Whole Genome Sequencing Reveals Virulence Potentials of Helicobacter pylori Strain KE21 Isolated from a Kenyan Patient with Gastric Signet Ring Cell Carcinoma

 ,
,
Abstract
:1. Introduction
2. Results
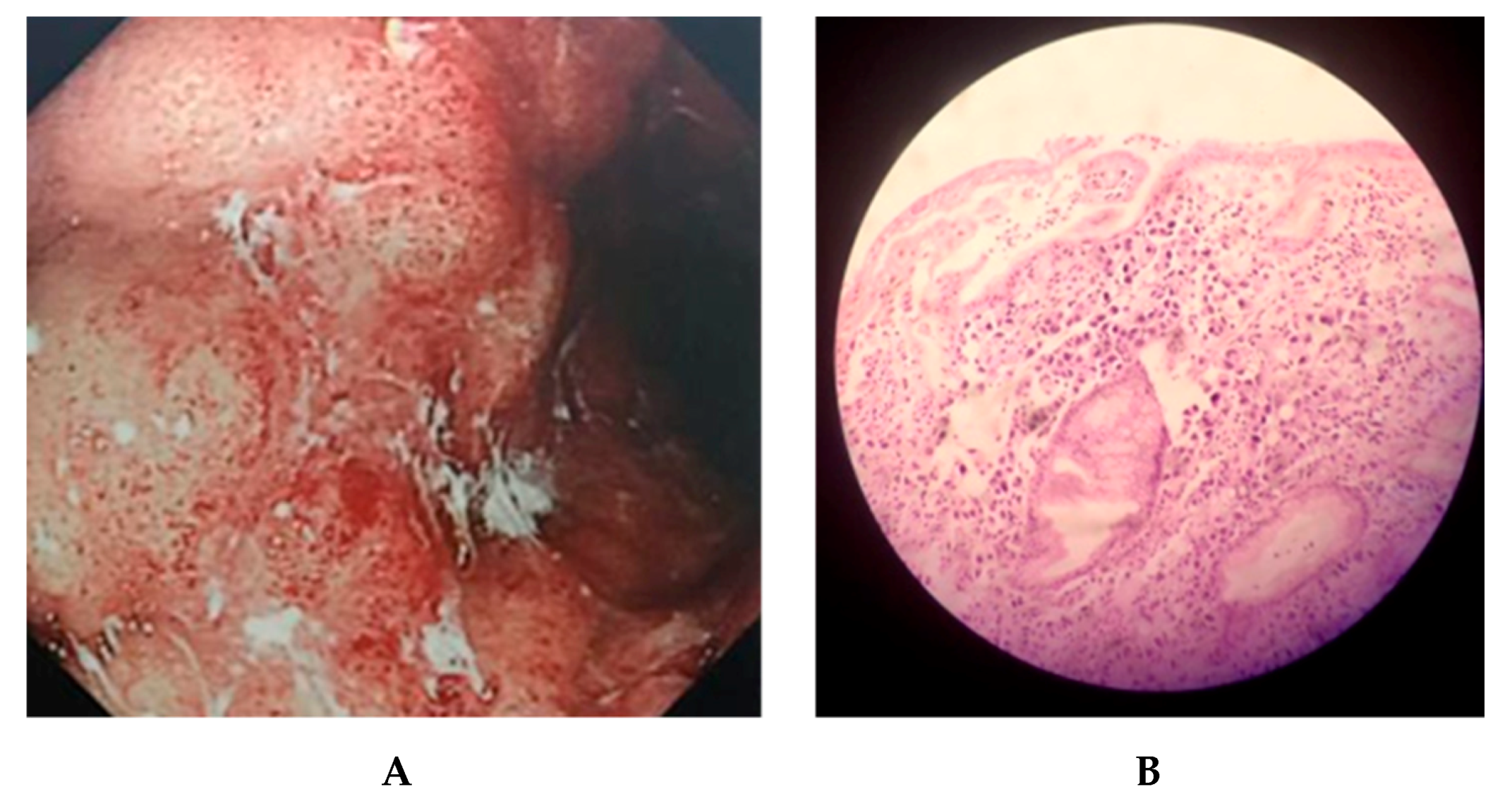
2.1. Clinical Outcomes of KE21 Clinical Isolate
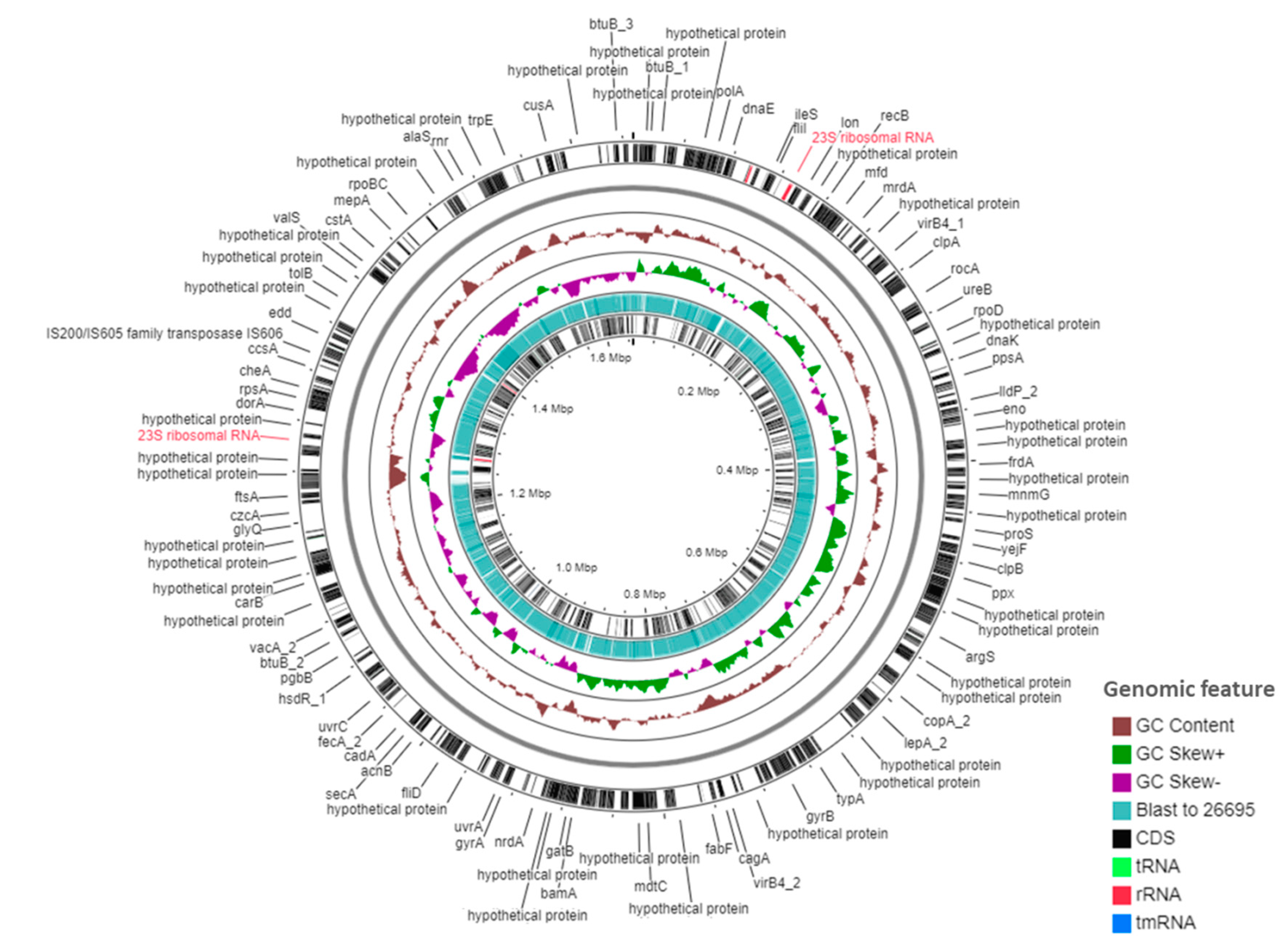
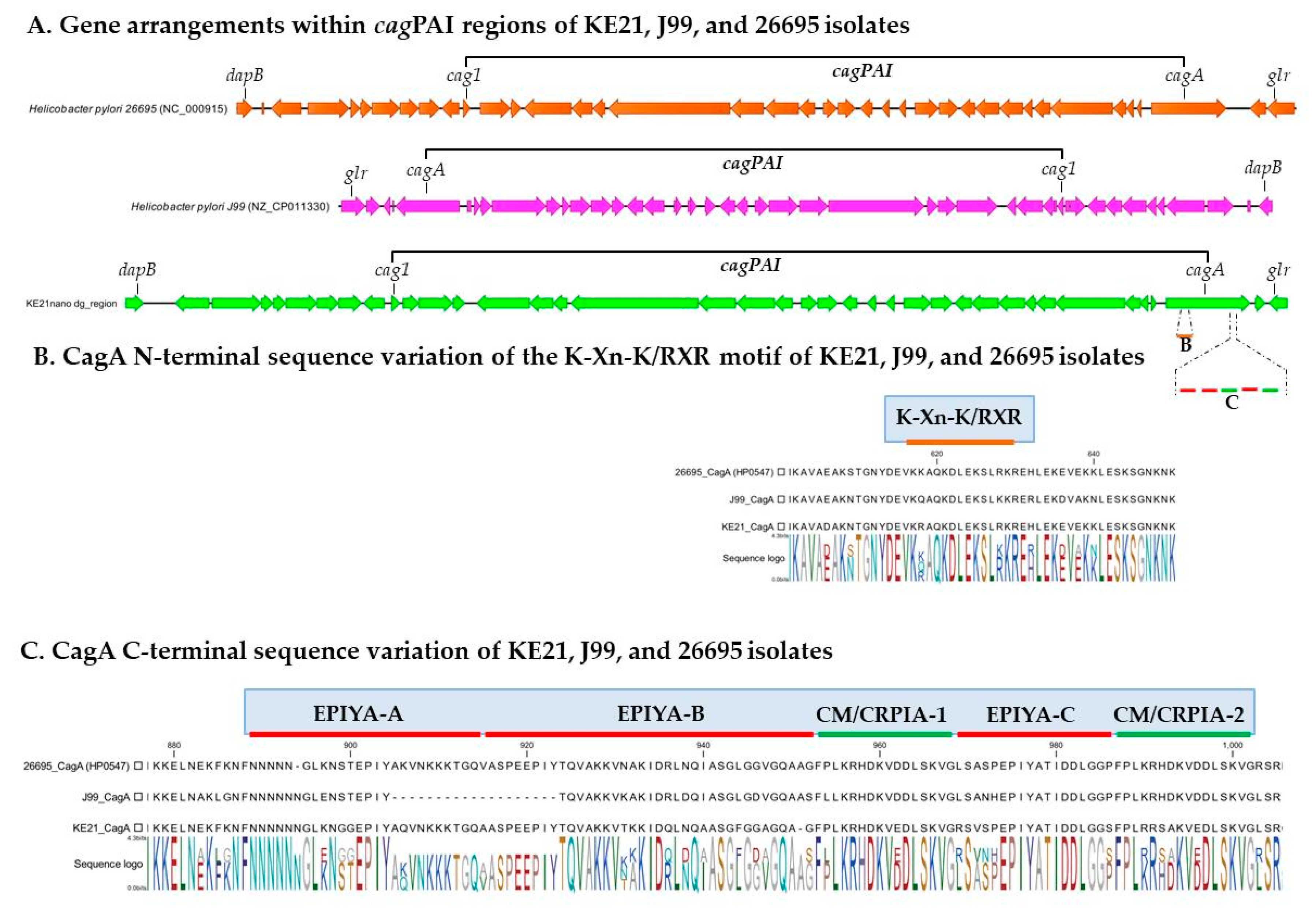
2.2. Genomic Features of KE21 Clinical Isolate
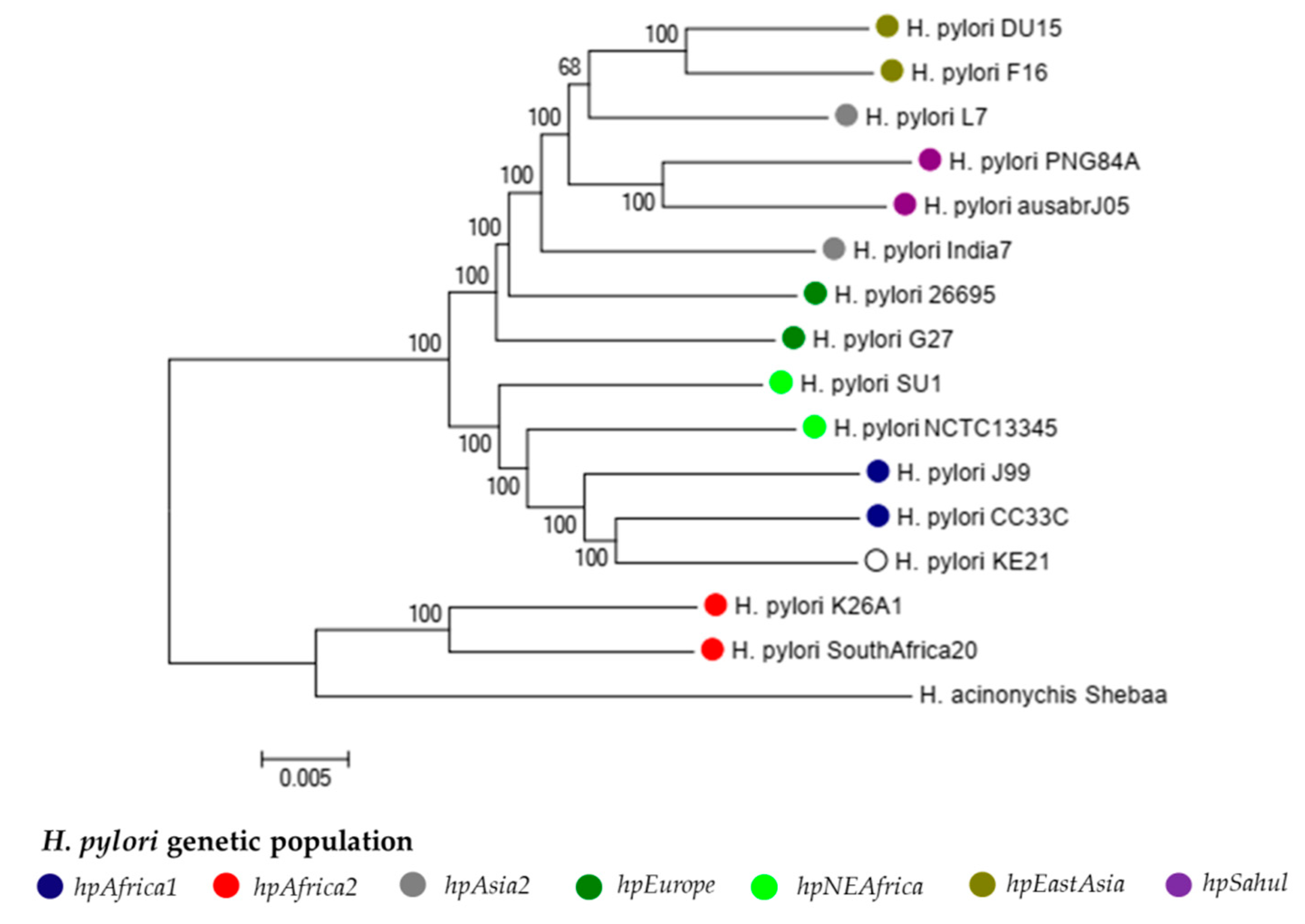
2.2.1. Population Genetics of Strain KE21
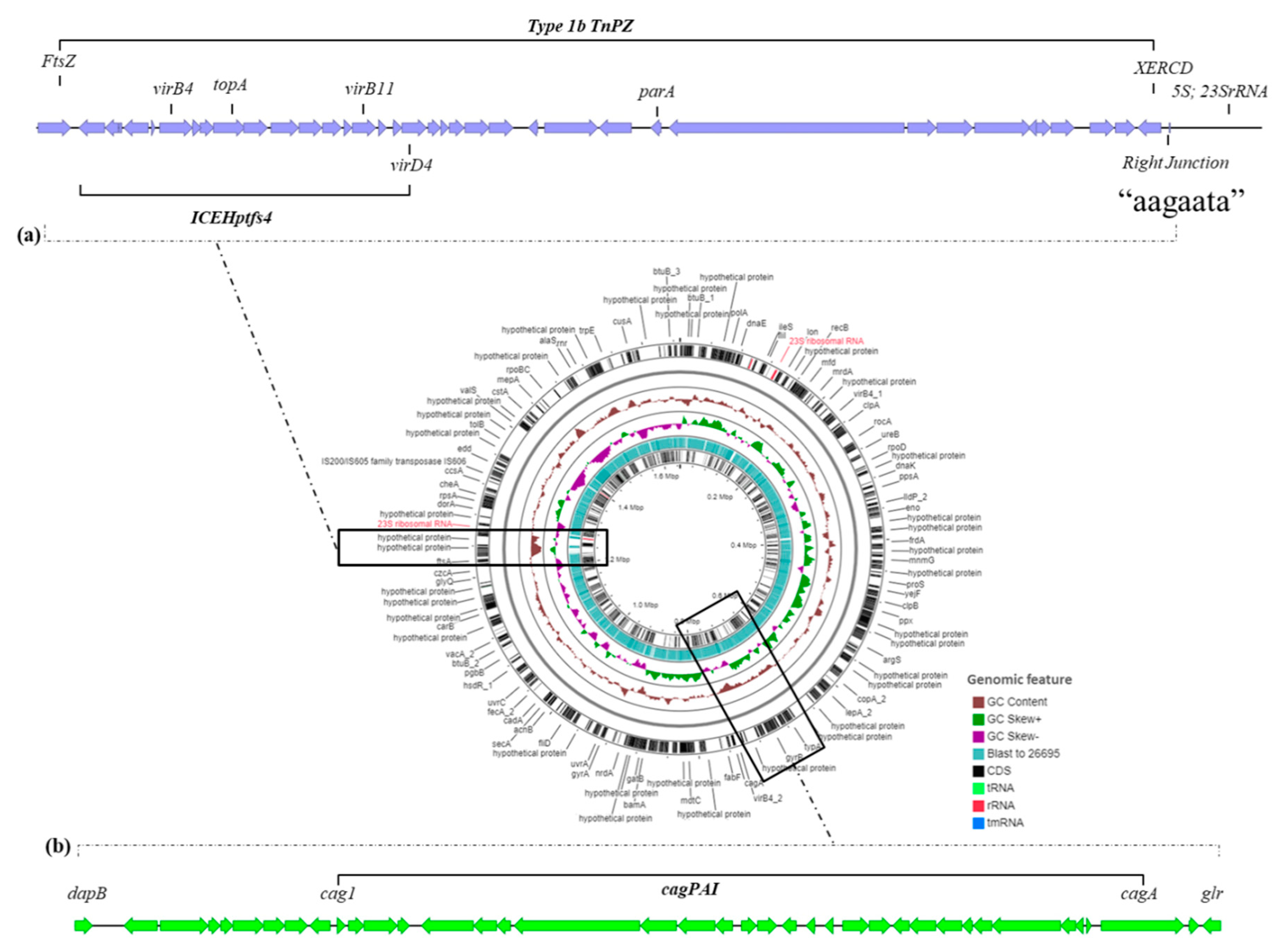
2.2.2. The Mobilome of Strain KE21
2.2.3. The Virulome of Strain KE21
2.3. Experimental Virulence Assays on KE21
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Patient and Biological Sampling
5.2. Antimicrobial Susceptibility Testing
5.3. DNA Extraction, Library Preparation, and Whole Genome Sequencing
5.3.1. Short-Read Illumina Sequencing
5.3.2. Long-Read MinION Sequencing.
5.4. Bioinformatic Analyses
5.5. AGS Cell Line Co-Infection with H. pylori KE21
5.6. Nucleotide Sequence Accession Number
5.7. Ethical Consideration
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Polk, D.B.; Peek, R.M., Jr. Helicobacter pylori: Gastric cancer and beyond. Nat. Rev. Cancer 2010, 10, 403–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loughlin, M.F. Novel therapeutic targets in Helicobacter pylori. Expert Opin. Ther. Targets 2003, 7, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Linz, B.; Balloux, F.; Moodley, Y.; Manica, A.; Liu, H.; Roumagnac, P.; Falush, D.; Stamer, C.; Prugnolle, F.; van der Merwe, S.W.; et al. An African origin for the intimate association between humans and Helicobacter pylori. Nature 2007, 445, 915–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsonnet, J.; Friedman, G.D.; Vandersteen, D.P.; Chang, Y.; Vogelman, J.H.; Orentreich, N.; Sibley, R.K. Helicobacter pylori infection and the risk of gastric carcinoma. N. Engl. J. Med. 1991, 325, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N.; Okamoto, S. Effect of Helicobacter pylori eradication on subsequent development of cancer after endoscopic resection of early gastric cancer in Japan. Gastroenterol. Clin. N. Am. 2000, 29, 819–827. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Schistosomes, liver flukes and Helicobacter pylori, In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 1994; Volume 61, pp. 1–241. [Google Scholar]
