Zearalenone and Metabolites in Livers of Turkey Poults and Broiler Chickens Fed with Diets Containing Fusariotoxins

Abstract
:1. Introduction
2. Results and Discussion
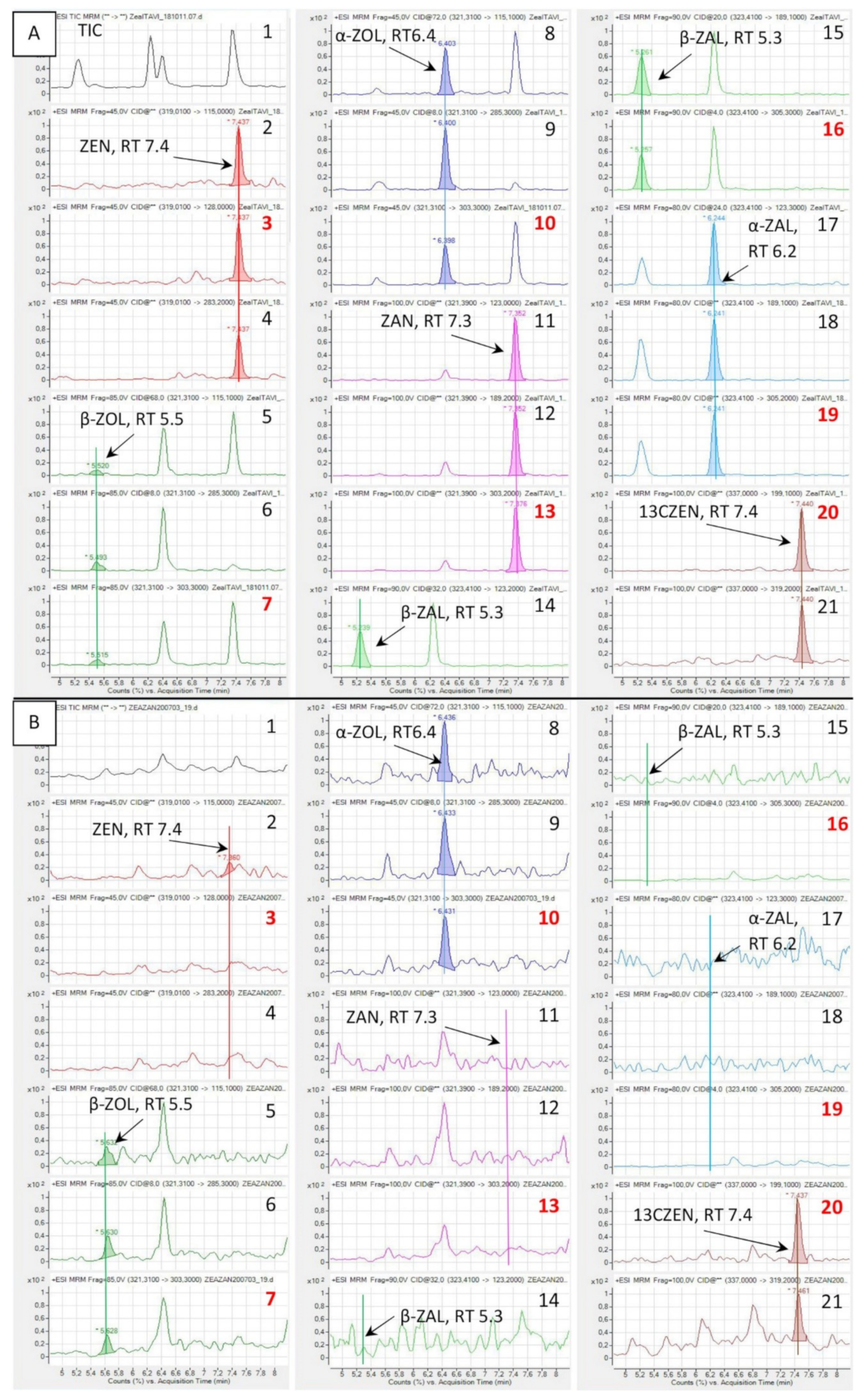
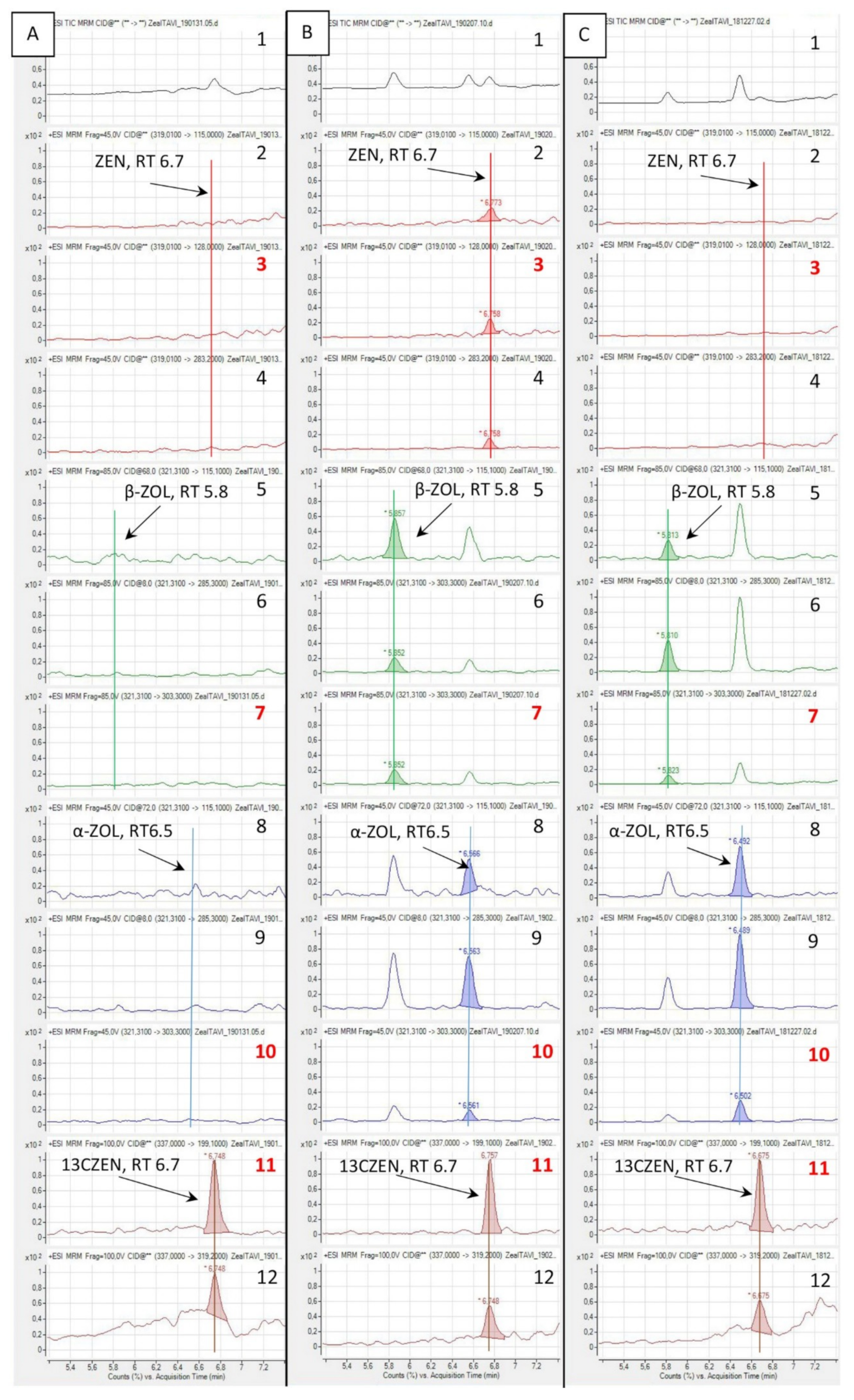
2.1. Separation of the Analytes and Matrix Interactions
2.2. Recovery of the Analytes and Determination of the LOQ
2.3. Detection of ZAN and ZALs
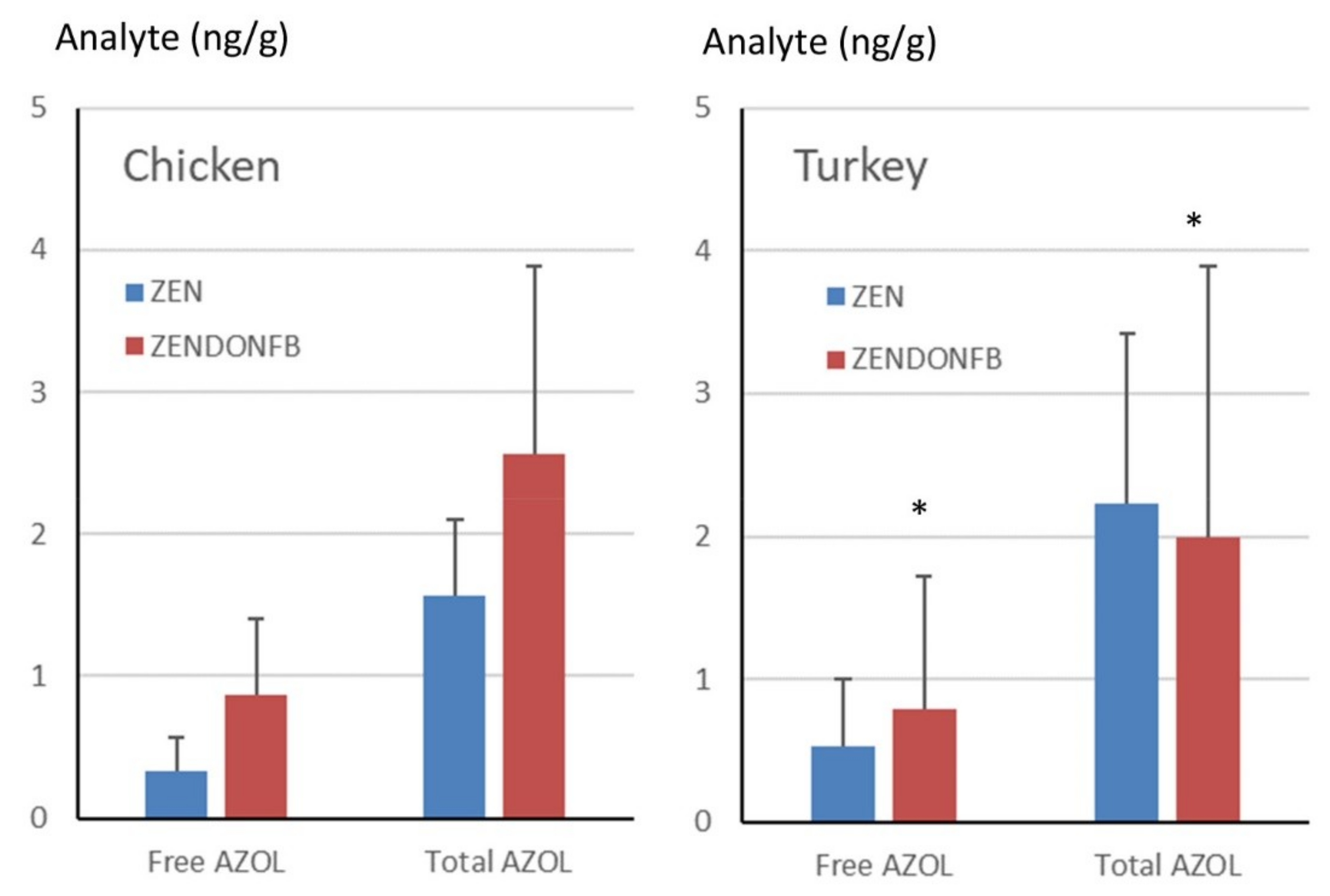
2.4. Total ZEN and ZOLs in Turkey Livers
2.5. Total ZEN and ZOLs in Chicken Livers
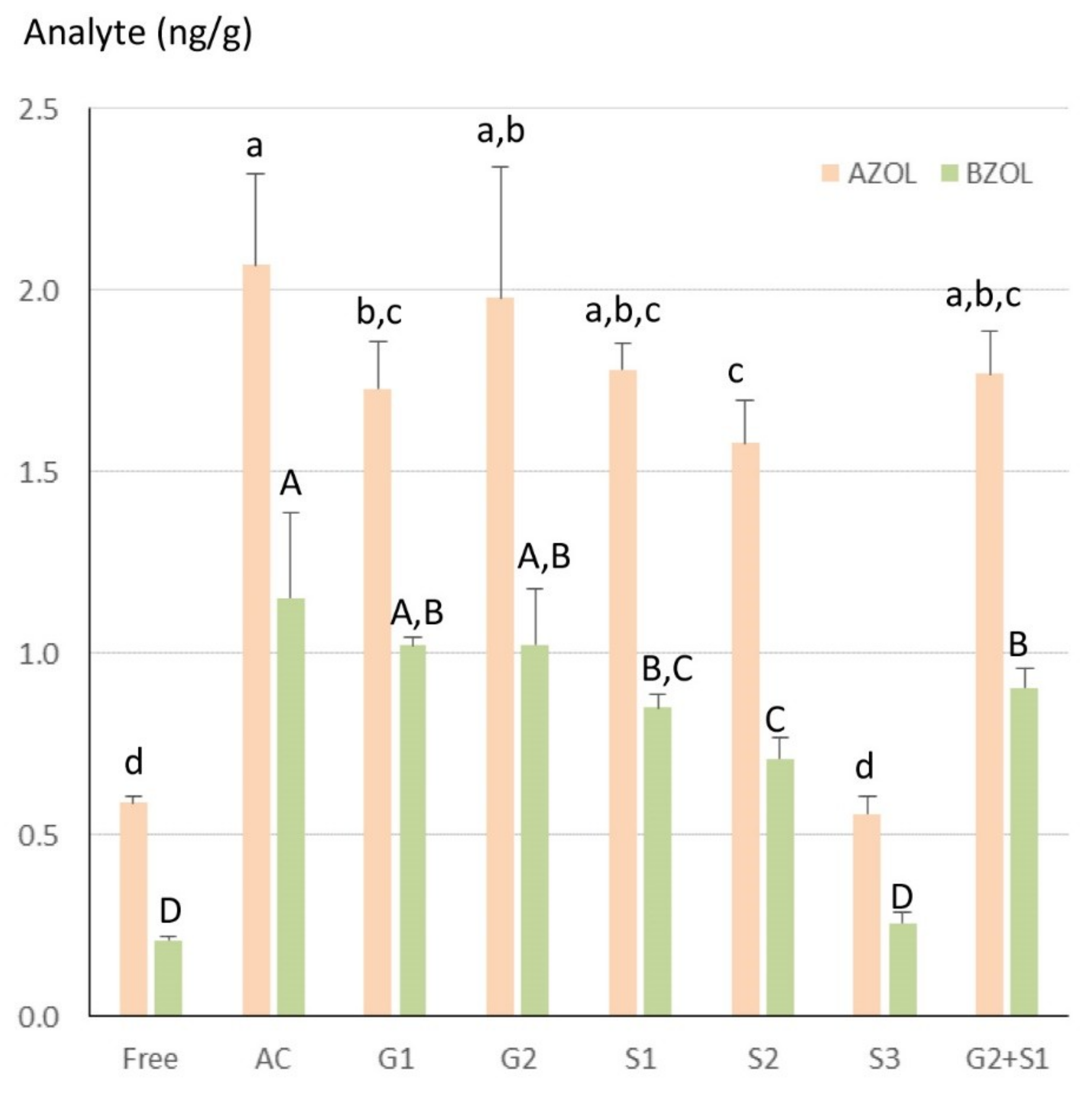
2.6. Hydrolysis of the Conjugates and Comparison of the Free Forms
2.7. General Discussion and Between-Study Comparison
3. Material and Methods
3.1. Analytes and Reagents
3.2. Tissue Samples
3.3. LC/MS/MS Conditions
3.4. Analysis of Standard Solutions
3.5. Matrix Interactions
3.6. Recovery Rates
3.7. Metabolites in Tissues
3.7.1. Total Metabolites
3.7.2. Free Metabolites
3.8. Comparison of Enzymes Used for the Hydrolysis and Acid Hydrolysis
3.9. Acceptability Parameters and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ZEN | zearalenone |
| ZAN | zearalanone |
| ZOLs | zearalenols |
| α-ZOL | alpha-zearalenol |
| β-ZOL | beta-zearalenol |
| FB | fumonisins B |
| FB1 | fumonisin B1 |
| FB2 | fumonisin B2 |
| DON | deoxynivalenol |
| RPFs | relative potency factors |
| TDI | tolerable daily intake |
| b.w. | body weight |
| IA | immunoaffinity |
| G1 | H-1 β-glucuronidase from Helix pomatia |
| G2 | H-2 β-glucuronidase from Helix pomatia |
| S1 | H-1 sulfatase from Helix pomatia |
| S2 | H-2 sulfatase from Helix pomatia |
| S3 | VI sulfatase from Aerobacter aerogenes |
| ACN | acetonitrile |
| MetOH | methanol |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry |
| UPLC-MS/MS | ultraperformance liquid chromatography-tandem mass spectrometry |
| MRM | multiple reaction monitoring |
| RE | extraction recovery |
| SSE | signal suppression and enhancement |
| SNR | signal-to-noise ratio |
| RSD | relative standard deviation |
| LOQ | limit of quantitation |
References
