1. Introduction
Aquaculture is the fastest growing food production industry in the world, and by 2030 it is expected to provide 60 percent of the fish available for human consumption [
1]. In aquaculture, plant-based protein alternatives are used to replace or partially replace fish meal since exclusive use of fish meal is not sustainable [
2,
3,
4]. However, plant-based ingredients are easily contaminated with mycotoxins, which increase the health risks to fish [
5,
6]. Aflatoxin B
1 (AFB
1), the secondary metabolite of fungi
Aspergillus parasiticus and
A. flavus [
7], is one of the most harmful mycotoxins [
8]. In poultry and livestock, the detrimental effects of AFB
1 include low productivity, high mortality of offspring, anorexia, poor growth, immune dysfunction, and AFB
1 residues in edible animal parts [
9,
10,
11,
12,
13,
14,
15]. In fish, there were studies of various species exposed to AFB
1. Most of these studies concentrated on the impact on growth performance, liver lesions and immunosuppression induced by dietary AFB
1. It had been reported that when sea bass (
Dicentrarchus labrax) were fed a diet with 18 μg AFB
1/kg of body weight, adverse effects (liver lesions and AFB
1 residues in musculature) were induced [
16]. Studies on rainbow trout (
Oncorhynchus mykiss) indicated that toxic effects of AFB
1 could be induced when fish were fed more than 0.05 μg AFB
1/kg diet [
17,
18,
19,
20,
21,
22]. There was also plenty of research in nile tilapia (
Oreochromis niloticus) [
23,
24,
25,
26], the poor growth performance was caused by more than 250 μg AFB
1/kg in diet. Other relevant studies also included gibel carp (
Carassius auratus gibelio) [
27,
28,
29], grass carp (
Ctenopharyngodon idella) [
30], rohu (
Labeo rohita) [
31,
32,
33], red tilapia (
O. niloticus ×
O. mossambicus) [
34], channel catfish (
Ictalurus punctatus) [
35], tambaqui fingerlings (
Colossoma macropomum) [
36], beluga (
Huso huso) [
37] and Thai koi (
Anabas testudineus) [
38]. Usually the toxic effects of AFB
1 occurred when the dose was greater than 100 μg/kg diet.
AFB
1 is classified as Group 1 carcinogen of hepatocellular carcinoma to human by International Agency for Research on Cancer [
39]. Consequently, food consumption of AFB
1 presents a serious risk to human health [
40,
41,
42]. Few studies showed the results of AFB
1 residues in musculature in fish such as sea bass [
16], gibel carp [
27], tambaqui [
36] and Thai koi [
38], whereas other studies did not detect AFB
1 tissue residues in nile tilapia [
26] and red tilapia [
34]. The disparities observed in these studies show the potential differences in responses and absorption/metabolization processes amongst fish species following AFB
1 exposure. In addition, information on the effect of AFB
1 on intestinal microbiota of fish is limited. Therefore, a more complete comprehensive understanding of the effects of AFB
1 in fish is necessary.
AFB
1 is stable and difficult to remove from contaminated feed; therefore, one of the strategies is to decrease its bioavailability [
43]. Nowadays, adsorbing agents which could prevent AFB
1 from being absorbed by the intestine are widely studied and used [
41,
43]. The studies of these products, including hydrated sodium calcium aluminosilicate, activated carbon, zeolites and yeast cell wall, etc., have been reviewed comprehensively [
43,
44,
45,
46,
47,
48]. Due to the negative effects of some of the inorganic adsorbents, such as adsorption of micronutrients [
44,
45], high inclusion rates [
49] and limited adsorbing capacity to multiple mycotoxins [
48,
50], research has shifted to focus on composite-type mycotoxin adsorbents [
43,
51,
52,
53]. Yeast cell wall extract (YCWE) is an adsorbent that contains yeast cell wall, beer yeast powder, calcium carbonate and hydrated sodium calcium aluminosilicate, which have shown favorable effects in livestock and poultry challenged with mycotoxin exposure [
54,
55,
56,
57,
58,
59,
60,
61,
62,
63].
Turbot (
Scophthalmus maximus) is an important commercial marine species in aquaculture. Recently, plant-based ingredients have been more widely used in the feeds of marine fish [
2], with some inclusion levels higher than 50% of the total feed ingredients [
64,
65,
66,
67]. Research about the effects of AFB
1 and mycotoxin adsorbents on turbot health and performance is lacking. Therefore, this study was aimed to investigate the impacts of AFB
1 in turbot and evaluate the effects of YCWE on turbot fed AFB
1 contaminated diets.
3. Discussion
In the present study, dietary AFB
1 (20 and 500 μg/kg) did not remarkably affect the growth performance of turbot in a 67-day feeding trial. The adverse effects of higher level of AFB
1 in diet on growth performance had been reported in several fish species. In nile tilapia, diets with AFB
1 (2000 or 4000 μg/kg) remarkably reduced the weight gain (WG), FE, and the content of crude lipid [
23], while results reported by Tuan et al. [
25] and Deng et al. [
26] demonstrated that WG, FI and FE were significantly reduced by 250 μg/kg or higher dietary AFB
1. A similar result was also showed in tambaqui (500 μg/kg or higher level of AFB
1) [
36]. However, previous studies found that a diet with low levels of AFB
1 could reduce the WGR, SGR and FI of grass carp (less than 147 μg/kg AFB
1) [
30]. Beluga fed diets with 75 or 100 μg/kg AFB
1 affected WG and FE but not the SGR [
37]. The effects of dietary AFB
1 on the growth performance of fish is closely tied to the level of AFB
1 in diet but is also dependent on the fish species, the development stage, the environment and the length of feeding terms.
