Bioaccumulation and Phytotoxicity and Human Health Risk from Microcystin-LR under Various Treatments: A Pot Study


Abstract
:1. Introduction
2. Results and Discussion
2.1. Microcystin-LR (MC-LR) Bioaccumulation in Vegetables under Various Treatments
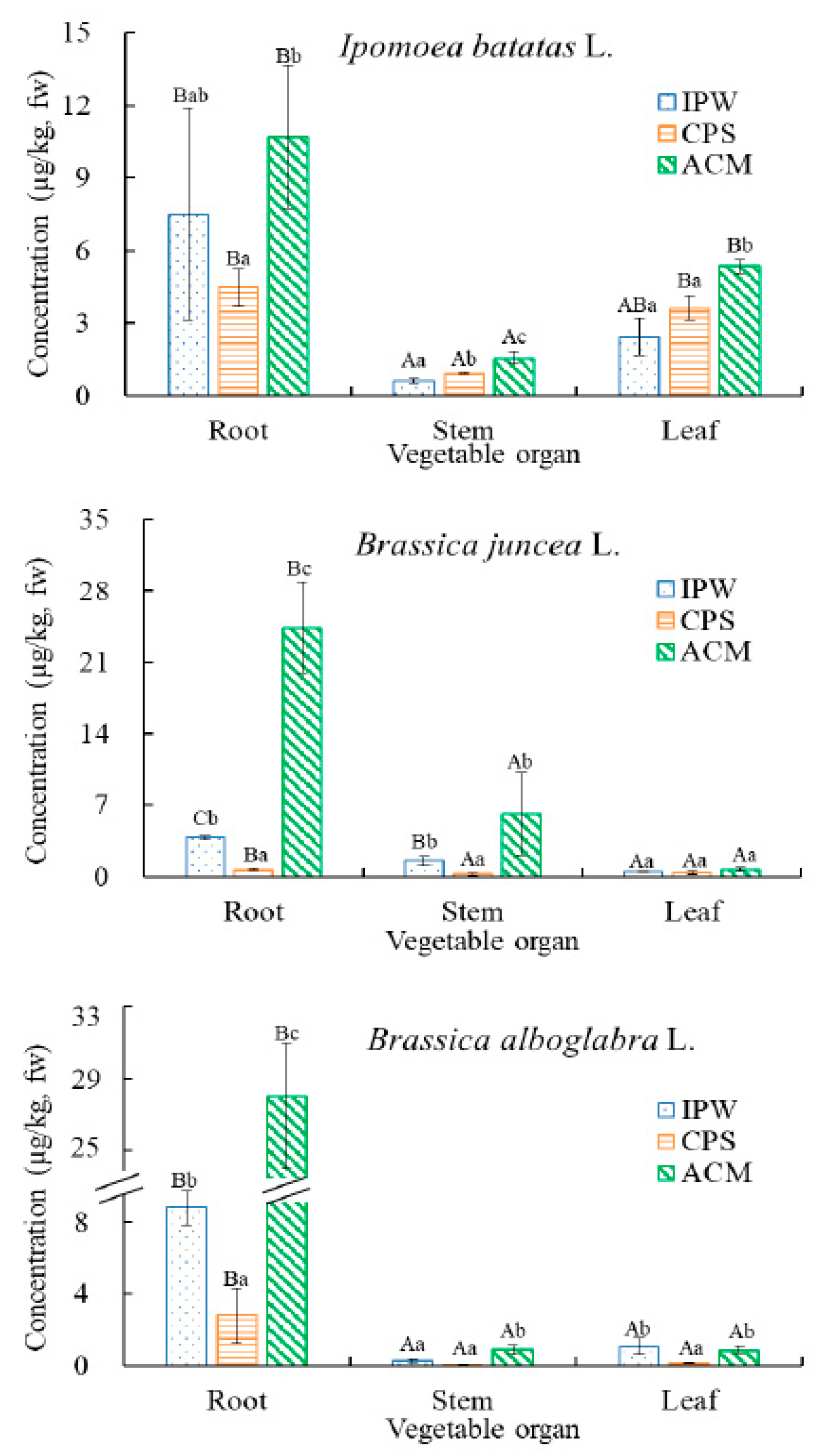
2.2. Uptake and Transfer of MC-LR in Vegetables under Various Treatments
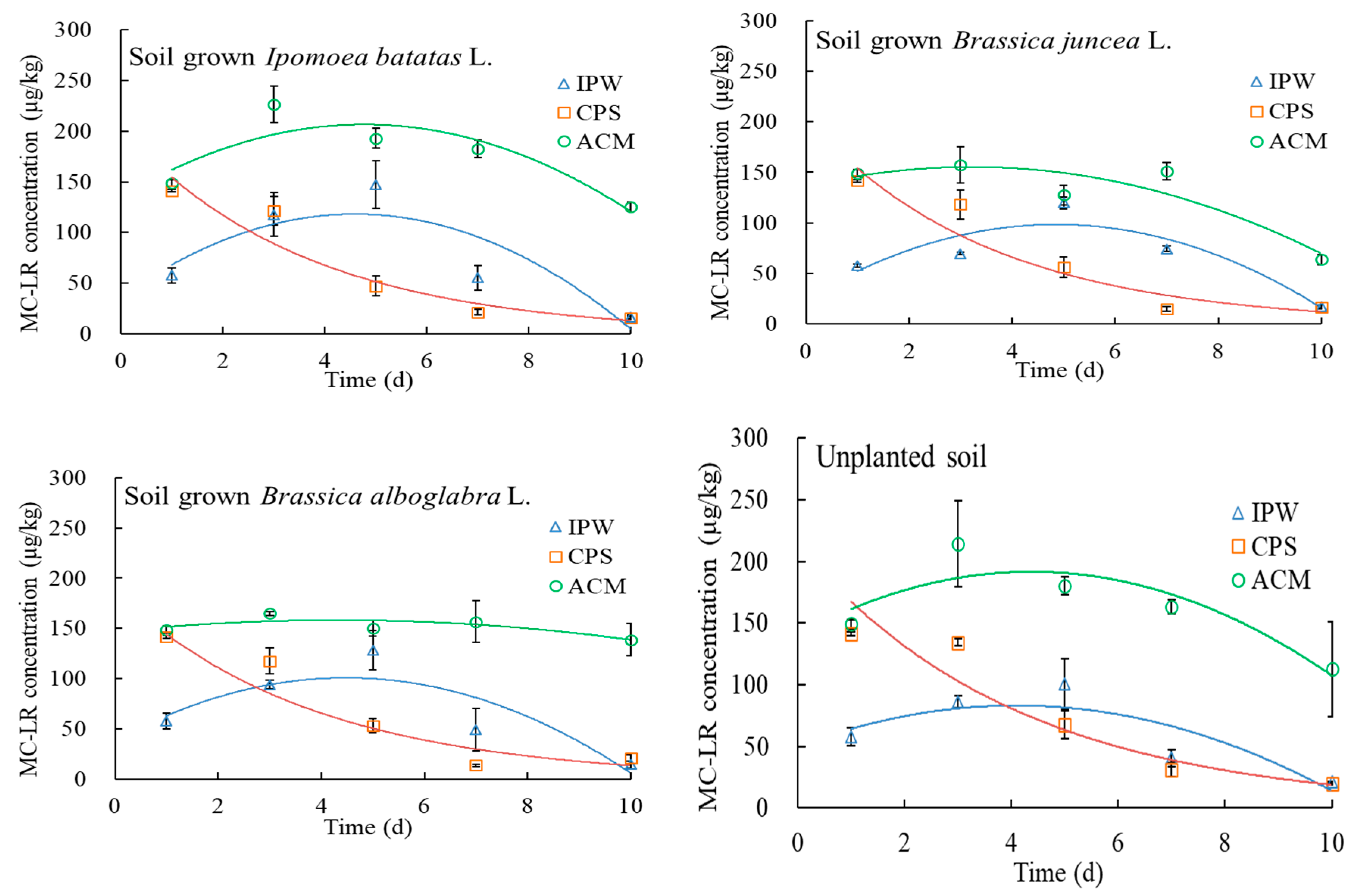
2.3. MC-LR Degradation in Soils under Various Treatments
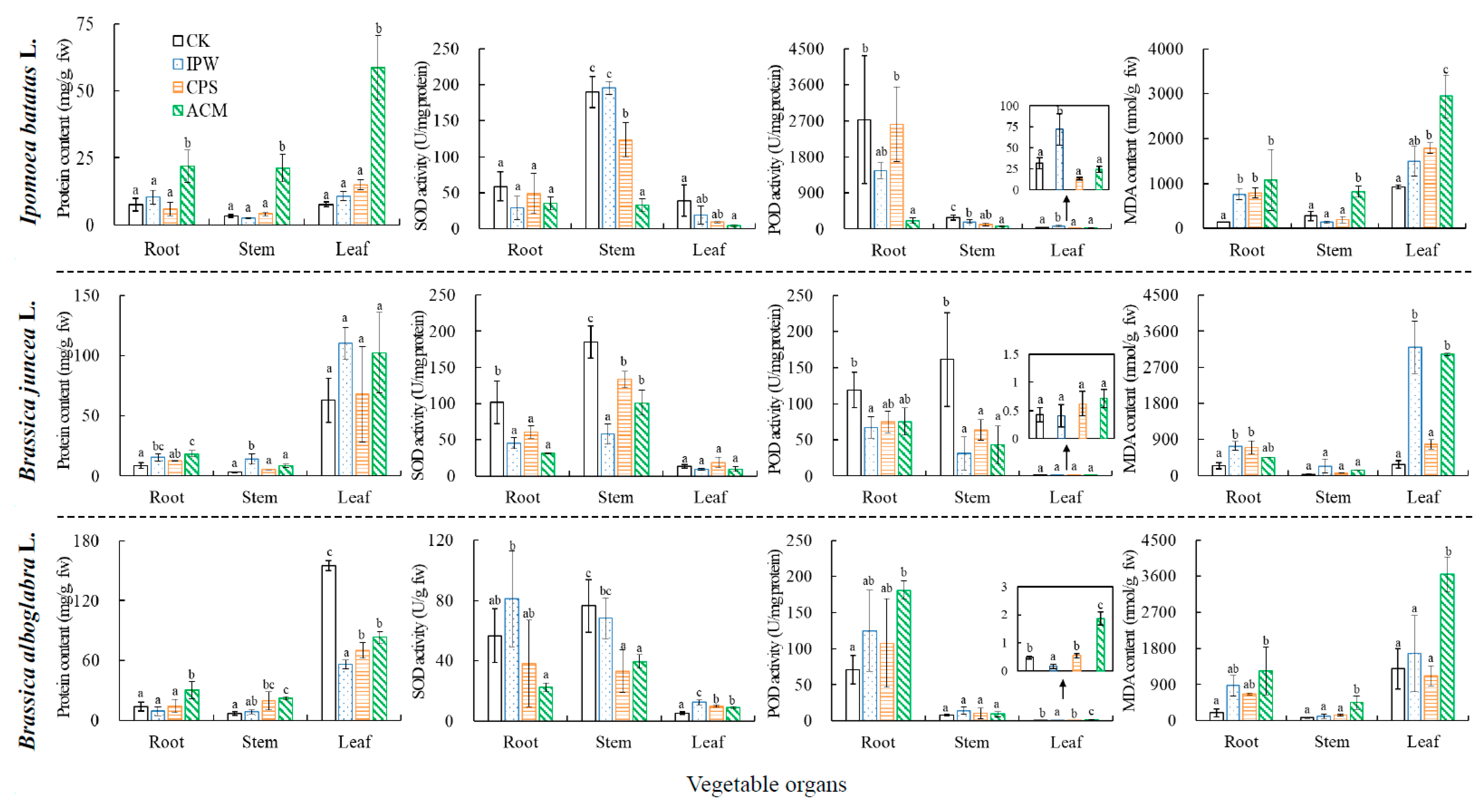
2.4. Toxicity to Vegetables from MC-LR under Various Treatments
2.5. Health Risk from Consuming the Vegetables under Various Treatments
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Pot Design
4.2.2. Extraction and Analysis of MC-LR in Soil and Vegetable Samples
4.2.3. Determination of Antioxidant Enzymes and MDA Content
4.2.4. Data Processing
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bolotaolo, M.; Kurobe, T.; Puschner, B.; Hammock, B.G.; Hengel, M.J.; Lesmeister, S.; Teh, S.J. Analysis of covalently bound microcystins in sediments and clam tissue in the sacramento–San Joaquin River Delta, California, USA. Toxins 2020, 12, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Gu, P.; Zhang, C.; Luo, X.; Zhang, H.; Zhang, J.; Zheng, Z. Combined toxic effects of anatoxin-a and microcystin-LR on submerged macrophytes and biofilms. J. Hazard. Mater. 2020, 389, 122053. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ersan, M.S.; Plewa, M.J.; Amy, G.; Karanfil, T. Formation of iodinated trihalomethanes and noniodinated disinfection byproducts during chloramination of algal organic matter extracted from Microcystis aeruginosa. Water Res. 2019, 162, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Wan, X.; Shu, X.; Xie, L. Bioaccumulation and detoxication of microcystin-LR in three submerged macrophytes: The important role of glutathione biosynthesis. Chemosphere 2019, 225, 935–942. [Google Scholar] [CrossRef]
- Catherine, Q.; Susanna, W.; Isidora, E.S.; Mark, H.; Aurelie, V.; Jeanfrançois, H. A review of current knowledge on toxic benthic freshwater cyanobacteria ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef]