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.-K.; Alexander, J.; Barregard, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar] [PubMed] [Green Version]
- Zearalenone (WHO Food Additives Series 44). Available online: http://www.inchem.org/documents/jecfa/jecmono/v44jec14.htm (accessed on 2 August 2019).
- Guerre, P. Worldwide Mycotoxins Exposure in Pig and Poultry Feed Formulations. Toxins 2016, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Authority, E.F.S. Evaluation of the increase of risk for public health related to a possible temporary derogation from the maximum level of deoxynivalenol, zearalenone and fumonisins for maize and maize products. EFSA J. 2014, 12, 3699. [Google Scholar]
- Devreese, M.; Antonissen, G.; Broekaert, N.; De Baere, S.; Vanhaecke, L.; De Backer, P.; Croubels, S. Comparative toxicokinetics, absolute oral bioavailability, and biotransformation of zearalenone in different poultry species. J. Agric. Food Chem. 2015, 63, 5092–5098. [Google Scholar] [CrossRef] [PubMed]
- Guerre, P. Fusariotoxins in Avian Species: Toxicokinetics, Metabolism and Persistence in Tissues. Toxins 2015, 7, 2289–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Applegate, T. Zearalenone (ZEN) in Livestock and Poultry: Dose, Toxicokinetics, Toxicity and Estrogenicity. Toxins 2020, 12, 377. [Google Scholar] [CrossRef]
- Olsen, M.; Mirocha, C.J.; Abbas, H.K.; Johansson, B. Metabolism of high concentrations of dietary zearalenone by young male turkey poults. Poult. Sci. 1986, 65, 1905–1910. [Google Scholar] [CrossRef]
- Mauro, T.; Hao, L.; Pop, L.C.; Buckley, B.; Schneider, S.H.; Bandera, E.V.; Shapses, S.A. Circulating zearalenone and its metabolites differ in women due to body mass index and food intake. Food Chem. Toxicol. 2018, 116, 227–232. [Google Scholar] [CrossRef]
- Gambacorta, L.; Olsen, M.; Solfrizzo, M. Pig Urinary Concentration of Mycotoxins and Metabolites Reflects Regional Differences, Mycotoxin Intake and Feed Contaminations. Toxins 2019, 11, 378. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhao, H.; Xi, C.; Wang, G.; Chen, D.; Ding, S. Determination of chloramphenicol and zeranols in pig muscle by immunoaffinity column clean-up and LC-MS/MS analysis. Food Addit. Contam. Part A 2014, 31, 1171–1186. [Google Scholar] [CrossRef]