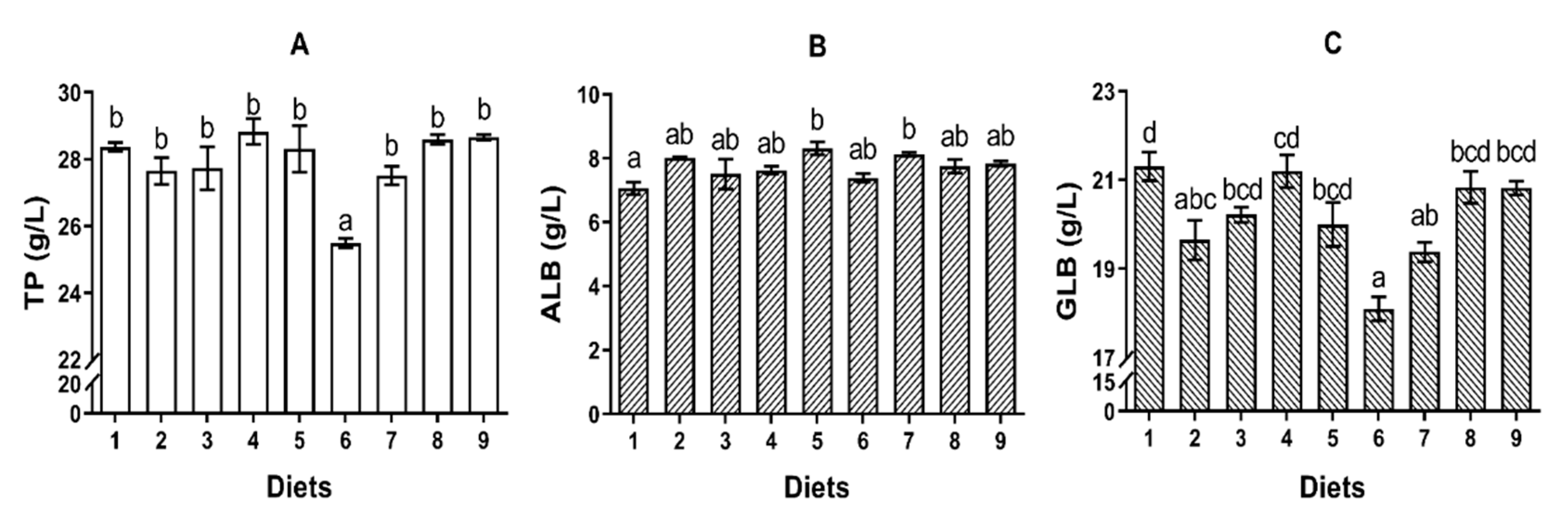
It has been identified that dietary AFB
1 could induce immunosuppressive response in aquatic animals, such as sea bass [
16], grass carp [
30], rohu [
32], nile tilapia [
68,
69] and pacific white shrimp [
70]. The level of TP and GLB can reflect protein synthesis capacity and immunity [
32,
71,
72]. C3 and C4 are the key components of both classical and lectin pathways responsible for various immune effector functions [
73]. In the current study, the concentrations of TP and GLB, as well as the concentrations of C3 and C4 in serum, were reduced by Diet 6, which suggested an immunosuppressive effect. It has been reported that the reduction of TP might be attributed to the hepatocellular damage [
32], and reduction of GLB might be resulting from lymphocytolysis [
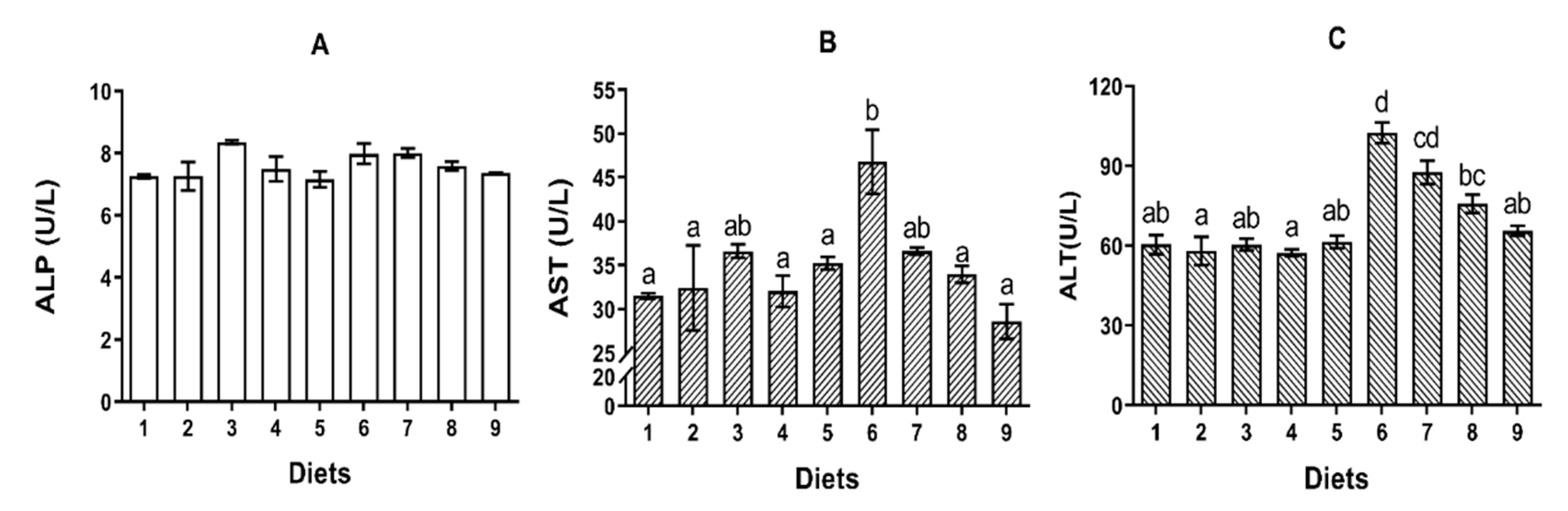
74]. Generally, AST and ALT are recognized as biomarkers to identify the hepatic functions and cell membrane permeability. In the present study, turbot fed Diet 6 had higher activities of AST and ALT in serum indicating that AFB