- Liu, C.; Ersan, M.S.; Plewa, M.J.; Amy, G.; Karanfil, T. Formation of regulated and unregulated disinfection byproducts during chlorination of algal organic matter extracted from freshwater and marine algae. Water Res. 2018, 142, 313–324. [Google Scholar] [CrossRef]
- Freitas, M.; Azevedo, J.; Pinto, E.; Neves, J.; Campos, A.; Vasconcelos, V. Effects of microcystin-LR, cylindrospermopsin and a microcystin-LR/cylindrospermopsin mixture on growth, oxidative stress and mineral content in lettuce plants (Lactuca sativa L.). Ecotoxicol. Environ. Saf. 2015, 116, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Preece, E.P.; Hardy, F.J.; Moore, B.C.; Bryan, M. A review of microcystin detections in estuarine and marine waters: Environmental implications and human health risk. Harmful Algae 2017, 61, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Ufelmann, H.; Krüger, T.; Luckas, B.; Schrenk, D. Human and rat hepatocyte toxicity and protein phosphatase 1 and 2A inhibitory activity of naturally occurring desmethyl-microcystins and nodularins. Toxicology 2012, 293, 59–67. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Ingested Nitrate and Nitrite and Cyanobacterial Peptide Toxins: IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, 94th ed. Available online: https://publications.iarc.fr/Book-And-Report-Series/Iarc-Monographs-On-The-Identification-Of-Carcinogenic-Hazards-To-Humans/Ingested-Nitrate-And-Nitrite-And-Cyanobacterial-Peptide-Toxins-2010 (accessed on 14 August 2020).
- Zhou, L.; Chen, K. Relationship between microcystin in drinking water and colorectal cancer. Biomed. Environ. Sci. 2002, 15, 166–171. [Google Scholar] [CrossRef]
- Xiang, L.; Li, Y.-W.; Liu, B.-L.; Zhao, H.-M.; Li, H.; Cai, Q.-Y.; Mo, C.-H.; Wong, M.-H.; Li, Q.X. High ecological and human health risks from microcystins in vegetable fields in southern China. Environ. Int. 2019, 133, 105142. [Google Scholar] [CrossRef]
- Chen, W.; Song, L.; Gan, N.; Li, L. Sorption, degradation and mobility of microcystins in Chinese agriculture soils: Risk assessment for groundwater protection. Environ. Pollut. 2006, 144, 752–758. [Google Scholar] [CrossRef]