- Yamaoka, Y. Mechanisms of disease: Helicobacter pylori virulence factors. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaoka, Y.; Kato, M.; Asaka, M. Geographic differences in gastric cancer incidence can be explained by differences between Helicobacter pylori strains. Intern. Med. 2008, 47, 1077–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holcombe, C. Helicobacter pylori: The African enigma. Gut 1992, 33, 429–431. [Google Scholar] [CrossRef] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Graham, D.Y.; Lu, H.; Yamaoka, Y. African, Asian or Indian enigma, the East Asian Helicobacter pylori: Facts or medical myths. J. Dig. Dis. 2009, 10, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Campbell, D.I.; Warren, B.F.; Thomas, J.E.; Figura, N.; Telford, J.L.; Sullivan, P.B. The African enigma: Low prevalence of gastric atrophy, high prevalence of chronic inflammation in West African adults and children. Helicobacter 2001, 6, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Asombang, A.W.; Rahman, R.; Ibdah, J.A. Gastric cancer in Africa: Current management and outcomes. World J. Gastroenterol. 2014, 20, 3875–3879. [Google Scholar] [CrossRef] [PubMed]
- Agha, A.; Graham, D.Y. Evidence-based examination of the African enigma in relation to Helicobacter pylori infection. Scand. J. Gastroenterol. 2005, 40, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, U.C.; Chaturvedi, R.; Correa, P. The enigma of Helicobacter pylori infection and gastric cancer. Indian J. Gastroenterol. Off. J. Indian Soc. Gastroenterol. 2010, 29, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Asombang, A.W.; Kelly, P. Gastric cancer in Africa: what do we know about incidence and risk factors? Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 69–74. [Google Scholar] [CrossRef]
- Juul, S.; Izquierdo, F.; Hurst, A.; Dai, X.; Wright, A.; Kulesha, E.; Pettett, R.; Turner, D.J. What’s in my pot? Real-time species identification on the MinION™. BioRxiv 2015, 030742. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Montano, V.; Didelot, X.; Foll, M.; Linz, B.; Reinhardt, R.; Suerbaum, S.; Moodley, Y.; Jensen, J.D. Worldwide population structure, long-term demography, and local adaptation of Helicobacter pylori. Genetics 2015, 200, 947–963. [Google Scholar] [CrossRef] [Green Version]
- Yahara, K.; Furuta, Y.; Oshima, K.; Yoshida, M.; Azuma, T.; Hattori, M.; Uchiyama, I.; Kobayashi, I. Chromosome painting in silico in a bacterial species reveals fine population structure. Mol. Biol. Evol. 2013, 30, 1454–1464. [Google Scholar] [CrossRef] [Green Version]
- Roosaare, M.; Puustusmaa, M.; Möls, M.; Vaher, M.; Remm, M. PlasmidSeeker: Identification of known plasmids from bacterial whole genome sequencing reads. PeerJ 2018, 6, e4588. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, W.; Breithaupt, U.; Kern, B.; Smith, S.I.; Spicher, C.; Haas, R. A comprehensive analysis of Helicobacter pylori plasticity zones reveals that they are integrating conjugative elements with intermediate integration specificity. BMC Genom. 2014, 15, 310. [Google Scholar] [CrossRef] [Green Version]