- Songsermsakul, P.; Sontag, G.; Cichna-Markl, M.; Zentek, J.; Razzazi-Fazeli, E. Determination of zearalenone and its metabolites in urine, plasma and faeces of horses by HPLC–APCI–MS. J. Chromatogr. B 2006, 843, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Veršilovskis, A.; Huybrecht, B.; Tangni, E.K.; Pussemier, L.; De Saeger, S.; Callebaut, A. Cross-reactivity of some commercially available deoxynivalenol (DON) and zearalenone (ZEN) immunoaffinity columns to DON- and ZEN-conjugated forms and metabolites. Food Addit. Contam. Part A 2011, 28, 1687–1693. [Google Scholar] [CrossRef] [PubMed]
- Olsen, M.E.; Pettersson, H.I.; Sandholm, K.A.; Kiessling, K.H. Quantitative liquid chromatographic method using fluorescence detection for determining zearalenone and its metabolites in blood plasma and urine. J. Assoc. Off. Anal. Chem. 1985, 68, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Tulayakul, P.; Li, J.-Y.; Dong, K.-S.; Manabe, N.; Kumagai, S. Metabolic conversion of zearalenone to alpha-zearalenol by goat tissues. J. Vet. Med. Sci. 2010, 72, 307–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, N.; Degen, G.H. Urinary biomarkers of exposure to the mycoestrogen zearalenone and its modified forms in German adults. Arch. Toxicol. 2018, 92, 2691–2700. [Google Scholar] [CrossRef]
- Solfrizzo, M.; Gambacorta, L.; Lattanzio, V.M.T.; Powers, S.; Visconti, A. Simultaneous LC-MS/MS determination of aflatoxin M1, ochratoxin A, deoxynivalenol, de-epoxydeoxynivalenol, α and β-zearalenols and fumonisin B1 in urine as a multi-biomarker method to assess exposure to mycotoxins. Anal. Bioanal. Chem. 2011, 401, 2831–2841. [Google Scholar] [CrossRef]
- Woźny, M.; Obremski, K.; Jakimiuk, E.; Gusiatin, M.; Brzuzan, P. Zearalenone contamination in rainbow trout farms in north-eastern Poland. Aquaculture 2013, 416, 209–211. [Google Scholar] [CrossRef]