- Liu, H.; Guo, X.; Liu, L.; Yan, M.; Li, J.; Hou, S.; Wan, J.; Feng, L.L. Simultaneous Microcystin Degradation and Microcystis Aeruginosa Inhibition with Single Enzyme Microcystinase A. Environ. Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Yang, F.; Huang, F.; Feng, H.; Wei, J.; Massey, I.Y.; Liang, G.; Zhang, F.; Yin, L.; Kacew, S.; Zhang, X.; et al. A complete route for biodegradation of potentially carcinogenic cyanotoxin microcystin-LR in a novel indigenous bacterium. Water Res. 2020, 174, 115638. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Steinman, A.D.; Wan, X.; Xie, L. Bioaccumulation of microcystin congeners in soil-plant system and human health risk assessment: A field study from Lake Taihu region of China. Environ. Pollut. 2018, 240, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Jiang, X.; Manubolu, M.; Riedl, K.; Ludsin, S.A.; Martin, J.F.; Lee, J. Fresh produce and their soils accumulate cyanotoxins from irrigation water: Implications for public health and food security. Food Res. Int. 2017, 102, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Song, L.; Dai, J.; Gan, N.; Liu, Z. Effects of microcystins on the growth and the activity of superoxide dismutase and peroxidade of rape (Brassica napus L.) and rice (Oryza sativa L.). Toxicon 2004, 43, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Steinman, A.D.; Wan, X.; Xie, L. Combined toxicity of microcystin-LR and copper on lettuce (Lactuca sativa L.). Chemosphere 2018, 206, 474–482. [Google Scholar] [CrossRef]
- Cordeiro-Araújo, M.K.; Chia, M.A.; de Toledo Arruda-Neto, J.D.; Tornisielo, V.L.; Vilca, F.Z.; do Carmo Bittencourt-Oliveira, M. Microcystin-LR bioaccumulation and depuration kinetics in lettuce and arugula: Human health risk assessment. Sci. Total Environ. 2016, 566–567, 1379–1386. [Google Scholar] [CrossRef]
- Wen, H.-F. Toxicological effects of microcystin-LR on earthworm (Eisenia fetida) in soil. Biol. Fertil. Soils 2017, 53, 849–860. [Google Scholar] [CrossRef]
