Mechanisms of Tolerance and Resistance to Chlorhexidine in Clinical Strains of Klebsiella pneumoniae Producers of Carbapenemase: Role of New Type II Toxin-Antitoxin System, PemIK

 , , , , , ,
, , , , , ,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Results Subsection
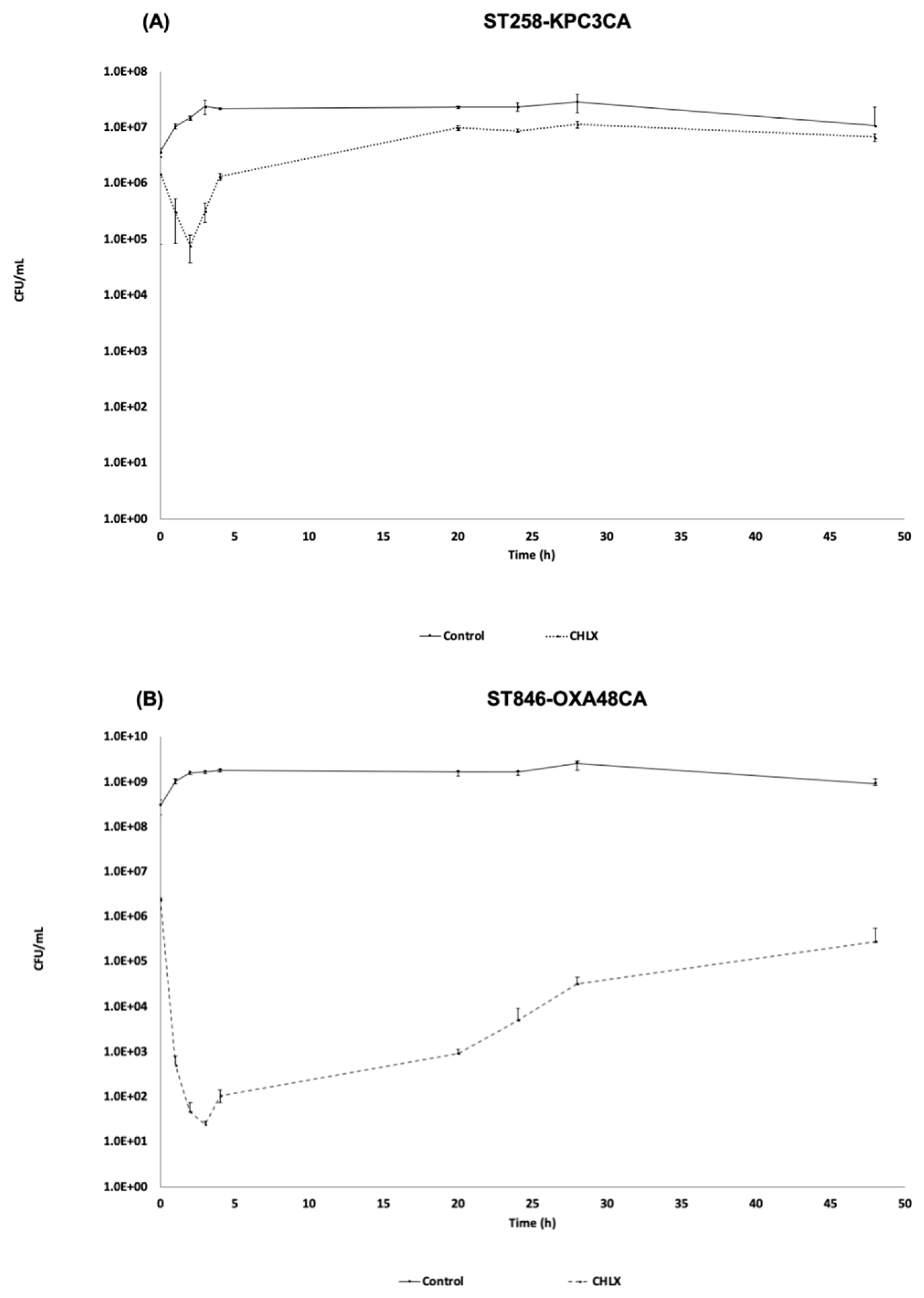
2.1.1. Time-Killing Curve in the Presence of CHLX (10 × MIC)
2.1.2. Transcriptomic Study
2.1.3. Antimicrobial Susceptibly Testing
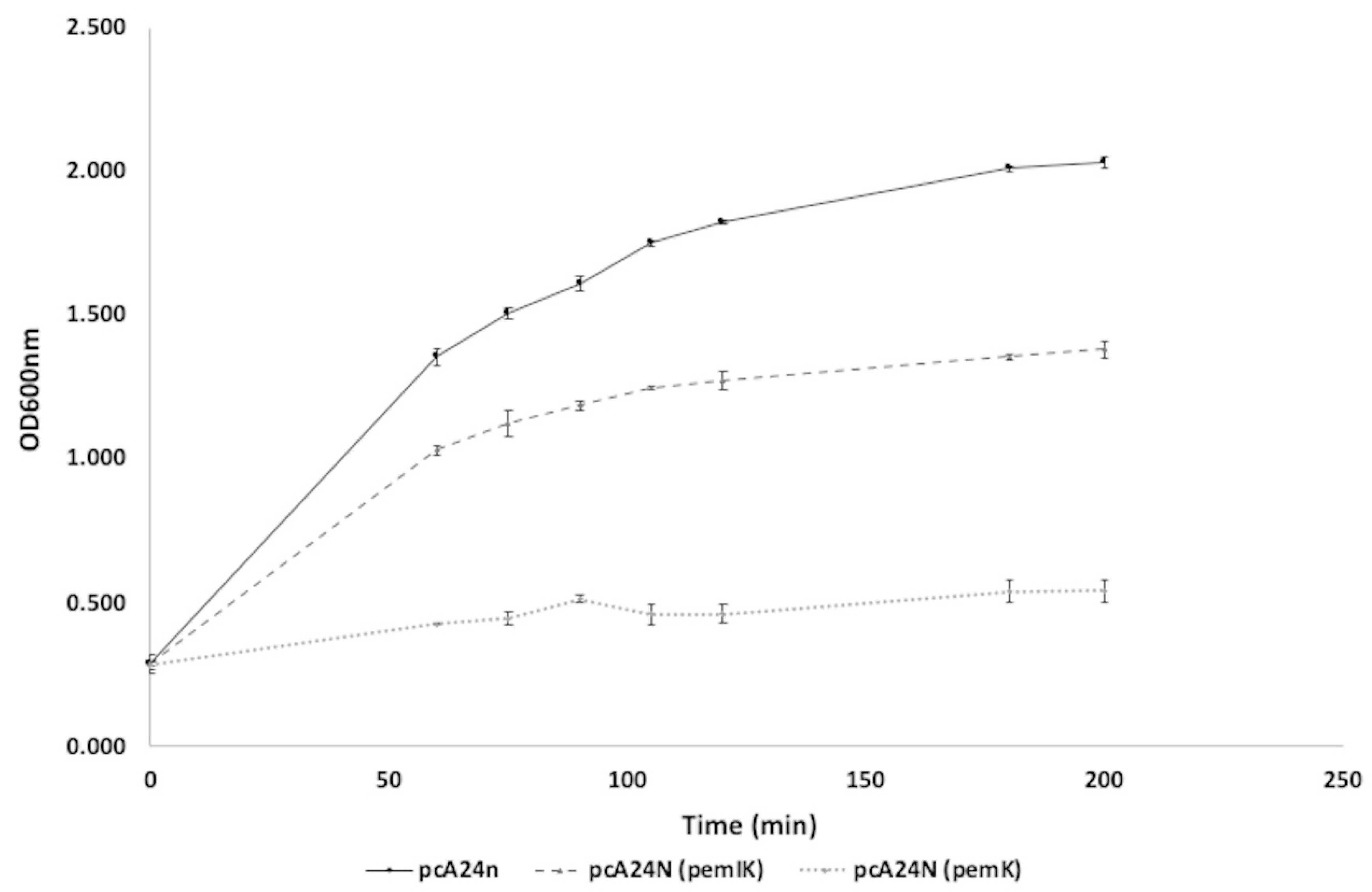
2.1.4. Characterization of the New TA System, PemI/PemK, Present in a Plasmid in the Strain ST846-OXA48CA
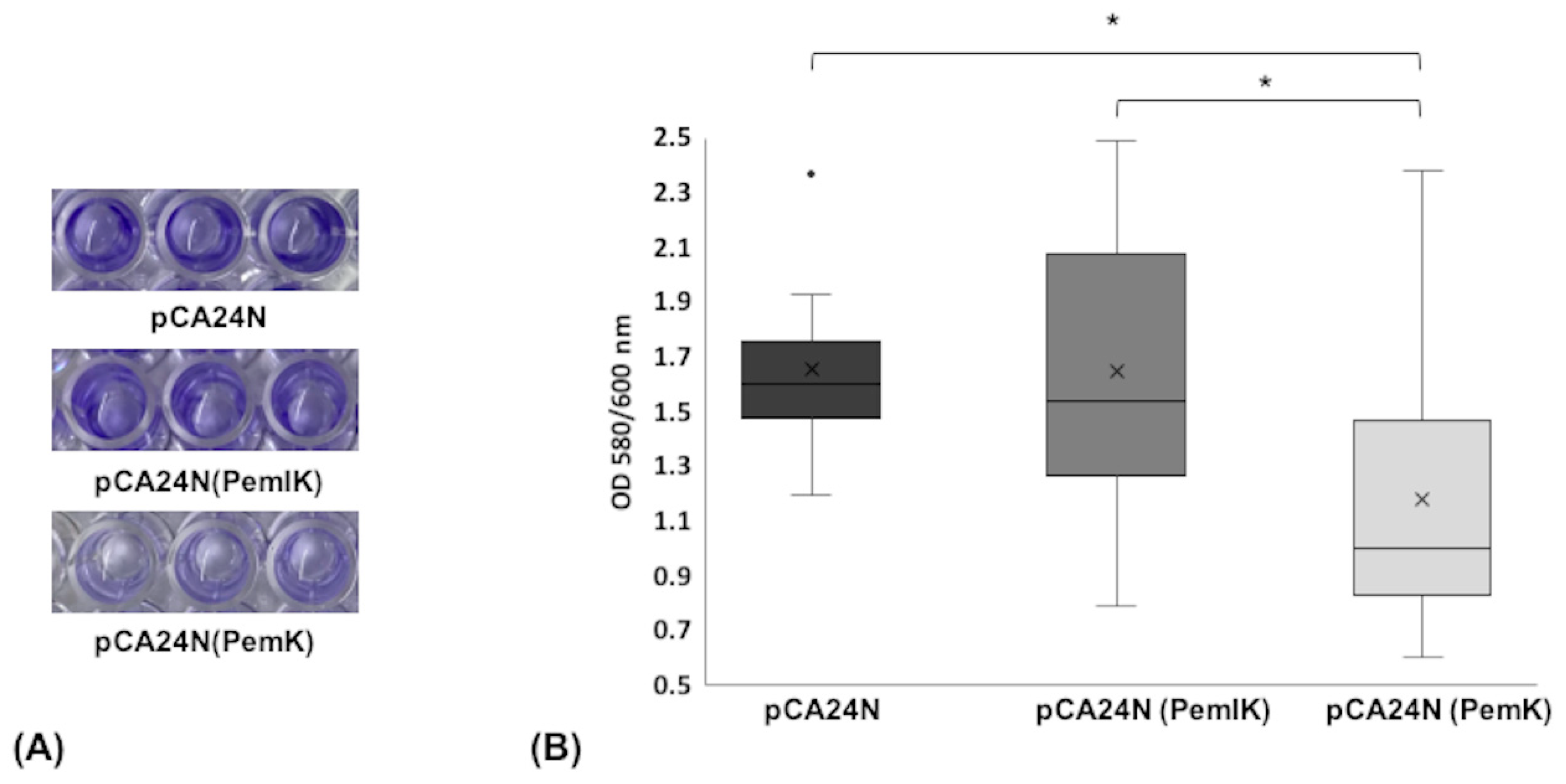
2.1.5. Biofilm Formation Assay
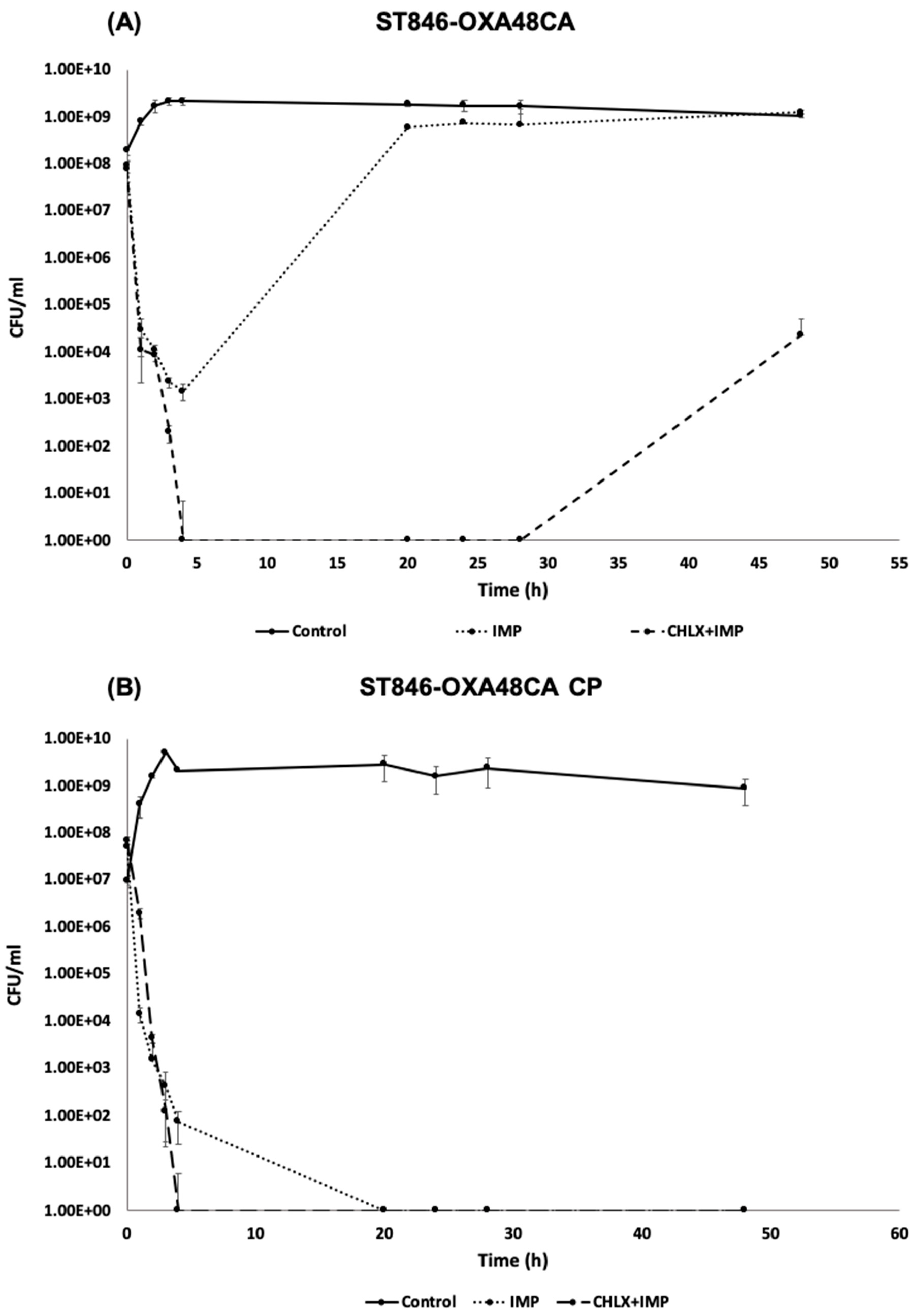
2.1.6. Time-Killing Curve in the Presence of Imipenem or in Combination with Chlorhexidine for ST846-OXA48CA and ST846-OXA48CA CP
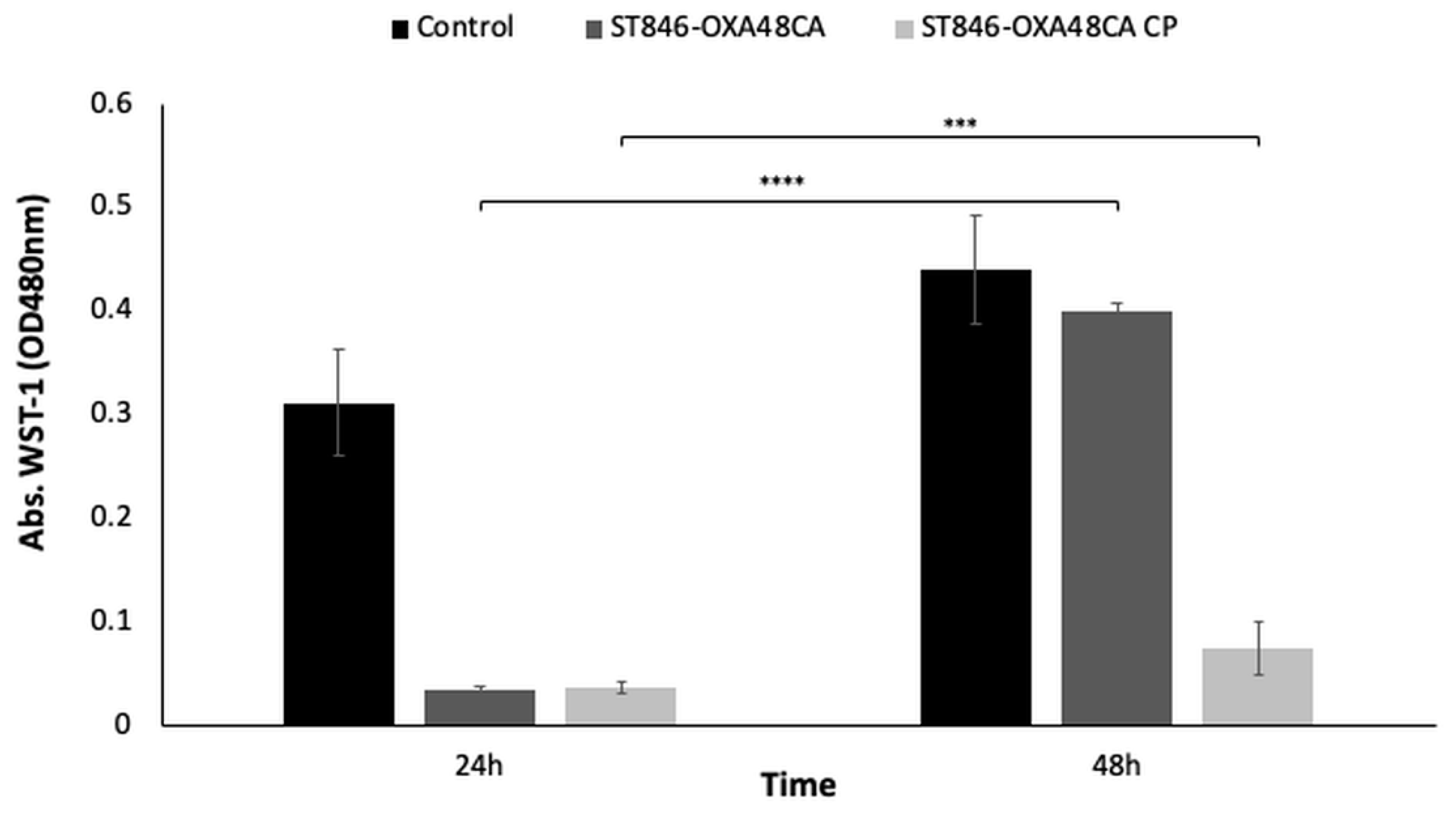
2.1.7. Enzymatic Analysis Using the Cell Proliferation Reagent WST-1
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bacterial Strains and CHLX Adaptation
5.2. Time-Killing Curve
5.3. Transcriptomic Study
5.4. Antimicrobial Susceptibility Test
5.5. Construction of pCA24N (pemIK) and pCA24N (pemK)
5.6. Toxicity Assay
5.7. Biofilm Formation Assay
5.8. Enzymatic Assay Using the Cell Proliferation Reagent WST-1
5.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.E.; Walsh, T.R. Toxin-antitoxin systems and their role in disseminating and maintaining antimicrobial resistance. FEMS Microbiol. Rev. 2017, 41, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Santajit, S.; Indrawattana, N. Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. BioMed Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [Green Version]
- Cadavid, E.; Robledo, S.M.; Quiñones, W.; Echeverri, F. Induction of Biofilm Formation in. Antibiotics 2018, 7, 103. [Google Scholar] [CrossRef] [Green Version]
- Cieplik, F.; Jakubovics, N.S.; Buchalla, W.; Maisch, T.; Hellwig, E.; Al-Ahmad, A. Resistance Toward Chlorhexidine in Oral Bacteria—Is There Cause for Concern? Front. Microbiol. 2019, 10, 587. [Google Scholar] [CrossRef] [Green Version]
