New Data on Cylindrospermopsin Toxicity

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
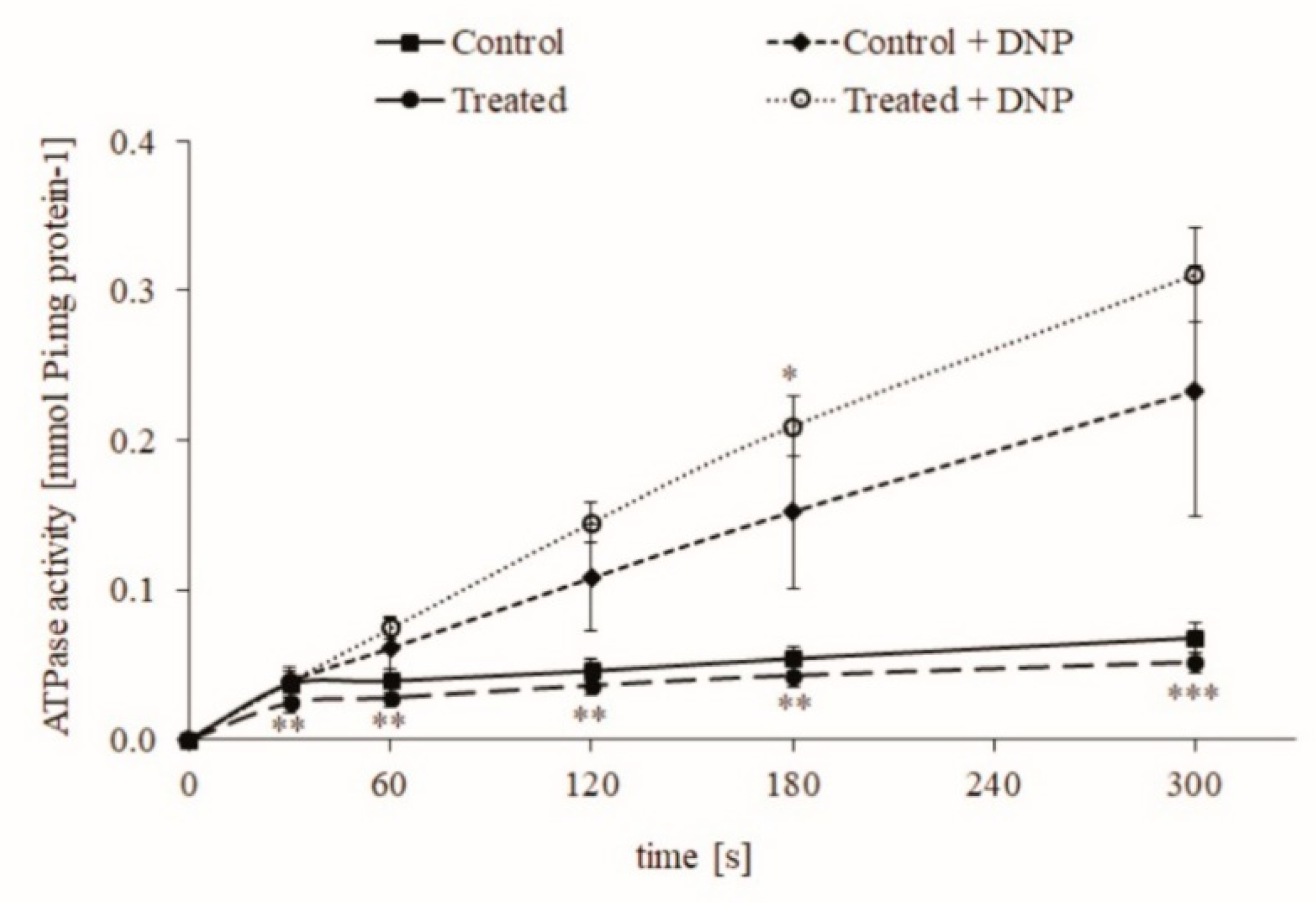
2.1. The Effect of Chronic Intake of Contaminated Water on Rat Liver Mitochondria
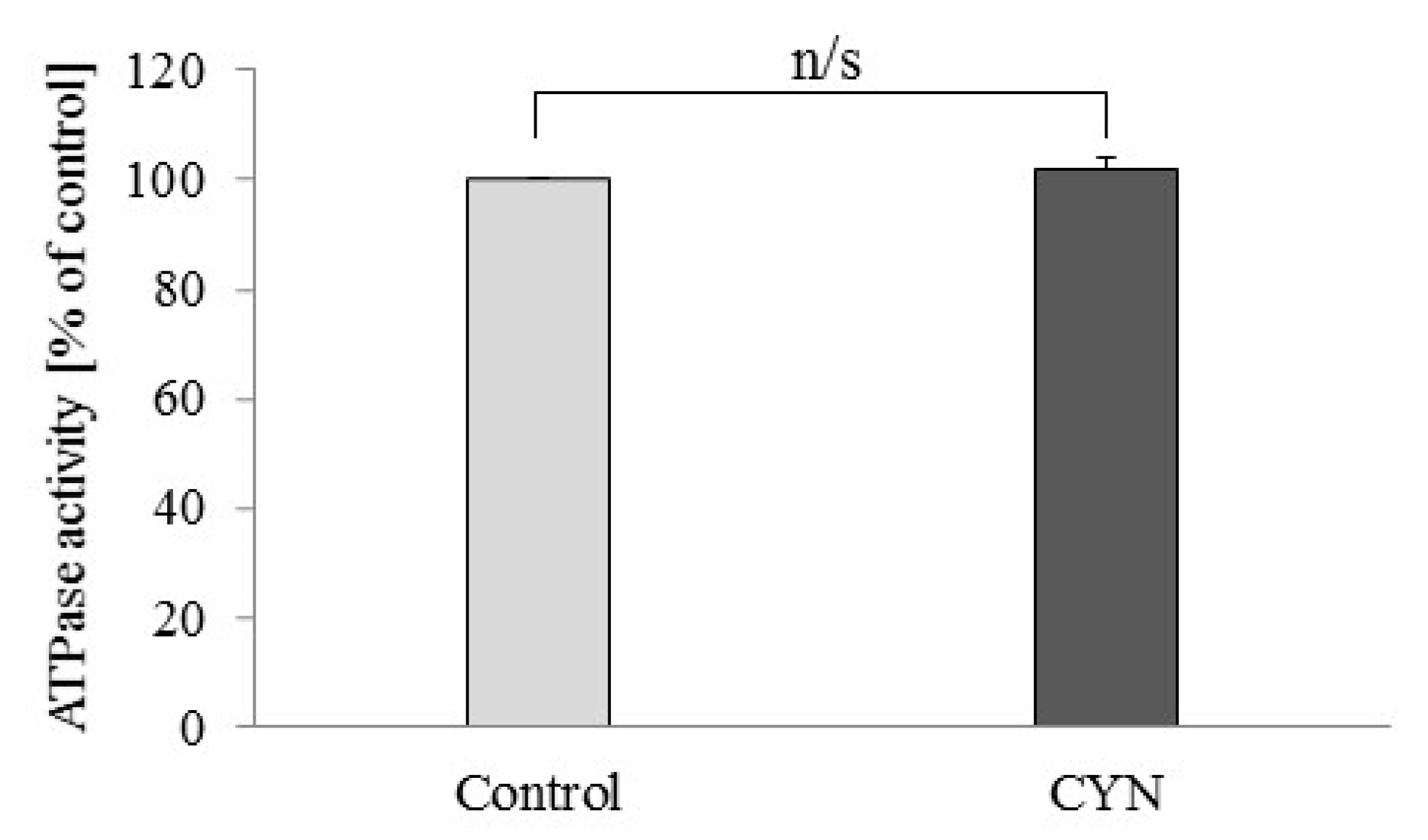
2.2. Effect of CYN on Freeze-Thawed Mitochondria
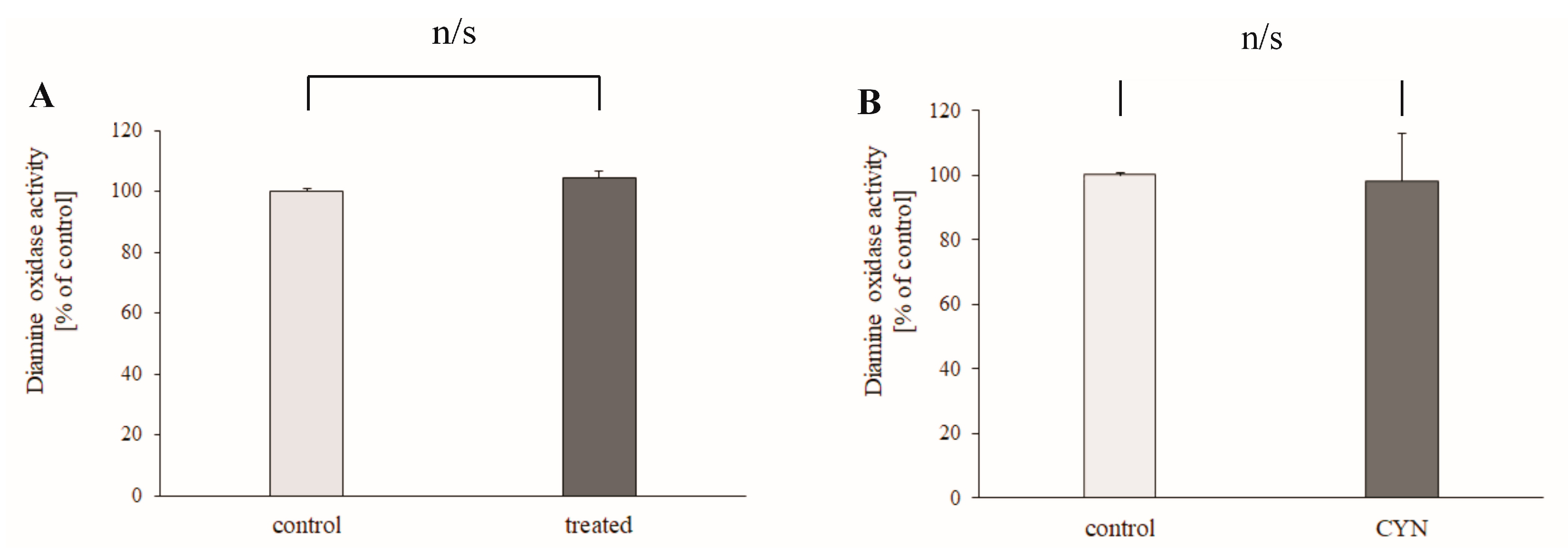
2.3. DAO Activity
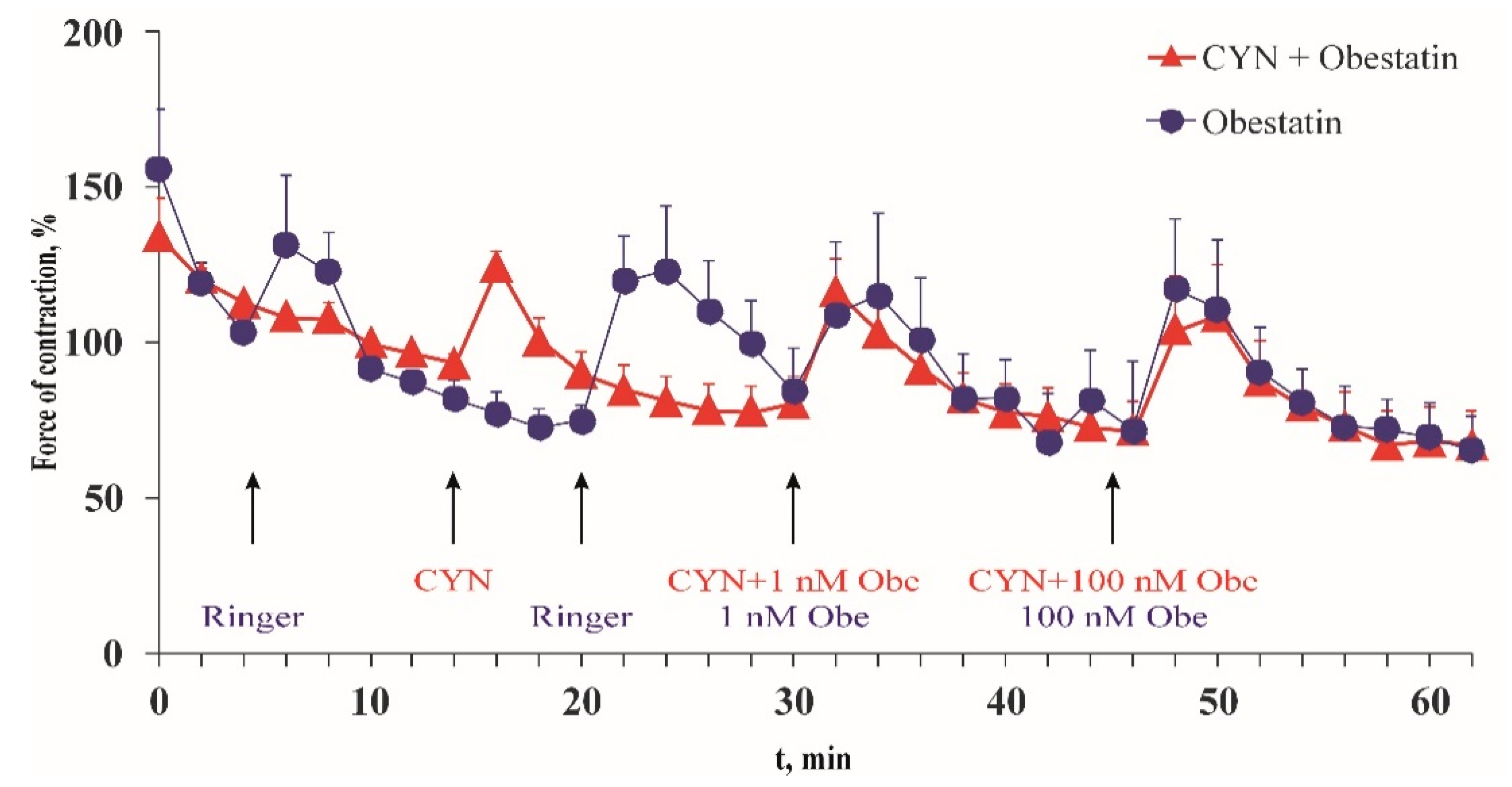
2.4. Effect of CYN on Frog Heart Preparations In Vitro
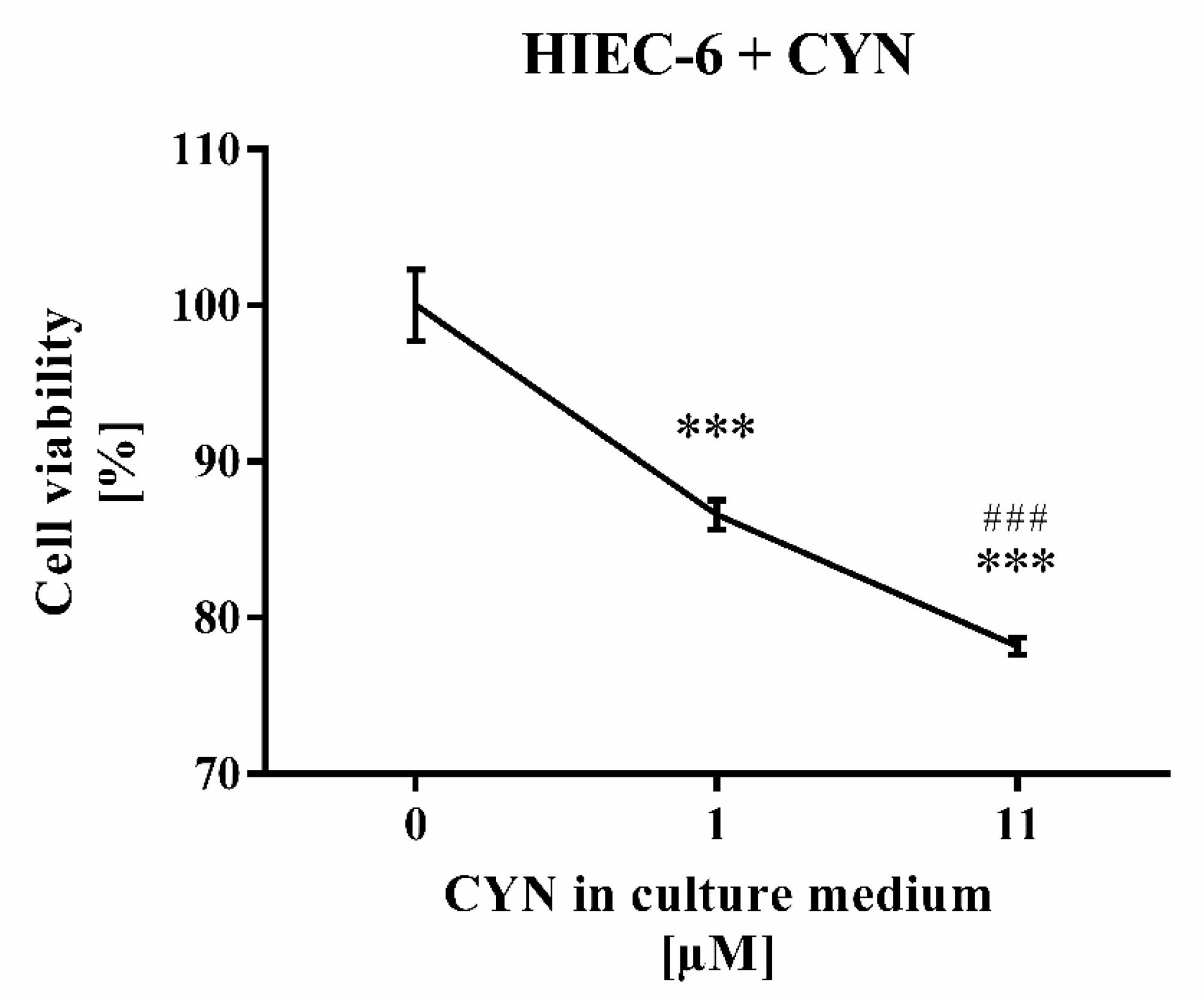
2.5. CYN Cytotoxicity Tested on HIEC-6
3. Discussion
4. Materials and Methods
4.1. Isolation of Intact Rat Liver Mitochondria
4.2. Assay of Mitochondrial ATPase Activity
4.3. ATPase Activity of Intact Rat Liver Mitochondria
4.4. ATPase Activity of Freeze-Thawed Mitochondria
4.5. Assay of Rat Liver DAO Activity
4.6. Study of Excised Frog Heart Contraction