- Ding, Q.; Liu, K.; Song, Z.; Sun, R.; Zhang, J.; Yin, L.; Pu, Y. Effects of microcystin-LR on metabolic functions and structure succession of sediment bacterial community under anaerobic conditions. Toxins 2020, 12, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Zhu, L.; Fang, S.; Wang, C.; Shan, G. Photocatalytic degradation efficiency and mechanism of microcystin-RR by mesoporous Bi2WO6 under near ultraviolet light. Environ. Sci. Technol. 2012, 46, 2345–2351. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.; Campos, A.; Vasconcelos, V.; Freitas, M. Effects of microcystin-LR and cylindrospermopsin on plant-soil systems: A review of their relevance for agricultural plant quality and public health. Environ. Res. 2017, 153, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Wang, C.; Tian, C.; Xiao, B.; Song, L. Evaluation of the potential of anoxic biodegradation of intracellular and dissolved microcystins in lake sediments. J. Hazard. Mater. 2015, 286, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Liu, W.; Jiang, W.; Shu, X.; Xie, L. Glutathione biosynthesis plays an important role in microcystin-LR depuration in lettuce and spinach. Environ. Pollut. 2019, 253, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Corbel, S.; Bouaïcha, N.; Nélieu, S.; Mougin, C. Soil irrigation with water and toxic cyanobacterial microcystins accelerates tomato development. Environ. Chem. Lett. 2015, 13, 447–452. [Google Scholar] [CrossRef]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial toxins: Modes of actions, fate in aquatic and soil ecosystems, phytotoxicity and bioaccumulation in agricultural crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef]
- Chen, S.; Ma, Z.; Li, S.; Waigi, M.G.; Jiang, J.; Liu, J.; Ling, W. Colonization of polycyclic aromatic hydrocarbon-degrading bacteria on roots reduces the risk of PAH contamination in vegetables. Environ. Int. 2019, 132, 105081. [Google Scholar] [CrossRef]
- Wang, J.; Liu, J.; Ling, W.; Huang, Q.; Gao, Y. Composite of PAH-degrading endophytic bacteria reduces contamination and health risks caused by PAHs in vegetables. Sci. Total Environ. 2017, 598, 471–478. [Google Scholar] [CrossRef]
- Berg, K.A.; Lyra, C.; Sivonen, K.; Paulin, L.; Suomalainen, S.; Tuomi, P.; Rapala, J. High diversity of cultivable heterotrophic bacteria in association with cyanobacterial water blooms. ISME J. 2009, 3, 314–325. [Google Scholar] [CrossRef]