- Hashemi, M.M.; Holden, B.S.; Coburn, J.; Taylor, M.F.; Weber, S.; Hilton, B.; Zaugg, A.L.; McEwan, C.; Carson, R.; Andersen, J.L.; et al. Proteomic Analysis of Resistance of Gram-Negative Bacteria to Chlorhexidine and Impacts on Susceptibility to Colistin, Antimicrobial Peptides, and Ceragenins. Front. Microbiol. 2019, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Kampf, G. Acquired resistance to chlorhexidine—Is it time to establish an ‘antiseptic stewardship’ initiative? J. Hosp. Infect. 2016, 94, 213–227. [Google Scholar] [CrossRef]
- Venter, H.; Henningsen, M.L.; Begg, S.L. Antimicrobial resistance in healthcare, agriculture and the environment: The biochemistry behind the headlines. Essays Biochem. 2017, 61, 1–10. [Google Scholar] [CrossRef]
- Wand, M.E.; Bock, L.J.; Bonney, L.C.; Sutton, J.M. Mechanisms of Increased Resistance to Chlorhexidine and Cross-Resistance to Colistin following Exposure of Klebsiella pneumoniae Clinical Isolates to Chlorhexidine. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Trastoy, R.; Manso, T.; Fernández-García, L.; Blasco, L.; Ambroa, A.; Pérez Del Molino, M.L.; Bou, G.; García-Contreras, R.; Wood, T.K.; Tomás, M. Mechanisms of Bacterial Tolerance and Persistence in the Gastrointestinal and Respiratory Environments. Clin. Microbiol. Rev. 2018, 31. [Google Scholar] [CrossRef] [Green Version]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, L.; Fernandez-Cuenca, F.; Blasco, L.; López-Rojas, R.; Ambroa, A.; Lopez, M.; Pascual, Á.; Bou, G.; Tomás, M. Relationship between Tolerance and Persistence Mechanisms in Acinetobacter baumannii Strains with AbkAB Toxin-Antitoxin System. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, D.; Bedzyk, L.A.; Thomas, S.M.; Ye, R.W.; Wood, T.K. Gene expression in Escherichia coli biofilms. Appl. Microbiol. Biotechnol. 2004, 64, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Wang, X.; Ma, Q.; Zhang, X.S.; Wood, T.K. Toxin-antitoxin systems in Escherichia coli influence biofilm formation through YjgK (TabA) and fimbriae. J. Bacteriol. 2009, 191, 1258–1267. [Google Scholar] [CrossRef] [Green Version]