4.7. Cytotoxicity Test
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funar, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the Chemistry, Toxicology and Genetics of the Cyanobacterial Toxins, Microcystin, Nodularin, Saxitoxin and Cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, M.; Ploux, O.; Metcalf, J.S.; Mejean, A.; Pawlik-Skowronska, B.; Furey, A. Anatoxin-a, Homoanatoxin-a, and Natural Analogues. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons: Chichester, UK, 2017; pp. 138–147. [Google Scholar]
- Guzmán-Guillén, R.; Prieto, A.I.; Moyano, R.; Blanco, A.; Vasconcelos, V.; Cameán, A.M. Dietary l-carnitine prevents histopathological changes in tilapia (Oreochromis niloticus) exposed to cylindrospermopsin. Environ. Toxicol. 2017, 32, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Kokociński, M.; Cameán, A.M.; Carmeli, S.; Guzmán-Guillén, R.; Jos, A.; Mankiewicz-Boczek, J.; Metcalf, J.S.; Moreno, I.M.; Prieto, A.I.; Sukenik, A. Cylindrospermopsin and Congeners. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 127–137. [Google Scholar]
- Catherine, A.; Bernard, C.; Spoof, L.; Bruno, M. Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 109–126. [Google Scholar]
- Ballot, A.; Bernar, C.; Fastner, J. Saxitoxin and Analogues. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 148–154. [Google Scholar]
- Jos, A.; Cameán, A.M. Freshwater Algal Toxins: Monitoring and Toxicity Profile. Toxins 2020, 12, 653. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Xie, P.; Liu, Y.; Li, G.; Xiong, Q.; Hao, L.; Li, H. The profound effects of microcystin on cardiac antioxidant enzymes, mitochondrial function and