1 might cause hepatocellular damage in turbot. Similar results had been observed in other aquatic animals such as sea bass [
16], nile tilapia [
26], gibel carp [
29] and pacific white shrimp [
75]. The reduction of the concentrations of TG and T-CHO in serum induced by Diet 6 was similar when compared to previous studies performed in broiler chicks and ducks [
76,
77]. Due to the hepatocellular damage, the synthesis of TG and T-CHO was decreased, which was consistent with the lower concentrations of TP and GLB of serum as well. On the other hand, AFB
1 requires metabolic activation by the cytochrome p450 enzymes system to generate AFB
1-
exo-8, 9-epoxide (AFBO), which can exert cytotoxic effects [
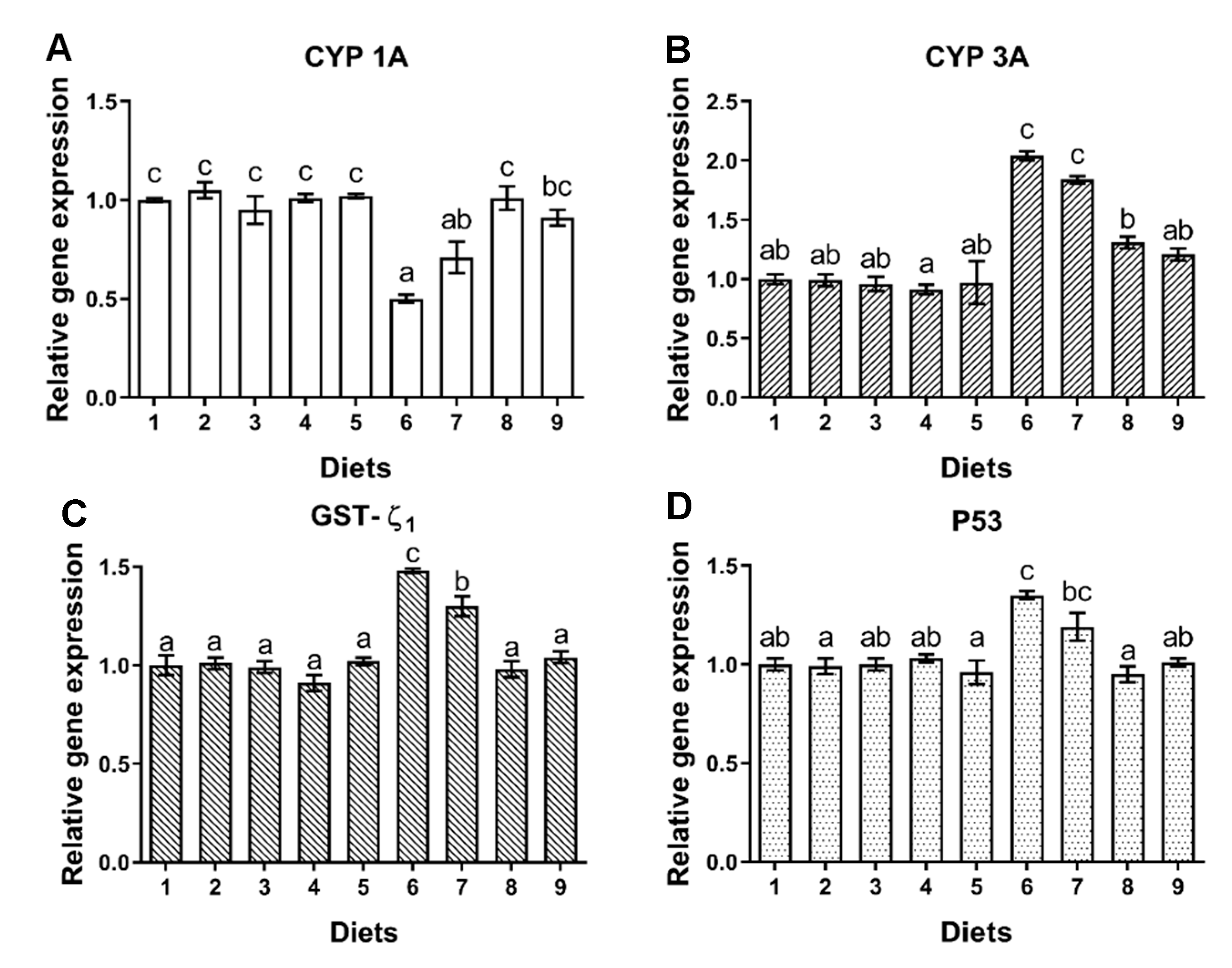
78]. GSTs is one of the important detoxifiers of AFBO [
79]. In the process of hepatocellular function, CYP1A and CYP3A play key roles in AFB
1 activation [