- Ozaki, K.; Ohta, A.; Iwata, C.; Horikawa, A.; Tsuji, K.; Ito, E.; Ikai, Y.; Harada, K. Lysis of cyanobacteria with volatile organic compounds. Chemosphere 2008, 71, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Maghsoudi, E.; Fortin, N.; Greer, C.; Maynard, C.; Page, A.; Duy, S.V.; Sauve, S.; Prevost, M.; Dorner, S. Cyanotoxin degradation activity and mlr gene expression profiles of a Sphingopyxis sp. isolated from Lake Champlain, Canada. Environ. Sci. Process. Impacts 2016, 18, 1417–1426. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Ai, H.; Kang, L.; Sun, X.; He, Q. Simultaneous microcystis algicidal and microcystin degrading capability by a single Acinetobacter bacterial strain. Environ. Sci. Technol. 2016, 50, 11903–11911. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Han, F.X.; Zhang, H.Q.; Shi, Z.Q. Accumulation and phytotoxicity of microcystin-LR in rice (Oryza sativa). Ecotoxicol. Environ. Saf. 2012, 76, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.Y.; Xiang, L.; Wu, X.L.; Jiang, Y.N.; Li, Y.-W.; Mo, C.-H.; Cai, Q.-Y.; Huang, X.P.; Zhao, H.-M. Difference of anti-oxidation activities in Chinese flowering cabbages (Brassica campestris L.) with high and low accumulation of ciprofloxacin. Acta Sci. Circumstantiae 2016, 36, 2694–2700. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Wiegand, C.; Beattie, K.A.; Krause, E.; Steinberg, C.E.W.; Codd, G.A. Uptake, effects, and metabolism of cyanobacterial toxins in the emergent reed plant Phragmites australis (Cav.) Trin. Ex steud. Environ. Toxicol. Chem. 2001, 20, 846–852. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Wiegand, C.; Oberemm, A.; Beattie, K.A.; Krause, E.; Codd, G.A.; Steinberg, C.E.W. Identification of an enzymatically formed glutathione conjugate of thecyanobacterial hepatotoxin microcystin-LR: The first step of detoxication. Biochim. Biophys. Acta 1998, 1425, 527–533. [Google Scholar] [CrossRef]
- Vuolo, M.M.; Lima1, V.S.; Junior, M.R.M. Chapter 2—Phenolic Compounds: Structure, Classification, and Antioxidant Power, Bioactive Compounds; Woodhead Publishing: Sawston, UK, 2019; pp. 33–50. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar] [CrossRef]