- De Baere, S.; Osselaere, A.; Devreese, M.; Vanhaecke, L.; De Backer, P.; Croubels, S. Development of a liquid–chromatography tandem mass spectrometry and ultra-high-performance liquid chromatography high-resolution mass spectrometry method for the quantitative determination of zearalenone and its major metabolites in chicken and pig plasma. Anal. Chim. Acta 2012, 756, 37–48. [Google Scholar] [CrossRef]
- Winkler, J.; Kersten, S.; Meyer, U.; Engelhardt, U.; Dänicke, S. Residues of zearalenone (ZEN), deoxynivalenol (DON) and their metabolites in plasma of dairy cows fed Fusarium contaminated maize and their relationships to performance parameters. Food Chem. Toxicol. 2014, 65, 196–204. [Google Scholar] [CrossRef]
- Pajewska, M.; Łojko, M.; Cendrowski, K.; Sawicki, W.; Kowalkowski, T.; Buszewski, B.; Gadzała-Kopciuch, R. The determination of zearalenone and its major metabolites in endometrial cancer tissues. Anal. Bioanal. Chem. 2018, 410, 1571–1582. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Wang, L.; Wang, J.; Tan, Y.; Yu, D.; Chang, X.; Fan, Y.; Zhao, D.; Wang, C.; De Boevre, M.; et al. A QuEChERS-Based Liquid Chromatography-Tandem Mass Spectrometry Method for the Simultaneous Determination of Nine Zearalenone-Like Mycotoxins in Pigs. Toxins 2018, 10, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Li, C.; Zhou, S.; Zhao, Y.; Gong, Y.Y.; Gong, Z.; Wu, Y. Determination of Trace Zearalenone and Its Metabolites in Human Serum by a High-Throughput UPLC-MS/MS Analysis. Appl. Sci. 2019, 9, 741. [Google Scholar] [CrossRef] [Green Version]
- Belhassen, H.; Jiménez-Díaz, I.; Ghali, R.; Ghorbel, H.; Molina-Molina, J.M.; Olea, N.; Hedili, A. Validation of a UHPLC–MS/MS method for quantification of zearalenone, α-zearalenol, β-zearalenol, α-zearalanol, β-zearalanol and zearalanone in human urine. J. Chromatogr. B 2014, 962, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Deng, C.; Zhou, S.; Zhao, Y.; Wang, D.; Wang, X.; Gong, Y.Y.; Wu, Y. High-throughput and sensitive determination of urinary zearalenone and metabolites by UPLC-MS/MS and its application to a human exposure study. Anal. Bioanal. Chem. 2018, 410, 5301–5312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolf-Clauw, M.; Ayouni, F.; Tardieu, D.; Guerre, P. Variations in zearalenone activation in avian food species. Food Chem. Toxicol. 2008, 46, 1467–1473. [Google Scholar] [CrossRef]