- Moyed, H.S.; Bertrand, K.P. hipA, a newly recognized gene of Escherichia coli K-12 that affects frequency of persistence after inhibition of murein synthesis. J. Bacteriol. 1983, 155, 768–775. [Google Scholar] [CrossRef] [Green Version]
- Korch, S.B.; Henderson, T.A.; Hill, T.M. Characterization of the hipA7 allele of Escherichia coli and evidence that high persistence is governed by (p)ppGpp synthesis. Mol. Microbiol. 2003, 50, 1199–1213. [Google Scholar] [CrossRef]
- Bokinsky, G.; Baidoo, E.E.; Akella, S.; Burd, H.; Weaver, D.; Alonso-Gutierrez, J.; García-Martín, H.; Lee, T.S.; Keasling, J.D. HipA-triggered growth arrest and β-lactam tolerance in Escherichia coli are mediated by RelA-dependent ppGpp synthesis. J. Bacteriol. 2013, 195, 3173–3182. [Google Scholar] [CrossRef] [Green Version]
- Page, R.; Peti, W. Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 2016, 12, 208–214. [Google Scholar] [CrossRef]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial persister cell formation and dormancy. Appl. Environ. Microbiol. 2013, 79, 7116–7121. [Google Scholar] [CrossRef] [Green Version]
- Ogura, T.; Hiraga, S. Mini-F plasmid genes that couple host cell division to plasmid proliferation. Proc. Natl. Acad. Sci. USA 1983, 80, 4784–4788. [Google Scholar] [CrossRef] [Green Version]
- Kamruzzaman, M.; Iredell, J. A ParDE-family toxin antitoxin system in major resistance plasmids of Enterobacteriaceae confers antibiotic and heat tolerance. Sci. Rep. 2019, 9, 9872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wood, T.K. Toxin-antitoxin systems influence biofilm and persister cell formation and the general stress response. Appl. Environ. Microbiol. 2011, 77, 5577–5583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecota, D.C.; Wood, T.K. Exclusion of T4 phage by the hok/sok killer locus from plasmid R1. J. Bacteriol. 1996, 178, 2044–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazan, R.; Engelberg-Kulka, H. Escherichia coli mazEF-mediated cell death as a defense mechanism that inhibits the spread of phage P1. Mol. Genet. Genom. 2004, 272, 227–234. [Google Scholar] [CrossRef]