- Wu, X.L.; Xiang, L.; Yan, Q.Y.; Jiang, Y.N.; Li, Y.-W.; Huang, X.P.; Li, H.; Cai, Q.-Y.; Mo, C.-H. Distribution and risk assessment of quinolone antibiotics in the soils from organic vegetable farms of a subtropical city, southern China. Sci. Total Environ. 2014, 487, 399–406. [Google Scholar] [CrossRef]
- Hitzfeld, B.C.; Höger, S.J.; Dietrich, D.R. Cyanobacterial toxins: Removal during drinking water treatment, and human risk assessment. Environ. Health Perspect. 2000, 108, S113–S122. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.K. Analytical Methods of Soil and Agri-Chemistry; Scientific and Technology Press: Beijing, China, 2000. [Google Scholar]
- Xiang, L.; Chen, L.; Yu, L.-Y.; Yu, P.-F.; Zhao, H.-M.; Mo, C.-H.; Li, Y.-W.; Li, H.; Cai, Q.-Y.; Zhou, D.-M.; et al. Genotypic variation and mechanism in uptake and translocation of perfluorooctanoic acid (PFOA) in lettuce (Lactuca sativa L.) cultivars grown in PFOA-polluted soils. Sci. Total Environ. 2018, 636, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-W.; Zhan, X.-J.; Xiang, L.; Deng, Z.-S.; Huang, B.-H.; Wen, H.-F.; Sun, T.-F.; Cai, Q.-Y.; Li, H.; Mo, C.-H. Analysis of Trace Microcystins in Vegetables Using Solid-Phase Extraction Followed by High Performance Liquid Chromatography Triple-Quadrupole Mass Spectrometry. J. Agric. Food Chem. 2014, 62, 11831–11839. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-W.; Huang, X.P.; Wu, X.L.; Xiang, L.; Zhan, X.-J.; Li, Z.J.; Wen, H.-F.; Zhong, F.L.; Mo, C.-H.; Hong, A.H. Simultaneous extraction and determination of three microcystins from soil using solid phase extraction and liquid chromatography-tandem mass spectrometry. Chin. J. Anal. Chem. 2013, 41, 88–92. [Google Scholar] [CrossRef]
- Zhao, H.-M.; Du, H.; Xiang, L.; Li, Y.-W.; Li, H.; Cai, Q.-Y.; Mo, C.-H.; Cao, G.; Wong, M.-H. Physiological differences in response to di-n-butyl phthalate (DBP) exposure between low- and high-DBP accumulating cultivars of Chinese flowering cabbage (Brassica parachinensis L.). Environ. Pollut. 2016, 208, 840–849. [Google Scholar] [CrossRef]
- Hao, Z.B.; Jing, C.; Xu, Z. Plant Physiology Experiment; Harbin Institute of Technology Press: Harbin, China, 2004. [Google Scholar]
- Ministry of Environmental Protection of China. Exposure Factors Handbook of Chinese Population; China Environmental Science Press: Beijing, China, 2013.