- Metayer, J.-P.; Travel, A.; Mika, A.; Bailly, J.-D.; Cleva, D.; Boissieu, C.; Guennec, J.L.; Froment, P.; Albaric, O.; Labrut, S.; et al. Lack of Toxic Interaction between Fusariotoxins in Broiler Chickens Fed throughout Their Life at the Highest Level Tolerated in the European Union. Toxins 2019, 11, 455. [Google Scholar] [CrossRef] [Green Version]
- Travel, A.; Metayer, J.-P.; Mika, A.; Bailly, J.-D.; Cleva, D.; Boissieu, C.; Guennec, J.L.; Albaric, O.; Labrut, S.; Lepivert, G.; et al. Toxicity of Fumonisins, Deoxynivalenol, and Zearalenone Alone and in Combination in Turkeys Fed with the Maximum European Union–Tolerated Level. Avian Dis. 2019, 63, 703–712. [Google Scholar] [CrossRef]
- Zou, Y.; Yang, Z.B.; Yang, W.R.; Jiang, S.Z.; Zhang, G.G.; Chi, F. Effect of purified zearalenone on nutrient digestibility in broilers fed 2 levels of fumonisin from naturally contaminated corn (Zea mays). J. Appl. Poult. Res. 2012, 21, 251–258. [Google Scholar] [CrossRef]
- Chen, S.S.; Li, Y.-H.; Lin, M.-F. Chronic Exposure to the Fusarium Mycotoxin Deoxynivalenol: Impact on Performance, Immune Organ, and Intestinal Integrity of Slow-Growing Chickens. Toxins 2017, 9, 334. [Google Scholar] [CrossRef] [Green Version]
- Awad, W.A.; Hess, M.; Twarużek, M.; Grajewski, J.; Kosicki, R.; Böhm, J.; Zentek, J. The impact of the Fusarium mycotoxin deoxynivalenol on the health and performance of broiler chickens. Int. J. Mol. Sci. 2011, 12, 7996–8012. [Google Scholar] [CrossRef]
- Yunus, A.W.; Blajet-Kosicka, A.; Kosicki, R.; Khan, M.Z.; Rehman, H.; Böhm, J. Deoxynivalenol as a contaminant of broiler feed: Intestinal development, absorptive functionality, and metabolism of the mycotoxin. Poult. Sci. 2012, 91, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Croubels, S.; Pasmans, F.; Ducatelle, R.; Eeckhaut, V.; Devreese, M.; Verlinden, M.; Haesebrouck, F.; Eeckhout, M.; De