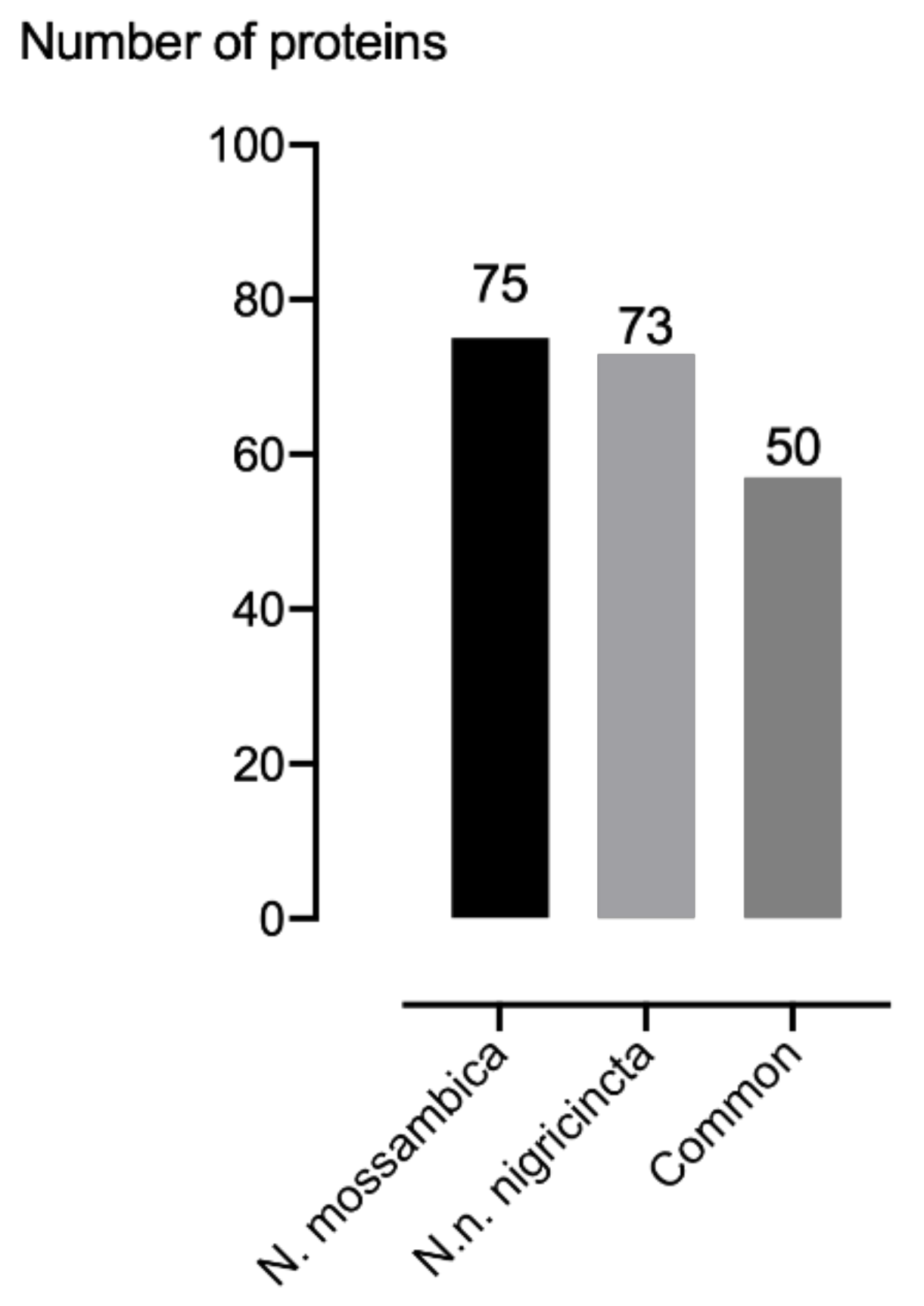
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Snake Venom Samples
5.2. Venom Protein Separation by SDS-PAGE and In-Gel Digestion
5.3. Acetonitrile Precipitation and In-Solution Digestion
5.4. LC-ESI-MS/MS Analysis
5.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Silva, J.; Kasturiratne, A.; Pathmeswaran, A.; Lalloo, D.G. Snakebite: The true disease burden has yet to be determined. Ceylon Med. J. 2013, 58, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Snakebite Envenoming. 2019. Available online: https://www.who.int/en/news-room/fact-sheets/detail/snakebite-envenoming (accessed on 7 August 2019).