80,
81,
82], but CYP1A or CYP3A could also convert AFB
1 to less toxic AFM
1 or AFQ
1, respectively [
82]. In this study, the down-regulated expression of CYP1A and up-regulated expression of CYP3A and GST-ζ
1 observed in fish fed Diet 6, might indicate that the affinity of CYP1A and CYP3A to AFB
1 is different in hepatocytes of turbot. Moreover, AFBO, one of the oxidation products of AFB
1 is easily bound to DNA, which could induce DNA damage [
83,
84]. Gene p53 could promote apoptosis when DNA damage is unrepaired [
85]. Consequently, tissue antioxidant capacity may be compromised, and the oxidative metabolism of AFB
1 may contribute further to oxidative stress [
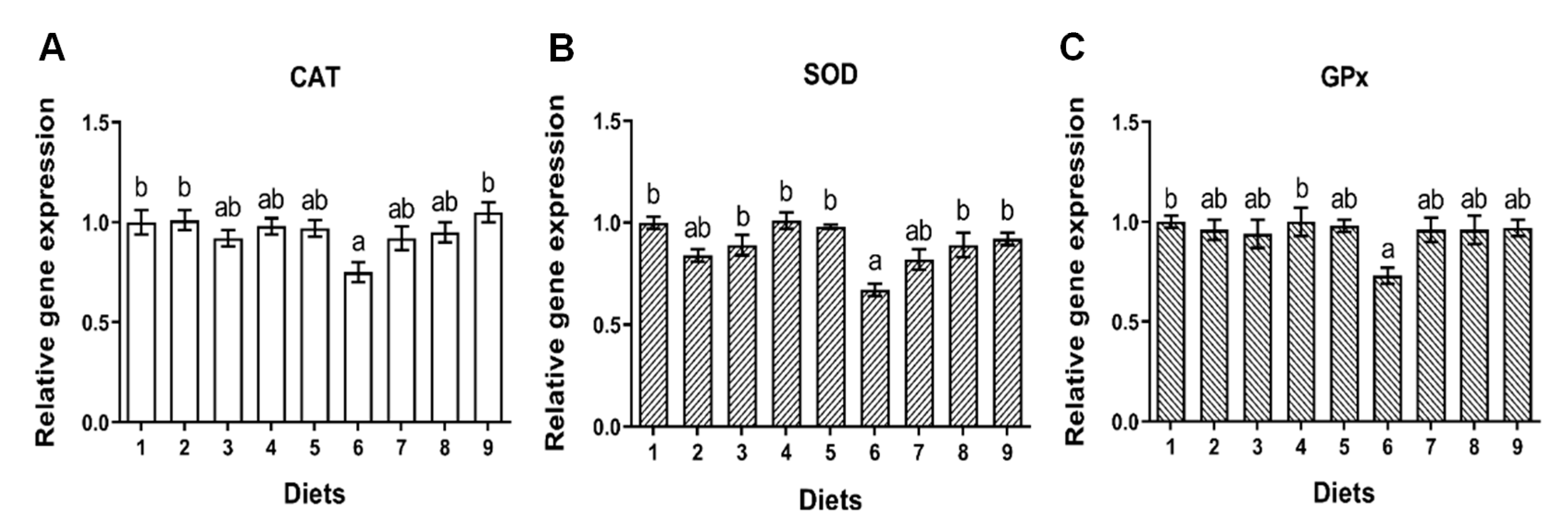
26,
86], eventually leading to oxidative damage as well. This was consistent with the decreased expression of liver p53, CAT, SOD and GPx in group of Diet 6. A previous study in rohu also reported that both DNA and oxidative damage of liver were induced by dietary AFB
1 [
87,
88].