- Song, S.; Wood, T.K. Post-segregational Killing and Phage Inhibition Are Not Mediated by Cell Death Through Toxin/Antitoxin Systems. Front. Microbiol. 2018, 9, 814. [Google Scholar] [CrossRef] [Green Version]
- Mathers, A.J.; Peirano, G.; Pitout, J.D. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [Green Version]
- Domokos, J.; Damjanova, I.; Kristof, K.; Ligeti, B.; Kocsis, B.; Szabo, D. Multiple Benefits of Plasmid-Mediated Quinolone Resistance Determinants in. Front. Microbiol. 2019, 10, 157. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lord, D.M.; Cheng, H.Y.; Osbourne, D.O.; Hong, S.H.; Sanchez-Torres, V.; Quiroga, C.; Zheng, K.; Herrmann, T.; Peti, W.; et al. A new type V toxin-antitoxin system where mRNA for toxin GhoT is cleaved by antitoxin GhoS. Nat. Chem. Biol. 2012, 8, 855–861. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Garcia, L.; Kim, J.S.; Tomas, M.; Wood, T.K. Toxins of toxin/antitoxin systems are inactivated primarily through promoter mutations. J. Appl. Microbiol. 2019, 127, 1859–1868. [Google Scholar] [CrossRef]
- Präbst, K.; Engelhardt, H.; Ringgeler, S.; Hübner, H. Basic Colorimetric Proliferation Assays: MTT, WST, and Resazurin. Methods Mol. Biol. 2017, 1601, 1–17. [Google Scholar] [CrossRef]
- Gilbert, P.; Moore, L.E. Cationic antiseptics: Diversity of action under a common epithet. J. Appl. Microbiol. 2005, 99, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Keren, I.; Shah, D.; Spoering, A.; Kaldalu, N.; Lewis, K. Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli. J. Bacteriol. 2004, 186, 8172–8180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tashiro, Y.; Kawata, K.; Taniuchi, A.; Kakinuma, K.; May, T.; Okabe, S. RelE-mediated dormancy is enhanced at high cell density in Escherichia coli. J. Bacteriol. 2012, 194, 1169–1176. [Google Scholar] [CrossRef]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef]