- Luo, D.; Mu, T.H.; Sun, H.N.; Chen, J.W. Optimization of the formula and processing of a sweet potato leaf powder-based beverage. Food Sci. Nutr. 2019, 8, 2680–2691. [Google Scholar] [CrossRef]
- Ramasamy, S.; Sotelo, P.; Lin, M.Y.; Heng, C.H.; Kang, S.; Sarika, S. Validation of a bio-based integrated pest management package for the control of major insect pests on Chinese mustard in Cambodia. Crop Prot. 2020, 135, 104728. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Vegetable | Treatment | RCF | RTF | STF |
|---|---|---|---|---|
| Ipomoea batatas L. | IPW | 0.05 ± 0.02 Aab | 0.12 ± 0.08 Ba | 4.07 ± 1.54 Ba a |
| CPS | 0.03 ± 0.01 Ba | 0.21 ± 0.02 Ca | 3.94 ± 0.39 Ba | |
| ACM | 0.07 ± 0.02 Ab | 0.15 ± 0.03 Ba | 2.87 ± 1.20 Ba | |
| Brassica juncea L. | IPW | 0.03 ± 0.01 Ab | 0.41 ± 0.11 Ca | 0.31 ± 0.11Aa |
| CPS | 0.004 ± 0.001 Aa | 0.36 ± 0.07 Ba | 1.90 ± 1.12 Ab | |
| ACM | 0.16 ± 0.02 Bc | 0.25 ± 0.12 Ba | 0.17 ± 0.01Aa | |
| Brassica alboglabra L. | IPW | 0.06 ± 0.01 Ab | 0.03 ± 0.01 Aa | 2.62 ± 1.80 Ba |
| CPS | 0.02 ± 0.01 Ba | 0.02 ± 0.01 Aa | 2.73 ± 0.80 ABa | |
| ACM | 0.19 ± 0.02 Bc | 0.03 ± 0.01 Aa | 1.03 ± 0.45 ABa |
| Vegetable | Treatment | Equation a | R2b | Half-life (t1/2, d) c |
|---|---|---|---|---|
| Ipomoea batatas L. | IPW | y = −3.9x2 + 35.6x + 36.5 | 0.744 * | 7.7 |
| CPS | y = 203.1e−0.28x | 0.936 ** | 3.6 | |
| ACM | y = −3.15x2 + 30.0x + 135.1 | 0.786 ** | 11.3 | |
| Brassica juncea L. | IPW | y = −3.1x2 + 30.1x + 25.4 | 0.832 ** | 7.6 |
| CPS | y = 203.9e−0.28x | 0.863 ** | 3.6 | |
| ACM | y = −1.9x2 + 12.0x + 136.0 | 0.827 ** | 9.8 | |
| Brassica alboglabra L. | IPW | y = −3.1x2 + 27.9x + 38.4 | 0.750 * | 7.4 |
| CPS | y = 187.3e−0.26x | 0.792 ** | 3.5 | |
| ACM | y = −0.61x2 + 5.2x + 146.9 | 0.643 * | 16.0 | |
| Non-planted vegetable | IPW | y = −2.0x2 + 16.2x + 50.0 | 0.726 * | 6.1 |
| CPS | y = 213.4e−0.24x | 0.947 ** | 4.3 | |
| ACM | y = −2.7x2 + 23.2x + 140.8 | 0.800 ** | 11.4 |
| Vegetable | Treatment | Plant Height (cm) | Main Root Length (cm) | Total Weight (g/plant) | Aerial Part Weight (g/plant) |
|---|---|---|---|---|---|
| Ipomoea batatas L. | CK a | 69.5 ± 7.8 a c | 25.5 ± 0.7 b | 26.4 ± 2.6 b | 25.1 ± 2.3 b |
| IPW b | 67.7 ± 18.0 a | 24.0 ± 1.0 b | 16.4 ± 0.1 a | 15.5 ± 0.9 a | |
| CPS | 69.0 ± 5.7 a | 25.0 ± 0.1 b | 15.5 ± 3.2 a | 15.1 ± 4.2 a | |