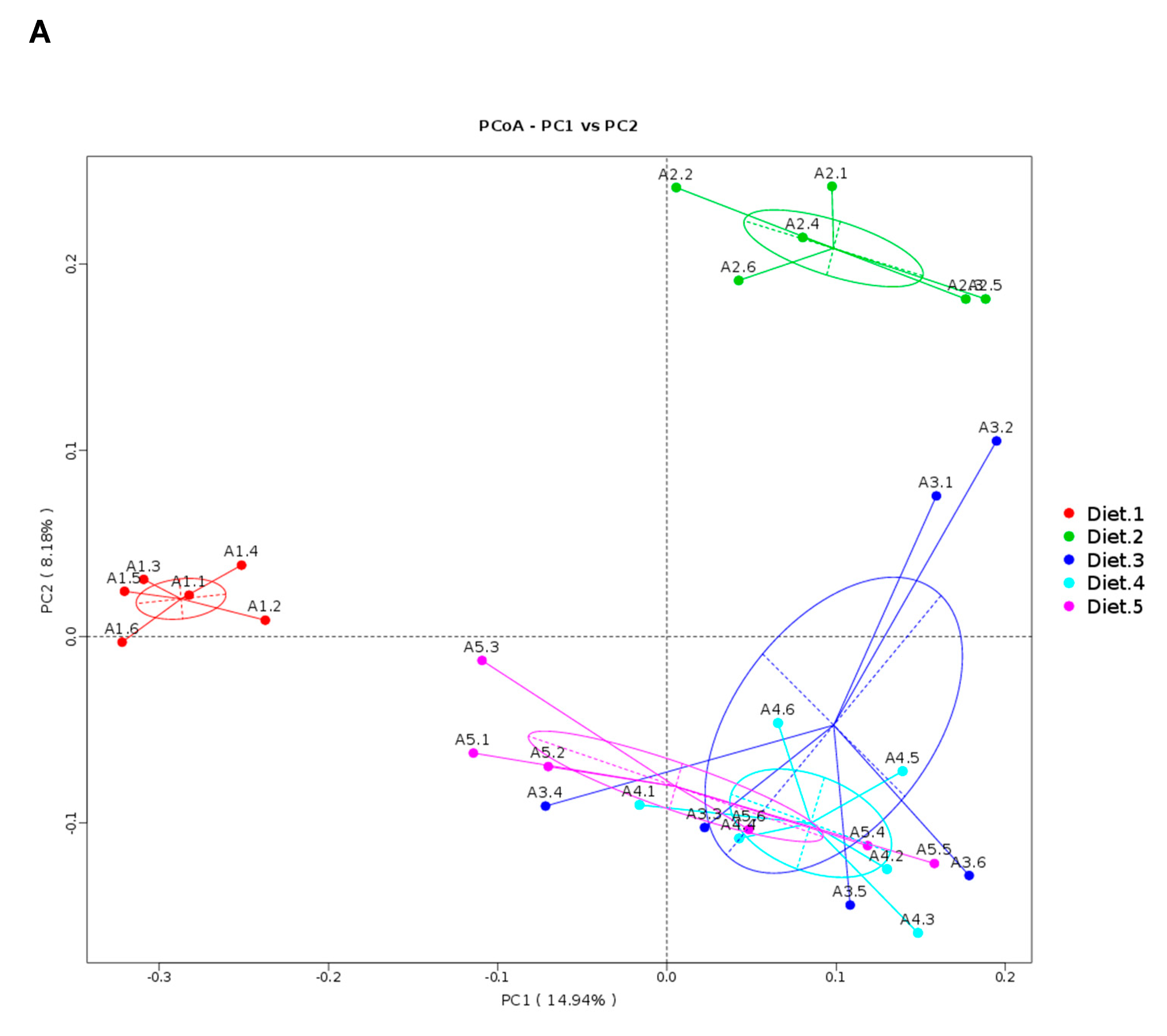
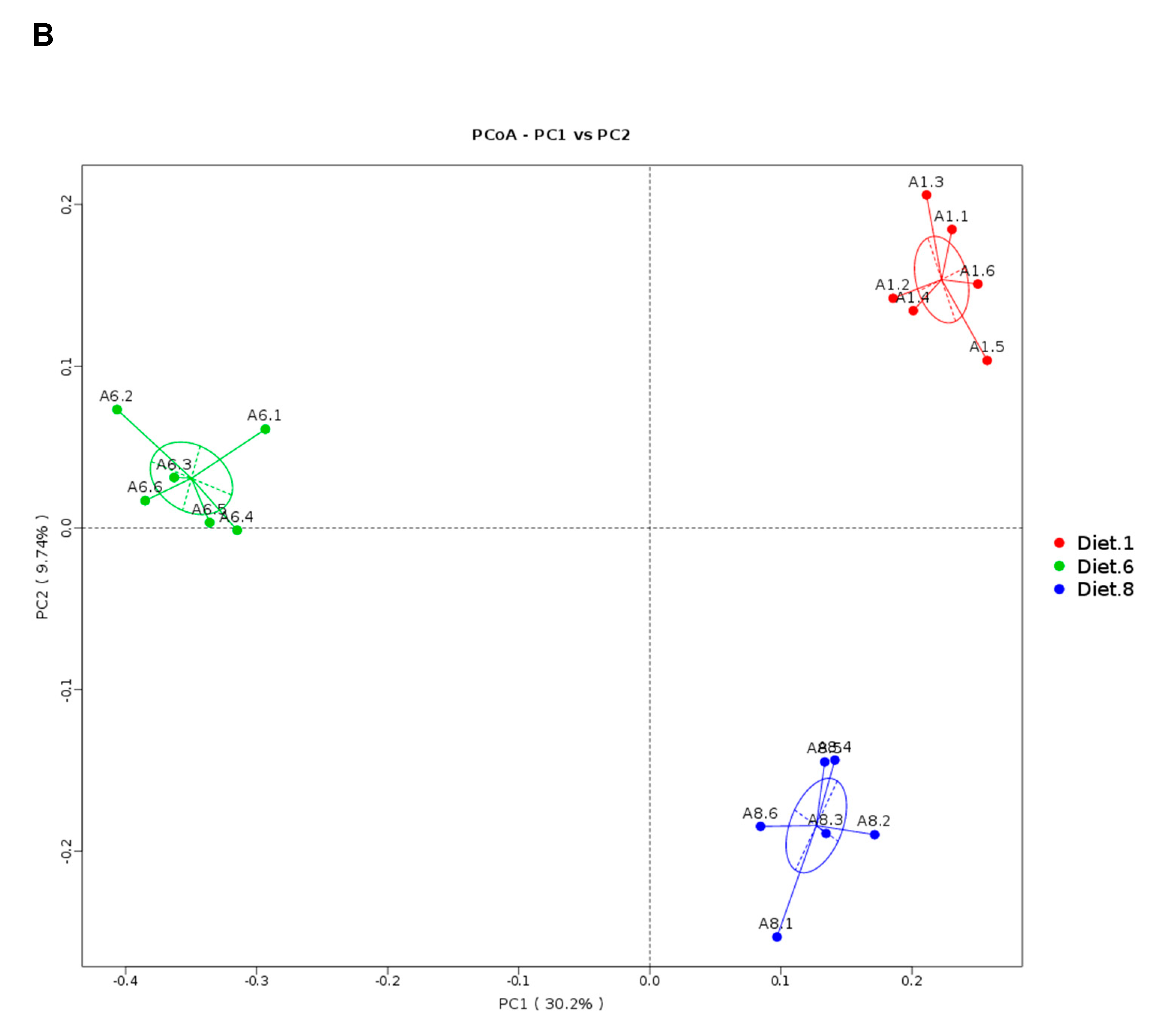
In the present study, high-throughput sequencing was used to assess the overall intestinal microbiota community of juvenile turbot in response to dietary AFB
1 and YCWE. The observation that the predominant phyla in the intestinal mucosa belonged to Firmicutes and Proteobacteria was in accordance with previous studies on turbot intestinal microbiota [
89,
90]. Compared with control group, turbot fed Diet 6 showed the lowest observed OTUs and phylogenetic diversity, and the microbiota community formed a different cluster from other groups. This was similar to a study in male Fischer 344 rat where AFB
1 significantly decreased the observed OTUs and phylogenetic diversity [
91]. In addition, Wang et al. reported the number of bacterial species at genus and phylum level were decreased by a dietary level of 5000 μg/kg AFB
1 in pacific white shrimp [
92]. The present study showed that Diet 6 significantly decreased the abundance of some potential beneficial microbiota, including
Lactobacillus,
Lactococcus,
Streptococcus,
Faecalibacterium genera, which are lactic acid producers [
93]. It has been reported that lactic acid could efficiently degrade AFB
1 into less toxic AFB
2 and AFB
2a [
94]. In addition, some studies had proved that some strains of
Lactobacillus,
Lactococcus,
Streptococcus and
Bifidobacterium could detoxify AFB
1 by cell binding mechanisms [
95,
96,
97]. Therefore, the intestinal bacteria might be involved in detoxification of AFB
1. Besides, previous studies had reported the intestinal microbiota alteration of liver diseases’ patients. For example, in the host with cirrhosis and hepatic encephalopathy diseases, the decreased abundance of non-pathogenic bacteria Lachnospiraceae and Clostridiales were observed; in the host with non-alcoholic fatty liver disease, the decreased abundance of potential probiotics
Faecalibacterium and
Bifidobacterium were observed [