- Habib, A.G.; Kuznik, A.; Hamza, M.; Abdullahi, M.I.; Chedi, B.A.; Chippaux, J.-P.; Warrell, D.A. Snakebite is Under Appreciated: Appraisal of Burden from West Africa. PLoS Negl. Trop. Dis. 2015, 9, e0004088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chippaux, J.-P. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Longbottom, J.; Shearer, F.M.; Devine, M.; Alcoba, G.; Chappuis, F.; Weiss, D.J.; Ray, S.E.; Ray, N.; Warrell, D.A.; De Castañeda, R.R.; et al. Vulnerability to snakebite envenoming: A global mapping of hotspots. Lancet 2018, 392, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Kasturiratne, A.; Wickremasinghe, A.R.; De Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; De Silva, H.J. The Global Burden of Snakebite: A Literature Analysis and Modelling Based on Regional Estimates of Envenoming and Deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef]
- Williams, D.J.; Faiz, M.A.; Abela-Ridder, B.; Ainsworth, S.; Bulfone, T.C.; Nickerson, A.D.; Habib, A.G.; Junghanss, T.; Fan, H.W.; Turner, M.; et al. Strategy for a globally coordinated response to a priority neglected tropical disease: Snakebite envenoming. PLoS Negl. Trop. Dis. 2019, 13, e0007059. [Google Scholar] [CrossRef] [Green Version]
- Hunter, C.J.; Piechazek, K.-H.; Nyarang’o, P.M.; Rennie, T.W. Snakebite envenoming. Lancet 2019, 393, 129–131. [Google Scholar] [CrossRef] [Green Version]
- Shine, R.; Branch, W.R.; Webb, J.K.; Harlow, P.S.; Shine, T.; Keogh, J.S. Ecology of cobras from southern Africa. J. Zool. 2007, 272, 183–193. [Google Scholar] [CrossRef]
- Wüster, W.; Crookes, S.; Ineich, I.; Mané, Y.; Pook, C.E.; Trape, J.-F.; Broadley, D.G. The phylogeny of cobras inferred from mitochondrial DNA sequences: Evolution of venom spitting and the phylogeography of the African spitting cobras (Serpentes: Elapidae: Naja nigricollis complex). Mol. Phylogenet. Evol. 2007, 45, 437–453. [Google Scholar] [CrossRef]
- World Health Organization. WHO Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; WHO: Geneva, Switzerland, 2010; p. 134. [Google Scholar]
- Panagides, N.; Jackson, T.N.; Ikonomopoulou, M.P.; Arbuckle, K.; Pretzler, R.; Yang, D.C.; Ali, S.A.; Koludarov, I.; Dobson, J.; Sanker, B.; et al. How the Cobra Got Its Flesh-Eating Venom: Cytotoxicity as a Defensive Innovation and Its Co-Evolution with Hooding, Aposematic Marking, and Spitting. Toxins 2017, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Kandiwa, E.; Mushonga, B.; Samkange, A.; Fabiano, E. Quantitative Characterization of the Hemorrhagic, Necrotic, Coagulation-Altering Properties and Edema-Forming Effects of Zebra Snake (Naja nigricincta nigricincta) Venom. J. Toxicol. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tianyi, F.-L.; Dimala, C.A.; Feteh, V.F. Shortcomings in snake bite management in rural Cameroon: A case report. BMC Res. Notes 2017, 10, 196. [Google Scholar] [CrossRef] [PubMed]
- Suleman, M.M.; Shahab, S.; Rab, M.A. Snake bite in the Thar Desert. J.-Pak. Med Assoc. 1998, 48, 306–307. [Google Scholar] [PubMed]
- Namibia Standard Treatment Guidelines, 1st ed.; Ministry of Health and Social Services: Windhoek, Namibia, 2011; p. 891.