- Agarwal, S.; Bhatnagar, R. Identification and characterization of a novel toxin-antitoxin module from Bacillus anthracis. FEBS Lett. 2007, 581, 1727–1734. [Google Scholar] [CrossRef] [Green Version]
- Kędzierska, B.; Hayes, F. Emerging Roles of Toxin-Antitoxin Modules in Bacterial Pathogenesis. Molecules 2016, 21, 790. [Google Scholar] [CrossRef] [Green Version]
- Martínez, L.C.; Vadyvaloo, V. Mechanisms of post-transcriptional gene regulation in bacterial biofilms. Front. Cell. Infect. Microbiol. 2014, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- González Barrios, A.F.; Zuo, R.; Hashimoto, Y.; Yang, L.; Bentley, W.E.; Wood, T.K. Autoinducer 2 controls biofilm formation in Escherichia coli through a novel motility quorum-sensing regulator (MqsR, B3022). J. Bacteriol. 2006, 188, 305–316. [Google Scholar] [CrossRef] [Green Version]
- García-Contreras, R.; Zhang, X.S.; Kim, Y.; Wood, T.K. Protein translation and cell death: The role of rare tRNAs in biofilm formation and in activating dormant phage killer genes. PLoS ONE 2008, 3, e2394. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Mandell, J.B.; Donegan, N.P.; Cheung, A.L.; Ma, W.; Rothenberger, S.; Shanks, R.M.Q.; Richardson, A.R.; Urish, K.L. The Toxin-Antitoxin MazEF Drives Staphylococcus aureus Biofilm Formation, Antibiotic Tolerance, and Chronic Infection. mBio 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteban-Cantos, A.; Aracil, B.; Bautista, V.; Ortega, A.; Lara, N.; Saez, D.; Fernández-Romero, S.; Pérez-Vázquez, M.; Navarro, F.; Grundmann, H.; et al. The Carbapenemase-Producing Klebsiella pneumoniae Population Is Distinct and More Clonal than the Carbapenem-Susceptible Population. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- El-Mansi, M.; Anderson, K.J.; Inche, C.A.; Knowles, L.K.; Platt, D.J. Isolation and curing of the Klebsiella pneumonia large indigenous plasmid using sodium dodecyl sulphate. Res. Microbiol. 2000, 151, 201–208. [Google Scholar] [CrossRef]