98]. Therefore, the role of intestinal bacteria in liver disease induced by AFB
1 is of great significance for further research.
Several studies have reported AFB
1 residues in fish fed dietary AFB
1. While few studies paid attention to the residues of AFB
1 in serum, the current study was the first to study AFB
1 residues both in serum and muscle of aquatic animals. The result showed that AFB
1 residues in serum was higher than in muscle of turbot fed Diet 6. Furthermore, Han et al. reported AFB
1 residues in muscle and ovaries of gibel carp [
27], and similar findings had been observed in kidney and spleen of grass carp [
30] and muscle, kidney and liver of tambaqui [
36]. El-Sayed and Khalil reported AFB
1 residues in musculature of sea bass at high level (about 5 μg/kg), the consumption of which could have negative effects on human health [
16]. In contrast, two experiments with nile tilapia and red tilapia concluded that the consumption of fish muscle had no effects on human health as no AFB
1 residue was detected when fish were exposed to AFB
1 [
26,
34]. These differences might be the result of different uptake doses of AFB
1 and different AFB
1 metabolism pathways in different fishes.
In the present study, Diet 2 altered intestinal microbiota composition; however, there were no significant effects observed on growth performance, immune response and the diversity and abundance of intestinal microbiota in 67 days feeding trial. Nonetheless, low level of AFB
1 residues in serum and muscle were observed in fish fed Diet 2. The AFB
1 residues of fish muscle might accumulate more under long-term cultivation. Food contaminated with AFB
1 residues may increase the risk to human hepatoma [
99]. The safety level of AFB
1 in human food has been set at 2 μg/kg by the European Union [
100]; however, experts of FAO and WHO have given a guidance value for a provisional maximum tolerated daily intake of 1 ng AFB
1/kg body weight per day [
101]. Hence, even a low dose of AFB
1 residues poses a danger to humans.