- Kersulyte, D.; Lee, W.; Dharmalingam Subramaniam, S.A.; Herrera, P.; Cabrera, L.; Balqui, J.; Barabas, O.; Kalia, A.; Gilman, R.H.; Berg, D.E. Helicobacter pylori’s plasticity zones are novel transposable elements. PLoS ONE 2009, 4, e6859. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Tissera, K.; Jang, S.; Choi, Y.H.; Kim, A.; Cho, Y.-J.; Li, M.; Gunawardhana, N.; Merrell, D.S.; Ge, L. Evolutionary mechanism leading to the multi-cagA genotype in Helicobacter pylori. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tegtmeyer, N.; Wessler, S.; Backert, S. Role of the cag-pathogenicity island encoded type IV secretion system in Helicobacter pylori pathogenesis. FEBS J. 2011, 278, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Knorr, J.; Ricci, V.; Hatakeyama, M.; Backert, S. Classification of Helicobacter pylori Virulence Factors: Is CagA a Toxin or Not? Trends Microbiol. 2019. [Google Scholar] [CrossRef]
- Murata-Kamiya, N.; Kikuchi, K.; Hayashi, T.; Higashi, H.; Hatakeyama, M. Helicobacter pylori exploits host membrane phosphatidylserine for delivery, localization, and pathophysiological action of the CagA oncoprotein. Cell Host Microbe 2010, 7, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Isomoto, H.; Moss, J.; Hirayama, T. Pleiotropic actions of Helicobacter pylori vacuolating cytotoxin, VacA. Tohoku J. Exp. Med. 2010, 220, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Palframan, S.L.; Kwok, T.; Gabriel, K. Vacuolating cytotoxin A (VacA), a key toxin for Helicobacter pylori pathogenesis. Front. Cell. Infect. Microbiol. 2012, 2, 92. [Google Scholar] [CrossRef] [Green Version]
- Voss, B.J.; Gaddy, J.A.; McDonald, W.H.; Cover, T.L. Analysis of surface-exposed outer membrane proteins in Helicobacter pylori. J. Bacteriol. 2014, 196, 2455–2471. [Google Scholar] [CrossRef] [Green Version]
- Alm, R.A.; Bina, J.; Andrews, B.M.; Doig, P.; Hancock, R.E. Comparative genomics of Helicobacter pylori: Analysis of the outer membrane protein families. Infect. Immun. 2000, 68, 4155–4168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaoka, Y.; Ojo, O.; Fujimoto, S.; Odenbreit, S.; Haas, R.; Gutierrez, O.; El-Zimaity, H.M.; Reddy, R.; Arnqvist, A.; Graham, D.Y. Helicobacter pylori outer membrane proteins and gastroduodenal disease. Gut 2006, 55, 775–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mobley, H.; Island, M.D.; Hausinger, R.P. Molecular biology of microbial ureases. Microbiol. Mol. Biol. Rev. 1995, 59, 451–480. [Google Scholar] [CrossRef]
- Eaton, K.; Suerbaum, S.; Josenhans, C.; Krakowka, S. Colonization of gnotobiotic piglets by Helicobacter pylori deficient in two flagellin genes. Infect. Immun. 1996, 64, 2445–2448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, E.; Chandan, V.; Li, J.; Vinogradov, E. Lipopolysaccharide structures of Helicobacter pylori wild-type strain 26695 and 26695 HP0826: Kan mutant devoid of the O-chain polysaccharide component. Carbohydr. Res. 2011, 346, 2437–2444. [Google Scholar] [CrossRef] [PubMed]