- Sintiprungrat, K.; Watcharatanyatip, K.; Senevirathne, W.; Chaisuriya, P.; Chokchaichamnankit, D.; Srisomsap, C.; Ratanabanangkoon, K. A comparative study of venomics of Naja naja from India and Sri Lanka, clinical manifestations and antivenomics of an Indian polyspecific antivenom. J. Proteom. 2016, 132, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.A.; Oluoch, G.O.; Ainsworth, S.; Alsolaiss, J.; Bolton, F.; Arias, A.-S.; Gutiérrez, J.-M.; Rowley, P.; Kalya, S.; Ozwara, H.; et al. Preclinical antivenom-efficacy testing reveals potentially disturbing deficiencies of snakebite treatment capability in East Africa. PLoS Negl. Trop. Dis. 2017, 11, e0005969. [Google Scholar] [CrossRef] [Green Version]
- Tilbury, C.R. Observations on the bite of the Mozambique spitting cobra (Naja mossambica mossambica). S. Afr. Med. J. 1982, 61, 308–313. [Google Scholar]
- Laustsen, A.H.; Lomonte, B.; Lohse, B.; Fernandez, J.; Gutiérrez, J.M. Unveiling the nature of black mamba (Dendroaspis polylepis) venom through venomics and antivenom immunoprofiling: Identification of key toxin targets for antivenom development. J. Proteom. 2015, 119, 126–142. [Google Scholar] [CrossRef]
- Pantanowitz, L.L. Development of antivenoms in South Africa. S. Afr. J. Sci. 1998, 94, 464–469. [Google Scholar]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [Green Version]
- Petras, D.; Sanz, L.; Segura, A.; Herrera, M.; Villalta, M.; Solano, D.; Vargas, M.; León, G.; Warrell, D.A.; Theakston, R.D.G.; et al. Snake Venomics of African Spitting Cobras: Toxin Composition and Assessment of Congeneric Cross-Reactivity of the Pan-African EchiTAb-Plus-ICP Antivenom by Antivenomics and Neutralization Approaches. J. Proteome Res. 2011, 10, 1266–1280. [Google Scholar] [CrossRef]
- Huang, M.Z.; Gopalakrishnakone, P.; Kini, R.M.; Chung, M.C. Complete Amino Acid Sequence of an Acidic, Cardiotoxic Phospholipase A2 from the Venom of Ophiophagus Hannah (King Cobra): A Novel Cobra Venom Enzyme with “Pancreatic Loop”. Arch. Biochem. Biophys. 1997, 338, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 structure/function, mechanism, and signaling1. J. Lipid Res. 2008, 50, S237–S242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kini, R.M.; Doley, R. Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets. Toxicon 2010, 56, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Girish, V.M.; Kumar, S.; Joseph, L.; Jobichen, C.; Kini, R.M.; Sivaraman, J. Identification and Structural Characterization of a New Three-Finger Toxin Hemachatoxin from Hemachatus haemachatus Venom. PLoS ONE 2012, 7, e48112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munawar, A.; Trusch, M.; Georgieva, D.; Hildebrand, D.; Kwiatkowski, M.; Behnken, H.; Harder, S.; Arni, R.; Spencer, P.J.; Schluter, H.; et al. Elapid Snake Venom Analyses Show the Specificity of the Peptide Composition at the Level of Genera Naja and Notechis. Toxins 2014, 6, 850–868. [Google Scholar] [CrossRef] [Green Version]
- Mackessy, S.P. Handbook of Venoms and Toxins of Reptiles; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2016; Available online: https://owlcation.com/stem/Snake-Venom-Composition (accessed on 7 August 2019).