As a complex adsorbent, previous studies on YCWE focused on livestock and poultry. In the present study, YCWE also showed favorable mitigation efficacy to the adverse effects caused by dietary AFB
1 in turbot. In addition, Diet 8 and Diet 9 lessened the immune function loss and liver damage induced by Diet 6. Apart from this, Diet 8 and Diet 9 resulted in lower AFB
1 residues in serum and muscle. These results indicate the positive adsorbing capacity of YCWE to AFB
1. Therefore, dietary YCWE could decrease the health risk resulting from feed consumption with AFB
1 contamination. The results agree with previous studies in cows (10 g YCWE/cow per day) [
58] and broilers (0.1% or 0.25% addition) [
61,
62,
102,
103]. Further, Diet 8 resulted in significantly higher alpha diversity and abundance of some potential beneficial bacteria compared to Diet 6; the abundance of potential pathogenic bacteria
Salmonella,
Aeromonas and
Comamonas was lowered by Diet 8 compared to Diet 1. This derived a suggestion that YCWE might be beneficial to regulate intestinal microbiota communities.
5. Materials and Methods
5.1. AFB1 Preparation and YCWE Preparation
Reference Standard AFB1 was purchased from Pribolab (Qingdao, China) Technology Co., Ltd. The AFB1 was dissolved in absolute ethanol (AR, Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) at 0.1 mg AFB1 per 1 mL ethanol.
Product of adsorbent (Mycosorb®) was provided by Beijing Alltech Biological Products (Beijing, China) Co., Ltd. YCWE was mixed with powder ingredients.
5.2. Experimental Diets
Based on previous studies establishing the negative effects of AFB
1 on fish, 0 µg, 20 μg (Low contamination) or 500 μg (High contamination) of AFB
1 per kg feed were included in the diets, resulting in the following nine isonitrogenous and isolipidic experimental diets: Diet 1 (control, basal diet): 0 µg AFB
1/kg diet; Diets 2–5: 20 μg/kg AFB
1 + 0%, 0.1%, 0.2%, or 0.4% YCWE; Diets 6–9: 500 μg/kg AFB
1 + 0%, 0.1%, 0.2%, or 0.4% YCWE. The basal experimental diet was formulated as shown in
Table 5. Fish meal, soybean meal and corn gluten meal were used as the main protein sources. Fish oil and soybean lecithin were used as lipid sources. Basal ingredients were purchased from Qingdao Great-seven Nutr-tech Co., Ltd (Qingdao, China). All the powder ingredients were thoroughly mixed, and then, the ethanol solution with AFB
1 was re-dissolved in water and mixed with powder ingredients. After mixing all ingredients, the feed was pelleted with an approximate diameter of 3 mm and dried until constant weight at 55 °C in a ventilated oven. The practical content of AFB
1 was detected by Beijing Alltech Biological Products (Beijing, China) Co., Ltd. The feeds were stored at −20 °C without light until the start of feeding. The chemical composition and AFB
1 content in feeds were shown in
Table 6.
5.3. Fish Husbandry and Sample Collection
Juvenile turbot (Scophthalmus maximus L.) was purchased from one commercial farm in Haiyang (Shandong, China). The feeding trial was carried out in Huanghai Aquaculture Co. Ltd. Prior to the start of the experiment, fish were acclimated to a commercial diet for two weeks with flowing water. Then the fish were fasted for 24 h and weighed (initial body weight of 12.43 ± 0.02 g). A total of 810 fish were randomly distributed to 27 cylindrical fiberglass tanks (200 L) in an indoor rearing system with flow-through seawater. In each tank 30 fish were cultured. The nine diets were randomly assigned to 27 tanks (three replications each group). Fish were fed to apparent satiation twice per day (08:00 and 18:00 h) for 67 days. The seawater of 2/3 volume was exchanged twice daily. To avoid the loss of AFB1, the experimental diets were stored in refrigerator (−20 °C), and a small portion of the feed was weighed to feed the fish every week. During the feeding trial, temperature was 12–14 °C; salinity was 30–33 ‰; pH was 7.5–8.0; dissolved oxygen was higher than 7 mg/L.
At the end of feeding trial, fish were fasted for 24 h, and then, all surviving fish were counted and weighed. After that, 2 fish of each tank were randomly selected and stored at −20 °C for whole-body analysis. Six fish of each tank were randomly selected to collect blood from caudal vein using 1 mL syringes. After clotting on ice, the serum was obtained by centrifugation with 3000 rpm for 10 min at 4 °C and stored at −80 °C for biochemical analysis. For enzyme activities and gene expressions analysis, 6 fish of each tank were randomly selected and dissected. The liver was obtained, transferred into 2 mL sterile tubes (Axygen, USA), frozen in liquid nitrogen and stored at −80 °C. For the analysis of intestinal microbiota, 2 fish of each tank were randomly selected. The exterior of fish was wiped with 70% ethanol, and the abdominal cavity was opened. After that, hind gut was obtained with sterile tools. The intestinal content was removed, the hind gut was transferred to 2 mL sterile tubes and immersed in liquid nitrogen immediately.
5.4. Growth Performance
Growth performance were calculated by using the following variables:
5.5. Feeds and Whole-Body Chemical Analysis
Chemical composition analysis of the feeds and the whole-body were performed following standard protocols of AOAC [
104]: dry matter was measured by drying samples to a constant weight at 105 °C; crude protein was determined by measuring nitrogen (N × 6.25) using Kjeldahl method; crude lipid was determined by mineral ether extraction using Soxhlet method; ash content was determined by incineration of samples at 550 °C in a muffle furnace.
The moisture was calculated with following equation:
W
1: Wet weight of matter; W
2: Dry weight of matter.
The ash content was calculated with following equation:
W
3: Ash weight; W
2: Dry weight of matter.