- Thi Huyen Trang, T.; Thanh Binh, T.; Yamaoka, Y. Relationship between vacA types and development of gastroduodenal diseases. Toxins 2016, 8, 182. [Google Scholar] [CrossRef] [PubMed]
- Sgouras, D.; Tegtmeyer, N.; Wessler, S. Activity and functional importance of Helicobacter pylori virulence factors. In Helicobacter Pylori in Human Diseases: Advances in Microbiology, Infectious Diseases and Public Health; Springer: Berlin/Heidelberg, Germany, 2019; Volume 11, pp. 35–56. [Google Scholar]
- Foegeding, N.J.; Caston, R.R.; McClain, M.S.; Ohi, M.D.; Cover, T.L. An overview of Helicobacter pylori VacA toxin biology. Toxins 2016, 8, 173. [Google Scholar] [CrossRef]
- Yorita, N.; Ito, M.; Boda, T.; Kotachi, T.; Nagasaki, N.; Abuduwaili, M.; Hata, K.; Hiyama, Y.; Oka, S.; Yoshihara, M.; et al. Potential of Helicobacter pylori-uninfected signet ring cell carcinoma to invade the submucosal layer. J. Gastroenterol. Hepatol. 2019, 34, 1955–1962. [Google Scholar] [CrossRef]
- Lauren, P. The two histological main types of gastric carcinoma: Diffuse and so-called intestinal-type carcinoma: An attempt at a histo-clinical classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef]
- Ansari, S.; Gantuya, B.; Tuan, V.P.; Yamaoka, Y. Diffuse gastric cancer: A summary of analogous contributing factors for its molecular pathogenicity. Int. J. Mol. Sci. 2018, 19, 2424. [Google Scholar] [CrossRef] [Green Version]
- Pernot, S.; Voron, T.; Perkins, G.; Lagorce-Pages, C.; Berger, A.; Taieb, J. Signet-ring cell carcinoma of the stomach: Impact on prognosis and specific therapeutic challenge. World J. Gastroenterol. 2015, 21, 11428–11438. [Google Scholar] [CrossRef]
- Asaka, M.; Kimura, T.; Kato, M.; Kudo, M.; Miki, K.; Ogoshi, K.; Kato, T.; Tatsuta, M.; Graham, D.Y. Possible role of Helicobacter pylori infection in early gastric cancer development. Cancer 1994, 73, 2691–2694. [Google Scholar] [CrossRef]
- Huang, J.Q.; Sridhar, S.; Chen, Y.; Hunt, R.H. Meta-analysis of the relationship between Helicobacter pylori seropositivity and gastric cancer. Gastroenterology 1998, 114, 1169–1179. [Google Scholar] [CrossRef]
- Kikuchi, S.; Wada, O.; Nakajima, T.; Nishi, T.; Kobayashi, O.; Konishi, T.; Inaba, Y. Serum anti-Helicobacter pylori antibody and gastric carcinoma among young adults. Research Group on Prevention of Gastric Carcinoma among Young Adults. Cancer 1995, 75, 2789–2793. [Google Scholar] [CrossRef]