- Kitagawa, M.; Ara, T.; Arifuzzaman, M.; Ioka-Nakamichi, T.; Inamoto, E.; Toyonaga, H.; Mori, H. Complete set of ORF clones of Escherichia coli ASKA library (a complete set of E. coli K-12 ORF archive): Unique resources for biological research. DNA Res. 2005, 12, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standard Institute. Performance Standards for Antimicrobial Sucesceptibilty Testing, 28th ed.; CLSI suplement M100; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Wood, T.L.; Wood, T.K. The HigB/HigA toxin/antitoxin system of Pseudomonas aeruginosa influences the virulence factors pyochelin, pyocyanin, and biofilm formation. Microbiologyopen 2016, 5, 499–511. [Google Scholar] [CrossRef]
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction. Biotechnol. Annu. Rev. 2005, 11, 127–152. [Google Scholar] [CrossRef]
- Tsukatani, T.; Suenaga, H.; Higuchi, T.; Akao, T.; Ishiyama, M.; Ezoe, K.; Matsumoto, K. Colorimetric cell proliferation assay for microorganisms in microtiter plate using water-soluble tetrazolium salts. J. Microbiol. Methods 2008, 75, 109–116. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mechanism | Gene a | Description | ST258-KPC3CA | ST846-OXA48CA | ||
|---|---|---|---|---|---|---|
| Log2 FoldChange | ID Gene | Log2 FoldChange | ID Gene | |||
| Transporter | smvA | Methyl viologen protein (cationic biocide resistance) | 3.635 | HGAILKPD_00917 | 1.209 | EMNICGIE_00134 |
| actP | Acetate permease ActP (cation/acetate symporter) | 2.724 | HGAILKPD_00571 | 0.649 | EMNICGIE_02128 | |
| csbX | MFS superfamily | 2.549 | HGAILKPD_04325 | 0.618 | EMNICGIE_04796 | |
| lldP | L-lactate permease | 2.486 | HGAILKPD_04496 | 0.085 | EMNICGIE_00243 | |
| cysW | Ferric iron ABC transporter | 2.181 | HGAILKPD_02877 | −0.266 | EMNICGIE_02673 | |
| potA | ABC transporter | 1.749 | HGAILKPD_02785 | −0.351 | EMNICGIE_03352 | |
| pmrD | Signal transduction protein PmrD (colistin resistance) | - | - | 2.360 | EMNICGIE_04427 | |
| pmrK | Polymyxin resistance protein PmrK (colistin resistance) | - | - | 1.570 | EMNICGIE_02839 | |