- Mayr, C. Evolution and Biological Roles of Alternative 3′UTRs. Trends Cell Biol. 2016, 26, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2012, 1824, 164–176. [Google Scholar] [CrossRef]
- Aird, S.D.; Watanabe, Y.; Villar-Briones, A.; Roy, M.C.; Terada, K.; Mikheyev, A.S. Quantitative high-throughput profiling of snake venom gland transcriptomes and proteomes (Ovophis okinavensis and Protobothrops flavoviridis). BMC Genom. 2013, 14, 790. [Google Scholar] [CrossRef] [Green Version]
- Lahiani, A.; Yavin, E.; Lazarovici, P. The Molecular Basis of Toxins’ Interactions with Intracellular Signaling via Discrete Portals. Toxins 2017, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.K.; Bay, B.H.; Gopalakrishnakone, P. L-amino acid oxidase from snake venom and its anticancer potential. Toxicon 2018, 144, 7–13. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Brown, R.L.; Morita, T. Purification and Cloning of Toxins from Elapid Venoms that Target Cyclic Nucleotide-Gated Ion Channels. Biochemistry 2002, 41, 11331–11337. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Structure and function of snake venom cysteine-rich secretory proteins. Toxicon 2004, 44, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Adade, C.M.; Carvalho, A.L.O.; Tomaz, M.A.; Costa, T.F.R.; Godinho, J.L.; Melo, P.A.; Lima, A.P.C.A.; Rodrigues, J.C.F.; Zingali, R.B.; Souto-Padrón, T. Crovirin, a Snake Venom Cysteine-Rich Secretory Protein (CRISP) with Promising Activity against Trypanosomes and Leishmania. PLoS Negl. Trop. Dis. 2014, 8, e3252. [Google Scholar] [CrossRef] [PubMed]
- Schweitz, H.; Vigne, P.; Moinier, D.; Frelin, C.; Lazdunski, M. A new member of the natriuretic peptide family is present in the venom of the green mamba (Dendroaspis angusticeps). J. Biol. Chem. 1992, 267, 13928–13932. [Google Scholar]
- Chen, H.H.; Lainchbury, J.G.; Burnett, J.C. Natriuretic peptide receptors and neutral endopeptidase in mediating the renal actions of a new therapeutic synthetic natriuretic peptide dendroaspis natriuretic peptide. J. Am. Coll. Cardiol. 2002, 40, 1186–1191. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-S.; Rejtar, T.; Andreev, V.; Moskovets, A.E.; Karger, B.L. Enhanced Characterization of Complex Proteomic Samples Using LC−MALDI MS/MS: Exclusion of Redundant Peptides from MS/MS Analysis in Replicate Runs. Anal. Chem. 2005, 77, 7816–7825. [Google Scholar] [CrossRef]
- Liu, H.; Sadygov, R.G.; Yates, I.J.R. A Model for Random Sampling and Estimation of Relative Protein Abundance in Shotgun Proteomics. Anal. Chem. 2004, 76, 4193–4201. [Google Scholar] [CrossRef]
- Ainsworth, S.; Petras, D.; Engmark, M.; Süssmuth, R.D.; Whiteley, G.; Albulescu, L.-O.; Kazandjian, T.D.; Wagstaff, S.; Rowley, P.; Wüster, W.; et al. The medical threat of mamba envenoming in sub-Saharan Africa revealed by genus-wide analysis of venom composition, toxicity and antivenomics profiling of available antivenoms. J. Proteom. 2018, 172, 173–189. [Google Scholar] [CrossRef]
- Saaiman, E.L.; Buys, P.J.C. Spitting cobra (Naja nigricincta nigricincta) bites complicated by rhabdomyolysis, possible intravascular haemolysis, and coagulopathy. S. Afr. Med. J. 2019, 109, 736–740. [Google Scholar] [CrossRef]
- O’Connell, K.L.; Stults, J.T. Identification of mouse liver proteins on two-dimensional electrophoresis gels by matrix-assisted laser desorption/ionization mass spectrometry of in situ enzymatic digests. Electrophoresis 1997, 18, 349–359. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass Spectrometric Sequencing of Proteins from Silver-Stained Polyacrylamide Gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Llinares, M.B.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2018, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Protein Family | Short Description | Mode of Action/Function | MW (Kda) | References |
|---|---|---|---|---|