- Jindal, Y.; Singh, A.; Kumar, R.; Varma, K.; Misra, V.; Misra, S.P.; Dwivedi, M. Expression of alpha methylacyl CoA racemase (AMACR) in gastric adenocarcinoma and its correlation with Helicobacter pylori infection. J. Clin. Diagn. Res. JCDR 2016, 10, EC10. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Gong, E.J.; Chung, E.J.; Park, H.W.; Bae, S.E.; Kim, E.H.; Kim, J.; Do, Y.S.; Kim, T.H.; Chang, H.-S. The characteristics and prognosis of diffuse-type early gastric cancer diagnosed during health check-ups. Gut Liver 2017, 11, 807. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Kato, J.; Inoue, I.; Yoshimura, N.; Yoshida, T.; Mukoubayashi, C.; Deguchi, H.; Enomoto, S.; Ueda, K.; Maekita, T. Development of gastric cancer in nonatrophic stomach with highly active inflammation identified by serum levels of pepsinogen and Helicobacter pylori antibody together with endoscopic rugal hyperplastic gastritis. Int. J. Cancer 2012, 131, 2632–2642. [Google Scholar] [CrossRef]
- Alm, R.A.; Ling, L.-S.L.; Moir, D.T.; King, B.L.; Brown, E.D.; Doig, P.C.; Smith, D.R.; Noonan, B.; Guild, B.C.; Dejonge, B.L. Genomic-sequence comparison of two unrelated isolates of the human gastric pathogen Helicobacter pylori. Nature 1999, 397, 176–180. [Google Scholar] [CrossRef]
- Hacker, J.; Kaper, J.B. Pathogenicity islands and the evolution of microbes. Annu. Rev. Microbiol. 2000, 54, 641–679. [Google Scholar] [CrossRef] [Green Version]
- Waskito, L.A.; Yih-Wu, J.; Yamaoka, Y. The role of integrating conjugative elements in Helicobacter pylori: A review. J. Biomed. Sci. 2018, 25, 1–10. [Google Scholar] [CrossRef]
- Odenbreit, S.; Püls, J.; Sedlmaier, B.; Gerland, E.; Fischer, W.; Haas, R. Translocation of Helicobacter pylori CagA into gastric epithelial cells by type IV secretion. Science 2000, 287, 1497–1500. [Google Scholar] [CrossRef]
- Backert, S.; Tegtmeyer, N. Type IV secretion and signal transduction of Helicobacter pylori CagA through interactions with host cell receptors. Toxins 2017, 9, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backert, S.; Tegtmeyer, N.; Fischer, W. Composition, structure and function of the Helicobacter pylori cag pathogenicity island encoded type IV secretion system. Future Microbiol. 2015, 10, 955–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, M. Helicobacter pylori CagA and gastric cancer: A paradigm for hit-and-run carcinogenesis. Cell Host Microbe 2014, 15, 306–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, M. Structure and function of Helicobacter pylori CagA, the first-identified bacterial protein involved in human cancer. Proc. Jpn. Acad. Ser. B 2017, 93, 196–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, M.J.; Costa, A.M.; Costa, A.C.; Ferreira, R.M.; Sampaio, P.; Machado, J.C.; Seruca, R.; Mareel, M.; Figueiredo, C. CagA associates with c-Met, E-cadherin, and p120-catenin in a multiproteic complex that suppresses Helicobacter pylori–induced cell-invasive phenotype. J. Infect. Dis. 2009, 200, 745–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.S.; Meitner, P.; Konkin, T.A.; Cho, Y.S.; Resnick, M.B.; Moss, S.F. Altered expression of Skp2, c-Myc and p27 proteins but not mRNA after H. pylori eradication in chronic gastritis. Mod. Pathol. 2006, 19, 49–58. [Google Scholar] [CrossRef] [Green Version]