5.6. Biochemical Analysis of Serum
Hematological parameters were determined by using automated biochemistry analyzer (Roche/Hitachi cobas c 311 analyzer, Tokyo, Japan). GLB was calculated by subtracting ALB values from TP. The content of IgM, C3 and C4 and activity of LZM were determined by using commercial Fish ELISA kits and following manufacturer instructions (IgM: 17025, C3: 17181, C4: 17200, LZM: 17094, Quantikine® ELISA kit, R and D Systems, Minnesota, MN, USA).
5.7. RNA Extract and Real-Time PCR
The total RNA of the liver was isolated using Trizol Reagent (9108; Takara Biotech, Dalian, China). Briefly, approximate 0.2 g liver tissue was homogenized in 1 mL RNAiso Plus using a tissue grinder. Then, chemicals were added in order following the reagent instruction. The RNA concentration and quality were assessed with NanoDrop ND-1000 Spectrophotometer (Thermo Scientific, Waltham, MA, USA). The integrity of extracted RNA was determined by electrophoresis on a 1.2% (w/v) agarose gel. After that, 1000 ng RNA was reverse transcribed to cDNA in 20 μL reactions using PrimeScript RT reagent Kit with gDNA Eraser (RR047A; Takara Biotech, Dalian, China). Then, real-time PCR was performed in a total 25 μL volume: 1 μL cDNA template (≤ 50 ng); 1 μL Forward primer (10 μM); 1 μL Reverse primer (10 μM); 9.5 μL DEPC-treated water (Sangon biotech, Shanghai, China); 12.5 μL TB Green™ Premix EX Taq II™ (RR820 A, Takara Biotech, Dalian, China). A two-step real-time PCR amplification program was used: 95 °C for 2 min and then 40 cycles of 95 °C for 10 s and 60 °C for 30 s. At last, melting curve analysis was used to ensure the specification of PCR product for each primer pair.
Specific primers for target genes and housekeeping genes, designed in NCBI, were synthesized by Sangon Biotech (Shanghai) Co., Ltd., and then the application efficiency was assessed (
Table S3). All the real-time PCR analysis were performed using a quantitative thermal cycler (CFX96 Touch™ Real-Time PCR Detection System, Bio-Rad, Richmond, CA, USA). The genes expressions levels were normalized using relative quantitative method (2
−ΔΔCT) referencing gene β-actin of turbot [
105].
5.8. DNA Extract of Intestinal Microbiota and Sequencing Analysis
Genomic DNA sample was extracted from the intestinal mucosa layer using the QIAamp Fast DNA Stool Mini Kit (51604, Qiagen, Hilden, Germany) under sterile conditions (alcohol flame) following the manufacturer manual with some modifications [
89,
90]. PCR amplification of V4 region of 16S rRNA (515F/806R primer), quality and purity of PCR product were assessed by Beijing Novogene Genomics Technology Co. Ltd. (Beijing, China). Sequencing was conducted on an Illumina NovaSeq platform provided by Beijing Novogene Genomics Technology Co. Ltd. (Beijing, China).
For the sequence data analysis, Fast Length Adjustment of SHort reads (FLASH) was used to merge paired-end reads from the original DNA fragments when there were overlaps between reads1 and reads2 [
106]. Sequencing reads were assigned to each sample with unique barcodes. Cutadapter was used to remove the adapter sequence, barcode sequence, primer sequence and to filter low-quality reads of raw reads [
107]. The UCHIME algorithm was used to detect and remove chimeric sequences and obtain effective reads that would be used for further analysis [
108]. After dereplication, abundance sort and discarding singletons reads, sequences with ≥97% similarity were clustered to the same OUTs (operational taxonomic units) using UPARSE [
109]. Representative sequence for each OTU was screened for further annotation using Silva Database(v132) based on Ribosomal Database Project (RDP) classifier [
110]. Alpha diversity indices (OTUs, Chao1, ACE, Shannon, Simpson and PD whole tree) were calculated with Quantitative Insights Into Microbial Ecology (QIIME) and displayed with R software (v 3.6.2) [
111]. Beta diversity on unweighted UniFrac for Principal Coordinate Analysis (PCoA) and Unweighted Pair Group Method with Arithmetic mean (UPGMA) Clustering was calculated with QIIME and displayed with R software as well.
5.9. AFB1 Residues of Serum and Muscle
The AFB
1 residues of serum and dorsal muscle of turbot were detected following the methods provided by Wang et al. [
112]. This analysis was completed at the Institute of Quality Standard and Testing Technology for Agro-Products of CAAS (Beijing, China).
5.10. Statistical Analysis
Statistical software SPSS 22.0 for Windows (IBM SPSS corporation, Chicago, IL, USA) was used for the data analysis. Results were analyzed by one-way analysis of variance (ANOVA). Tukey’s multiple-range test was used for the multiple comparisons of group means. Differences were regarded as significant when p < 0.05. T-test was used for the comparisons of AFB1 residues in fish tissues of any two groups; “*” was marked when p < 0.05, and “**” was marked when p < 0.01.
Multi Response Permutation Procedure (MRPP) and Adonis test were employed to assess the difference of microbiota composition within or between groups using the vegan package in R software (v 3.6.2). MetaStat analysis [
113] was conducted to identify the differential abundant taxa between groups.
5.11. Ethics Statement
The animal study was reviewed and approved by the Animal Care Committee of Ocean University of China (No. OUCFC2018001325, 24 August 2018).

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}