| PLA2 | Esterases with a unique pancreatic loop at 62–66 residues. | Cleaves the sn-2 acyl bond to hydrolyse phospholipids to release fatty acid and lyso-phospholipid. | 13–15 | [24,25] |
| 3FTxs | Non-enzymatic polypeptides consisting of 60–74 amino acids, of which three peptide loops are stabilised by four or five disulphide bridges. | Mainly function as potent inhibitors of neuromuscular transmission, cardiac function and acetylcholinesterase. Moreover, interfere with nicotinic and muscarinic acetylcholine receptors, acetylcholinesterase, ion channels, and cell membranes. | 6–9 | [26,27,28,29,30] |
| SVMPs | Monozinc endopeptidases. | Not well understood, but in general, they induce haemorrhage, inflammation, and apoptosis, while inhibiting aggregation of platelets. | 20–100 | [31,32] |
| KUN | Dendrotoxins consisting of 57–60 amino acids organised in a single polypeptide chain cross-linked by three disulfidebridges. | Facilitate the release of the neurotransmitter acetylcholine at the neuromuscular junction, resulting in blockage of certain voltage-dependent K+ channels in the nerve presynaptic terminals. | [33] | |
| LAAO | Flavoenzymes found in acidic, neutral and basic forms. | Not well understood but exist as homodimers which contribute the cytotoxic activities. | 50–70 | [34] |
| CRiSP | Single chain polypeptides containing 16 cysteine molecules, of which ten are clustered in the C-terminal of the sequence. | Act as inhibitors of cyclic nucleotide-gated channels and L-type Ca2+ and/or K+ channels, to prevent smooth muscle contraction. | 20–30 | [35,36,37] |
| NPs | 38 residue peptides with four proline residues, of which three are clustered in the C terminal of the molecule. | Diffuse into vascular smooth muscle cells to activate guanylate cyclase inducing vasodilation. | – | [38,39] |
| Method Approach | Number of Proteins Per Species | |
|---|---|---|
| N. mossambica n (%) | N. n. nigricintca n (%) | |
| In-gel digestion (Silver-stained) | 71 (94.7) | 68 (93.2) |
| In-gel digestion (Not stained) | 54 (72.0) | 58 (79.5) |
| In solution digestion | 32 (42.7) | 22 (30.1) |
| No digestion | 4 (5.3) | 3 (4.1) |
| Protein Family | Number of Proteins | ||
|---|---|---|---|
| N. mossambica | N. n. nigricintca | Common to Both Snakes | |
| 5’-nucleotidase | 0 | 1 | 1 |
| Cathelicidin | 2 | 0 | 0 |
| Cystatin | 1 | 0 | 1 |
| Cysteine-rich secretory protein (CRiSP) | 1 | 3 | 6 |
| Flavin monoamine oxidase | 1 | 1 | 4 |
| Glycosyl hydrolase 56 | 0 | 2 | 0 |
| Nerve growth factor (NGF-Beta) | 0 | 1 | 3 |
| Nucleotide pyrophosphate/phosphodiesterase | 0 | 0 | 1 |
| Ohanin/vespryn | 0 | 0 | 1 |
| Peptidase S1 | 0 | 0 | 1 |
| Peroxiredoxin | 0 | 1 | 0 |
| Phospholipase A2 (PLA2) | 1 | 1 | 6 |
| Phospholipase B | 0 | 0 | 2 |
| Three finger toxins (3FTx) | 10 | 5 | 20 |
| True venom lectin | 0 | 0 | 1 |
| Venom complement C3 homolog | 0 | 0 | 3 |
| Venom Kunitz-type family | 1 | 0 | 0 |
| Venom metalloproteinase M12B (SVMP) | 1 | 1 | 7 |
| Total | 18 | 16 | 57 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katali, O.; Shipingana, L.; Nyarangó, P.; Pääkkönen, M.; Haindongo, E.; Rennie, T.; James, P.; Eriksson, J.; Hunter, C.J. Protein Identification of Venoms of the African Spitting Cobras, Naja mossambica and Naja nigricincta nigricincta. Toxins 2020, 12, 520. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080520
Katali O, Shipingana L, Nyarangó P, Pääkkönen M, Haindongo E, Rennie T, James P, Eriksson J, Hunter CJ. Protein Identification of Venoms of the African Spitting Cobras, Naja mossambica and Naja nigricincta nigricincta. Toxins. 2020; 12(8):520. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080520
Chicago/Turabian StyleKatali, Ottilie, Loide Shipingana, Peter Nyarangó, Mirva Pääkkönen, Erastus Haindongo, Timothy Rennie, Peter James, John Eriksson, and Christian John Hunter. 2020. "Protein Identification of Venoms of the African Spitting Cobras, Naja mossambica and Naja nigricincta nigricincta" Toxins 12, no. 8: 520. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080520