- André, A.R.; Ferreira, M.V.P.; Mota, R.M.S.; Ferrasi, A.C.; Pardini, M.I.d.M.C.; Rabenhorst, S.H.B. Gastric adenocarcinoma and Helicobacter pylori: Correlation with p53 mutation and p27 immunoexpression. Cancer Epidemiol. 2010, 34, 618–625. [Google Scholar] [CrossRef]
- Yamamoto, E.; Suzuki, H.; Takamaru, H.; Yamamoto, H.; Toyota, M.; Shinomura, Y. Role of DNA methylation in the development of diffuse-type gastric cancer. Digestion 2011, 83, 241–249. [Google Scholar] [CrossRef]
- Perri, F.; Cotugno, R.; Piepoli, A.; Merla, A.; Quitadamo, M.; Gentile, A.; Pilotto, A.; Annese, V.; Andriulli, A. Aberrant DNA methylation in non-neoplastic gastric mucosa of H. pylori infected patients and effect of eradication. Am. J. Gastroenterol. 2007, 102, 1361–1371. [Google Scholar] [CrossRef]
- Jiménez-Soto, L.; Haas, R. The CagA toxin of Helicobacter pylori: abundant production but relatively low amount translocated. Sci. Rep. 2016, 6, 23227. [Google Scholar]
- Ren, S.; Higashi, H.; Lu, H.; Azuma, T.; Hatakeyama, M. Structural basis and functional consequence of Helicobacter pylori CagA multimerization in cells. J. Biol. Chem. 2006, 281, 32344–32352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashi, K.; Murata-Kamiya, N.; Varon, C.; Mégraud, F.; Dominguez-Bello, M.G.; Hatakeyama, M. Natural variant of the Helicobacter pylori CagA oncoprotein that lost the ability to interact with PAR 1. Cancer Sci. 2014, 105, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Kiga, K.; Kersulyte, D.; Cok, J.; Hooper, C.C.; Mimuro, H.; Sanada, T.; Suzuki, S.; Oyama, M.; Kozuka-Hata, H. Attenuated CagA oncoprotein in Helicobacter pylori from Amerindians in Peruvian Amazon. J. Biol. Chem. 2011, 286, 29964–29972. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, H.; Hayashi, T.; Arisaka, F.; Senda, T.; Hatakeyama, M. Impact of structural polymorphism for the Helicobacter pylori CagA oncoprotein on binding to polarity-regulating kinase PAR1b. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef]
- Hoy, B.; Löwer, M.; Weydig, C.; Carra, G.; Tegtmeyer, N.; Geppert, T.; Schröder, P.; Sewald, N.; Backert, S.; Schneider, G. Helicobacter pylori HtrA is a new secreted virulence factor that cleaves E-cadherin to disrupt intercellular adhesion. EMBO Rep. 2010, 11, 798–804. [Google Scholar] [CrossRef] [Green Version]
- Tegtmeyer, N.; Wessler, S.; Necchi, V.; Rohde, M.; Harrer, A.; Rau, T.T.; Asche, C.I.; Boehm, M.; Loessner, H.; Figueiredo, C. Helicobacter pylori employs a unique basolateral type IV secretion mechanism for CagA delivery. Cell Host Microbe 2017, 22, 552–560.e5. [Google Scholar] [CrossRef] [Green Version]