| ATP metabolism | atpD | ATP synthase beta chain | −0.209 | HGAILKPD_02375 | −0.232 | EMNICGIE_00435 |
| TA systems | ortT | Orphan toxin OrtT | 0.731 | HGAILKPD_02791 | 0.727 | EMNICGIE_00095 |
| pemI | Programmed cell death antitoxin PemI | - | - | −0.100 | EMNICGIE_05097 | |
| pemK | Programmed cell death toxin PemK | - | - | −0.302 | EMNICGIE_05098 | |
| (p)ppGpp | gppA | Guanosine-5′-triphosphate,3′-diphosphate pyrophosphatase | −0.765 | HGAILKPD_02586 | −0.399 | EMNICGIE_03280 |
| ROS response | cydA | Cytochrome d ubiquinol oxidase subunit I | 1.318 | HGAILKPD_03209 | 0.441 | EMNICGIE_03423 |
| cybB | Cytochrome b561 | 0.456 | HGAILKPD_02756 | 0.196 | EMNICGIE_00060 | |
| SOS system | yedK | Putative SOS response-associated peptidase YedK | 1.117 | HGAILKPD_04848 | 0.003 | EMNICGIE_04152 |
| yebG | DNA damage-inducible gene in SOS regulon | 0.722 | HGAILKPD_02193 | 0.455 | EMNICGIE_04716 | |
| MIC (μg/mL) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | CHLX | CIP | TGC | TOB | IMP | MRP | GEN | CAZ | TZP | SAM | NET | DOX | AMK | MIN | CST |
| ST258-KPC3 | 9.8 | >32 | 2 | 64 | 4 | 8 | 4 | >32 | >32 | 1024 | 128 | 2 | 16 | 4 | 0.25 |
| ST258-KPC3CA | 39.1 | >32 | 2 | 64 | 4 | 8 | 4 | >32 | >32 | 1024 | 128 | 2 | 16 | 4 | 0.25 |
| ST846-OXA48 | 19.5 | 8 | 8 | 32 | 16 | >32 | 32 | >32 | >32 | 128 | 16 | 8 | 2 | 8 | 0.5 |
| ST846-OXA48CA | 78.2 | 8 | 8 | 32 | 16 | >32 | 32 | >32 | >32 | 128 | 16 | 8 | 2 | 8 | 16 |
| Strain or Plasmid | Main Characteristics | Source or Reference |
|---|---|---|
| ST258-KPC3 | K. pneumoniae high-risk clone carrying β-lactamase KPC3 | This study |
| ST258-KPC3 CA | K. pneumoniae high-risk clone carrying β-lactamase KPC3 adapted to CHLX | This study |
| ST846-OXA48 | K. pneumoniae low-risk clone carrying β-lactamase OXA48 | This study |
| ST846-OXA48 CA | K. pneumoniae low-risk clone carrying β-lactamase OXA48 adapted to CHLX | This study |
| ST846-OXA48 CA CP | K. pneumoniae low-risk clone carrying β-lactamase OXA48 adapted to CHLX and cured plasmid strain | This study |