Saeger, S.; et al. Fumonisins affect the intestinal microbial homeostasis in broiler chickens, predisposing to necrotic enteritis. Vet. Res. 2015, 46, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Janssens, G.P.J.; De Baere, S.; Mountzouris, K.C.; Su, S.; Wong, E.A.; De Meulenaer, B.; et al. Mycotoxins Deoxynivalenol and Fumonisins Alter the Extrinsic Component of Intestinal Barrier in Broiler Chickens. J. Agric. Food Chem. 2015, 63, 10846–10855. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Devreese, M.; Van Immerseel, F.; De Baere, S.; Hessenberger, S.; Martel, A.; Croubels, S. Chronic exposure to deoxynivalenol has no influence on the oral bioavailability of fumonisin B1 in broiler chickens. Toxins 2015, 7, 560–571. [Google Scholar] [CrossRef] [Green Version]
- Jodlbauer, J.; Zöllner, P.; Lindner, W. Determination of zeranol, taleranol, zearalenone, α- and β-zearalenol in urine and tissue by high-performance liquid chromatography-tandem mass spectrometry. Chromatographia 2000, 51, 681–687. [Google Scholar] [CrossRef]
- Zhu, R.; Zhao, Z.; Wang, J.; Bai, B.; Wu, A.; Yan, L.; Song, S. A simple sample pretreatment method for multi-mycotoxin determination in eggs by liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2015, 1417, 1–7. [Google Scholar] [CrossRef]
- Soleimany, F.; Jinap, S.; Faridah, A.; Khatib, A. A UPLC–MS/MS for simultaneous determination of aflatoxins, ochratoxin a, zearalenone, DON, fumonisins, T-2 toxin and HT-2 toxin, in cereals. Food Control 2012, 25, 647–653. [Google Scholar] [CrossRef]
- β-Glucuronidase from Helix Pomatia Type H-1, Partially Purified Powder, ≥300,000 units/g solid|Sigma-Aldrich. Available online: https://www.sigmaaldrich.com/catalog/product/sigma/g0751?lang=fr®ion=FR (accessed on 29 July 2020).
- Sulfatase from Helix Pomatia Type H-2, Aqueous Solution, ≥2,000 Units/mL|Aryl-Sulfatase|Sigma-Aldrich. Available online: https://www.sigmaaldrich.com/catalog/product/sigma/s9751?lang=fr®ion=FR (accessed on 29 July 2020).