- Kabamba, E.T.; Yamaoka, Y. Helicobacter pylori and related virulence factors for gastrointestinal diseases. In Gastric Cancer; Springer: Berlin/Heidelberg, Germany, 2019; pp. 31–50. [Google Scholar]
- Yamaoka, Y.; Kwon, D.H.; Graham, D.Y. A Mr 34,000 proinflammatory outer membrane protein (oipA) of Helicobacter pylori. Proc. Natl. Acad. Sci. USA 2000, 97, 7533–7538. [Google Scholar] [CrossRef] [Green Version]
- Leylabadlo, H.E.; Yekani, M.; Ghotaslou, R. Helicobacter pylori hopQ alleles (type I and II) in gastric cancer. Biomed. Rep. 2016, 4, 601–604. [Google Scholar] [CrossRef] [Green Version]
- Ohno, T.; Sugimoto, M.; Nagashima, A.; Ogiwara, H.; Vilaichone, R.K.; Mahachai, V.; Graham, D.Y.; Yamaoka, Y. Relationship between Helicobacter pylori hopQ genotype and clinical outcome in Asian and Western populations. J. Gastroenterol. Hepatol. 2009, 24, 462–468. [Google Scholar] [CrossRef]
- Cao, P.; Cover, T.L. Two different families of hopQ alleles in Helicobacter pylori. J. Clin. Microbiol. 2002, 40, 4504–4511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javaheri, A.; Kruse, T.; Moonens, K.; Mejías-Luque, R.; Debraekeleer, A.; Asche, C.I.; Tegtmeyer, N.; Kalali, B.; Bach, N.C.; Sieber, S.A. Helicobacter pylori adhesin HopQ engages in a virulence-enhancing interaction with human CEACAMs. Nat. Microbiol. 2016, 2, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gur, C.; Maalouf, N.; Gerhard, M.; Singer, B.B.; Emgård, J.; Temper, V.; Neuman, T.; Mandelboim, O.; Bachrach, G. The Helicobacter pylori HopQ outermembrane protein inhibits immune cell activities. Oncoimmunology 2019, 8, e1553487. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Wirth, T.; Linz, B.; Pritchard, J.K.; Stephens, M.; Kidd, M.; Blaser, M.J.; Graham, D.Y.; Vacher, S.; Perez-Perez, G.I.; et al. Traces of human migrations in Helicobacter pylori populations. Science 2003, 299, 1582–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moodley, Y.; Linz, B. Helicobacter pylori Sequences Reflect Past Human Migrations. Genome Dyn. 2009, 6, 62–74. [Google Scholar] [CrossRef]
- Moodley, Y.; Linz, B.; Bond, R.P.; Nieuwoudt, M.; Soodyall, H.; Schlebusch, C.M.; Bernhoft, S.; Hale, J.; Suerbaum, S.; Mugisha, L.; et al. Age of the association between Helicobacter pylori and man. PLoS Pathog. 2012, 8, e1002693. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Tomb, J.-F.; White, O.; Kerlavage, A.R.; Clayton, R.A.; Sutton, G.G.; Fleischmann, R.D.; Ketchum, K.A.; Klenk, H.P.; Gill, S.; Dougherty, B.A. The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature 1997, 388, 539–547. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- Choi, I.J.; Fujimoto, S.; Yamauchi, K.; Graham, D.Y.; Yamaoka, Y. Helicobacter pylori environmental interactions: Effect of acidic conditions on H. pylori-induced gastric mucosal interleukin-8 production. Cell. Microbiol. 2007, 9, 2457–2469. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virulence Factor | Gene | Start | End | % Coverage | % Identity | Characteristics |