| ACM | 53.8 ± 5.6 a | 16.8 ± 1.9 a | 14.3 ± 3.4 a | 13.1 ± 3.7 a | |
| Brassica juncea L. | CK | 58.0 ± 10.2 b | 21.7 ± 8.1 b | 55.6 ± 5.2 b | 54.1 ± 3.3 c |
| IPW | 39.7 ± 6.1 a | 10.0 ± 1.0 a | 39.8 ± 8.7 a | 37.6 ± 7.9 ab | |
| CPS | 46.7 ± 7.5 ab | 11.0 ± 2.7 a | 46.0 ± 2.0 ab | 39.6 ± 0.8 b | |
| ACM | 32.7 ± 4.7 a | 8.7 ± 0.6 a | 30.6 ± 7.5 a | 28.4 ± 0.4 a | |
| Brassica alboglabra L. | CK | 44.0 ± 7.0 c | 13.0 ± 6.1 b | 35.4 ± 4.6 c | 34.5 ± 4.5 c |
| IPW | 34.3 ± 1.8 b | 8.0 ± 0.1 a | 17.6 ± 5.5 b | 15.6 ± 7.5 ab | |
| CPS | 35.0 ± 1.4 ab | 10.5 ± 2.1 ab | 16.5 ± 2.2 b | 15.9 ± 2.2 b | |
| ACM | 25.6 ± 1.4 a | 8.7 ± 0.6 ab | 9.4 ± 0.9 a | 8.9 ± 0.9 a |
| Vegetable | Treatment | EDI (μg/kg/d) | RQ | Risk Level a |
|---|---|---|---|---|
| Ipomoea batatas L. | IPW | 0.013 ± 0.003 a b | 0.32 ± 0.08 a | Medium risk |
| CPS | 0.019 ± 0.002 a | 0.47 ± 0.04 a | Medium risk | |
| ACM | 0.022 ± 0.008 a | 0.55 ± 0.20 a | Medium risk | |
| Brassica juncea L. | IPW | 0.002 ± 0.000 a | 0.06 ± 0.008 a | Low risk |
| CPS | 0.002 ± 0.001 a | 0.05 ± 0.016 a | Low risk | |
| ACM | 0.003 ± 0.001 a | 0.09 ± 0.022 a | Low risk | |
| Brassica alboglabra L. | IPW | 0.004 ± 0.003 ab | 0.10 ± 0.01 ab | Low risk |
| CPS | 0.001 ± 0.000 a | 0.02 ± 0.01 a | Low risk | |
| ACM | 0.004 ± 0.001 b | 0.11 ± 0.01 b | Medium risk |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, L.; Li, Y.-W.; Wang, Z.-R.; Liu, B.-L.; Zhao, H.-M.; Li, H.; Cai, Q.-Y.; Mo, C.-H.; Li, Q.X. Bioaccumulation and Phytotoxicity and Human Health Risk from Microcystin-LR under Various Treatments: A Pot Study. Toxins 2020, 12, 523. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080523
Xiang L, Li Y-W, Wang Z-R, Liu B-L, Zhao H-M, Li H, Cai Q-Y, Mo C-H, Li QX. Bioaccumulation and Phytotoxicity and Human Health Risk from Microcystin-LR under Various Treatments: A Pot Study. Toxins. 2020; 12(8):523. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080523
Chicago/Turabian StyleXiang, Lei, Yan-Wen Li, Zhen-Ru Wang, Bai-Lin Liu, Hai-Ming Zhao, Hui Li, Quan-Ying Cai, Ce-Hui Mo, and Qing X. Li. 2020. "Bioaccumulation and Phytotoxicity and Human Health Risk from Microcystin-LR under Various Treatments: A Pot Study" Toxins 12, no. 8: 523. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080523