- Dänicke, S.; Ueberschär, K.H.; Halle, I.; Matthes, S.; Valenta, H.; Flachowsky, G. Effect of addition of a detoxifying agent to laying hen diets containing uncontaminated or Fusarium toxin-contaminated maize on performance of hens and on carryover of zearalenone. Poult. Sci. 2002, 81, 1671–1680. [Google Scholar] [CrossRef]
- Dänicke, S.; Ueberschär, K.H.; Halle, I.; Valenta, H.; Flachowsky, G. Excretion kinetics and metabolism of zearalenone in broilers in dependence on a detoxifying agent. Arch. Tierernahr. 2001, 55, 299–313. [Google Scholar] [CrossRef]
- Mirocha, C.J.; Robison, T.S.; Pawlosky, R.J.; Allen, N.K. Distribution and residue determination of [3H]zearalenone in broilers. Toxicol. Appl. Pharmacol. 1982, 66, 77–87. [Google Scholar] [CrossRef]
- Catteuw, A.; Broekaert, N.; De Baere, S.; Lauwers, M.; Gasthuys, E.; Huybrechts, B.; Callebaut, A.; Ivanova, L.; Uhlig, S.; De Boevre, M.; et al. Insights into In Vivo Absolute Oral Bioavailability, Biotransformation, and Toxicokinetics of Zearalenone, α-Zearalenol, β-Zearalenol, Zearalenone-14-glucoside, and Zearalenone-14-sulfate in Pigs. J. Agric. Food Chem. 2019, 67, 3448–3458. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.; Royce, S.G.; Alexander, J.A.; Buckley, B.; Isukapalli, S.S.; Bandera, E.V.; Zarbl, H.; Georgopoulos, P.G. Physiologically-Based Toxicokinetic Modeling of Zearalenone and Its Metabolites: Application to the Jersey Girl Study. PLoS ONE 2014, 9, e113632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Wu, W.; Pan, J.; Long, M. Detoxification Strategies for Zearalenone Using Microorganisms: A Review. Microorganisms 2019, 7, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, N.; Dänicke, S.; Edler, L.; Gottschalk, C.; Lassek, E.; Marko, D.; Rychlik, M.; Mally, A. A critical evaluation of health risk assessment of modified mycotoxins with a special focus on zearalenone. Mycotoxin Res. 2019, 35, 27–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hort, V.; Nicolas, M.; Travel, A.; Jondreville, C.; Maleix, C.; Baéza, E.; Engel, E.; Guérin, T. Carry-over assessment of fumonisins and zearalenone to poultry tissues after exposure of chickens to a contaminated diet – A study implementing stable-isotope dilution assay and UHPLC-MS/MS. Food Control 2020, 107, 106789. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, H.; Sun, F.; De Ruyck, K.; Zhang, J.; Jin, Y.; Li, Y.; Wang, Z.; Zhang, S.; De Saeger, S.; et al. Metabolic Profile of Zearalenone in Liver Microsomes from Different Species and Its in Vivo Metabolism in Rats and Chickens Using Ultra High-Pressure Liquid Chromatography-Quadrupole/Time-of-Flight Mass Spectrometry. J. Agric. Food Chem. 2017, 65, 11292–11303. [Google Scholar] [CrossRef]
- Devreese, M.; Antonissen, G.; Broekaert, N.; De Mil, T.; De Baere, S.; Vanhaecke, L.; De Backer, P.; Croubels, S. Toxicokinetic study and oral bioavailability of deoxynivalenol in turkey poults, and comparative biotransformation between broilers and turkeys. World Mycotoxin J. 2015, 8, 533–539. [Google Scholar] [CrossRef]
- Wan, D.; Huang, L.; Pan, Y.; Wu, Q.; Chen, D.; Tao, Y.; Wang, X.; Liu, Z.; Li, J.; Wang, L.; et al. Metabolism, distribution, and excretion of deoxynivalenol with combined techniques of radiotracing, high-performance liquid chromatography ion trap time-of-flight mass spectrometry, and online radiometric detection. J. Agric. Food Chem. 2014, 62, 288–296. [Google Scholar] [CrossRef]
- Malekinejad, H.; Maas-Bakker, R.; Fink-Gremmels, J. Species differences in the hepatic biotransformation of zearalenone. Vet. J. Lond. Engl. 1997 2006, 172, 96–102. [Google Scholar] [CrossRef]