| pCA24N | Expression plasmid CmR, LacIq | [45] |
| pCA24N (pemIK) | Expression plasmid pCA24N with the TA systems pemIK | This study |
| pCA24N (pemK) | Expression plasmid pCA24N with the TA systems pemIK | This study |
| Primer Name | Sequences | Sense | Reference |
|---|---|---|---|
| Oligonucleotide for clonation | |||
| PemI_Fow(BseRI) | GAGGAGAAATTAACTATCATGCATACCACTCGACTG | 5′-3′ | This study |
| PemK_Fow(BseRI) | GAGGAGAAATTAACTATCATGGAAAGAGGGGAAATC | 5′-3′ | This study |
| PemK_Rev(NotI) | ATAAGAATGCGGCCGCCGCTCAGGTCAGGATGGTGGC | 5′-3′ | This study |
| Oligonucleotide for sequencing | |||
| pCA24N Up | GCCCTTTCGTCTTCAC | 5′-3′ | This study |
| pCA24N Down | GAACTCCATCTGGATTTGTT | 5′-3′ | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bleriot, I.; Blasco, L.; Delgado-Valverde, M.; Gual-de-Torrella, A.; Ambroa, A.; Fernandez-Garcia, L.; Lopez, M.; Oteo-Iglesias, J.; Wood, T.K.; Pascual, A.; et al. Mechanisms of Tolerance and Resistance to Chlorhexidine in Clinical Strains of Klebsiella pneumoniae Producers of Carbapenemase: Role of New Type II Toxin-Antitoxin System, PemIK. Toxins 2020, 12, 566. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12090566
Bleriot I, Blasco L, Delgado-Valverde M, Gual-de-Torrella A, Ambroa A, Fernandez-Garcia L, Lopez M, Oteo-Iglesias J, Wood TK, Pascual A, et al. Mechanisms of Tolerance and Resistance to Chlorhexidine in Clinical Strains of Klebsiella pneumoniae Producers of Carbapenemase: Role of New Type II Toxin-Antitoxin System, PemIK. Toxins. 2020; 12(9):566. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12090566
Chicago/Turabian StyleBleriot, Ines, Lucia Blasco, Mercedes Delgado-Valverde, Ana Gual-de-Torrella, Anton Ambroa, Laura Fernandez-Garcia, Maria Lopez, Jesus Oteo-Iglesias, Thomas K. Wood, Alvaro Pascual, and et al. 2020. "Mechanisms of Tolerance and Resistance to Chlorhexidine in Clinical Strains of Klebsiella pneumoniae Producers of Carbapenemase: Role of New Type II Toxin-Antitoxin System, PemIK" Toxins 12, no. 9: 566. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12090566