|---|---|---|---|---|---|---|
| CagA | hp0547 | 750,676 | 754,221 | 99.49 | 94.58 | 1 copy |
| VacA | hp0887 | 1,114,652 | 1,118,533 | 99.69 | 91.78 | s1m1 |
| HtrA | hp1018 | 639,400 | 640,830 | 99.93 | 95.53 | 1 copy |
| CGT | hp0421 | 1,272,696 | 1,273,861 | 99.66 | 95.20 | 1 copy |
| GGT | hp1118 | 1,371,288 | 1,372,991 | 100.00 | 95.31 | 1 copy |
| DupA | hp0441 | − | − | − | − | Absent |
| IceA | hp1209 | − | − | − | − | Absent |
| OipA/HopH | hp0638 | 852,806 | 853,731 | 99.89 | 93.85 | 1 copy in “ON” status |
| AlpA/HopC | hp0912 | 1,143,117 | 1,144,676 | 100.00 | 94.49 | 1 copy |
| AlpB/HopB | hp0913 | 1,144,698 | 1,146,287 | 99.81 | 94.35 | 1 copy |
| HopQ/Omp27 | hp1177 | 1,434,514 | 1,436,428 | 99.17 | 88.18 | 1 copy hopQ type1 allele |
| BabA/HopS | hp1243 | 1,124,618; 1,507,238 | 1,126,840; 1,509,404 | 99.19; 96.69 | 87.64; 87.32 | 2 copies |
| BabB/HopT | hp0896 | − | − | − | − | Absent |
| BabC/HopU | hp0317 | − | − | − | − | Absent |
| HomA | hp0710 | 937,597; 1,173,611 | 93,9589; 1,175,576 | 98.03; 98.29 | 85.96; 90.64 | 2 copies |
| SabA/Omp17 | hp0725 | 952,657 | 954,511 | 97.99 | 89.10 | 1 copy |
| SabB/HopO | hp0722 | 948,177 | 949,975 | 98.30 | 88.06 | 1 copy |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mwangi, C.; Njoroge, S.; Tshibangu-Kabamba, E.; Moloo, Z.; Rajula, A.; Devani, S.; Matsumoto, T.; Nyerere, K.; Kariuki, S.; Revathi, G.; et al. Whole Genome Sequencing Reveals Virulence Potentials of Helicobacter pylori Strain KE21 Isolated from a Kenyan Patient with Gastric Signet Ring Cell Carcinoma. Toxins 2020, 12, 556. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12090556
Mwangi C, Njoroge S, Tshibangu-Kabamba E, Moloo Z, Rajula A, Devani S, Matsumoto T, Nyerere K, Kariuki S, Revathi G, et al. Whole Genome Sequencing Reveals Virulence Potentials of Helicobacter pylori Strain KE21 Isolated from a Kenyan Patient with Gastric Signet Ring Cell Carcinoma. Toxins. 2020; 12(9):556. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12090556
Chicago/Turabian StyleMwangi, Catherine, Stephen Njoroge, Evariste Tshibangu-Kabamba, Zahir Moloo, Allan Rajula, Smita Devani, Takashi Matsumoto, Kimang’a Nyerere, Samuel Kariuki, Gunturu Revathi, and et al. 2020. "Whole Genome Sequencing Reveals Virulence Potentials of Helicobacter pylori Strain KE21 Isolated from a Kenyan Patient with Gastric Signet Ring Cell Carcinoma" Toxins 12, no. 9: 556. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12090556