- ANSES_GuideValidation. Available online: https://www.anses.fr/fr/system/files/ANSES_GuideValidation.pdf (accessed on 19 December 2018).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α-ZAL | α-ZOL | β-ZAL | β-ZOL | ZAN | ZEN | [13C18]-ZEN | |
|---|---|---|---|---|---|---|---|
| Method 1 | |||||||
| SSE Liver (%) 1 | 88 | 87 | 63 | 25 | 84 | 75 | 97 |
| SSE Muscle (%) 1 | 108 | 106 | 55 | 98 | 105 | 73 | - |
| SNR Liver 2 | 6.5 | 8.3 | 4.7 | 0.7 | 4.5 | 2.4 | - |
| SNR Muscle 2 | 7.4 | 3.2 | 3.2 | 2 | 3.1 | 3.1 | - |
| Method 2 | |||||||
| SSE Liver (%) 1 | - | 83 | - | 99 | - | 80 | 78 |
| SSE Muscle (%) 1 | - | 94 | - | 94 | - | 84 | - |
| SNR Liver 2 | - | 7.3 | - | 6.7 | - | 2.3 | - |
| SNR Muscle 2 | 1.9 | - | 2.1 | - | 1.7 | - |
| α-ZAL | α-ZOL | β-ZAL | β-ZOL | ZAN | ZEN | [13C18]-ZEN | |
|---|---|---|---|---|---|---|---|
| Extraction Recovery (%) 1 | 76 ± 6 | 71 ± 8 | 44 ± 6 | 16 ± 11 | 70 ± 7 | 65 ± 10 | 57 ± 7 |
| Recovery 2 | 0.30 ± 0.04 (120%) | 0.28 ± 0.04 (113%) | 0.27 ± 0.05 (107%) | 0.32 ± 0.1 (127%) | 0.28 ± 0.02 (111%) | 0.26 ± 0.06 (104%) |
| α-ZOL | β-ZOL | ZEN | [13C18]-ZEN | |
|---|---|---|---|---|
| Extraction Recovery (%) 1 | 55 ± 12 | 62 ± 10 | 52 ± 8 | 55 ± 9 |
| Liver Spiked at 0.25 ng/g 2 | 0.29 ± 0.06 (116%) | 0.28 ± 0.01 (110%) | 0.25 ± 0.03 (101%) | |
| Liver Spiked at 1 ng/g 2 | 1.08 ± 0.05 (108%) | 1.08 ± 0.09 (108%) | 0.92 ± 0.09 (92%) | |
| Liver Spiked at 5 ng/g 2 | 5.20 ± 0.51 (104%) | 5.11 ± 0.85 (102%) | 4.60 ± 0.58 (97%) | |
| Liver Spiked at 1 ng/g with a Hydrolysis Step 2,3 | 0.96 ± 0.06 (96%) | 0.98 ± 0.05 (98%) | 0.84 ± 0.12 (84%) | |
| Intra-Day RSD (%) | 5 | 7 | 9 | |
| Inter-Day RSD (%) | 10 | 14 | 18 |
| α-ZOL | β-ZOL | ZEN | |
|---|---|---|---|
| Exposure (Days) | 14 | 14 | 14 |
| Feed Control (µg/kg) 55 to 70 d | <10 | <10 | 35 |
| Liver positive/total max; min (ng/g) | 2/8 0.44; <0.25 | 0/8 <0.25 | 0/8 <0.25 |
| Feed ZEN (µg/kg) 55 to 70 d | <10 | <10 | 470 |
| Liver (ng/g) mean ± SD max; min positive/total | 2.23 ± 1.19 4.08; 0.55 8/8 | 1.14 ± 0.83 2.14; <0.25 8/8 | 0.54 ± 0.43 2.06; <0.25 4/8 |
| Feed ZENDONFB (µg/kg) 2 55 to 70 d | <10 | <10 | 570 |
| Liver (ng/g) mean ± SD max; min positive/total | 1.99 ± 1.90 7.63; 0.63 8/8 | 1.00 ± 0.75 2.45; <0.25 8/8 | 0.50 ± 0.39 1.57; <0.25 5/8 |
| α-ZOL | β-ZOL | ZEN | |
|---|---|---|---|
| Exposure (Days) | 35 | 35 | 35 |
| Feed Control (µg/kg) 1 to 10 d | <10 | <10 | 35 |
| 11 to 35 d | <10 | <10 | 25 |
| Liver positive/total max; min (ng/g) | 1/8 0.27; <0.25 | 0/8 <0.25 | 0/8 <0.25 |
| Feed ZEN (µg/kg) 1 to 10 d | <10 | <10 | 465 |
| 11 to 35 d | <10 | <10 | 480 |
| Liver (ng/g) mean ± SD max; min positive/total | 1.56 ± 0.86 * 3.98; 0.52 8/8 | 0.80 ± 0.59 1.64; <0.25 7/8 | 0.27 ± 0.14 0.57; <0.25 3/8 |
| Feed ZENDONFB (µg/kg) 2 1 to 10 d | <10 | <10 | 415 |
| 11 to 35 d | <10 | <10 | 430 |
| Liver (ng/g) mean ± SD max; min positive/total | 2.57 ± 1.32 * 4.48; 0.76 8/8 | 1.37 ± 1.02 3.05; 0.52 8/8 | 0.35 ± 0.22 0.82; <0.25 3/8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tardieu, D.; Travel, A.; Metayer, J.-P.; Le Bourhis, C.; Guerre, P. Zearalenone and Metabolites in Livers of Turkey Poults and Broiler Chickens Fed with Diets Containing Fusariotoxins. Toxins 2020, 12, 525. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080525
Tardieu D, Travel A, Metayer J-P, Le Bourhis C, Guerre P. Zearalenone and Metabolites in Livers of Turkey Poults and Broiler Chickens Fed with Diets Containing Fusariotoxins. Toxins. 2020; 12(8):525. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080525
Chicago/Turabian StyleTardieu, Didier, Angelique Travel, Jean-Paul Metayer, Celeste Le Bourhis, and Philippe Guerre. 2020. "Zearalenone and Metabolites in Livers of Turkey Poults and Broiler Chickens Fed with Diets Containing Fusariotoxins" Toxins 12, no. 8: 525. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080525