cardiac toxicity in rat. Toxicology 2009, 257, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Díez-Quijada, L.; Medrano-Padial, C.; Llana-Ruiz-Cabello, M.; Cătunescu, G.M.; Moyano, R.; Risalde, M.A.; Cameán, A.M.; Jos, A. Cylindrospermopsin-Microcystin-LR Combinations May Induce Genotoxic and Histopathological Damage in Rats. Toxins 2020, 12, 348. [Google Scholar] [CrossRef]
- Puerto, M.; Jos, Á.; Pichardo, S.; Moyano, R.; Blanco, A.; Cameán, A.M. Acute exposure to pure Cylindrospermopsin results in oxidative stress and pathological alterations in Tilapia (Oreochromis niloticus). Environ. Toxicol. 2014, 29, 371–385. [Google Scholar] [CrossRef]
- Pichardo, S.; Cameán, A.M.; Jos, Á. In vitro toxicological assessment of Cylindrospermopsin: A review. Toxins 2017, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Froscio, S.M.; Humpage, A.R.; Burcham, P.C.; Falconer, I.R. Cylindrospermopsin-induced protein synthesis inhibition and its dissociation from acute toxicity in mouse hepatocytes. Environ. Toxicol. Int. J. 2003, 18, 243–251. [Google Scholar] [CrossRef]
- Chernoff, N.; Rogers, E.H.; Zehr, R.D.; Gage, M.I.; Malarkey, D.E.; Bradfield, C.A.; Liu, Y.; Schmid, J.E.; Jaskot, R.H.; Richards, J.H.; et al. Toxicity and recovery in the pregnant mouse after gestational exposure to the cyanobacterial toxin, cylindrospermopsin. J. Appl. Toxicol. 2011, 31, 242–254. [Google Scholar] [CrossRef] [Green Version]
- Runnegar, M.T.; Kong, S.-M.; Zhong, Y.-Z.; Lu, S.C. Inhibition of reduced glutathione synthesis by cyanobacterial alkaloid cylindrospermopsin in cultured rat hepatocytes. Biochem. Pharmacol. 1995, 49, 219–225. [Google Scholar] [CrossRef]
- Gutiérrez-Praena, D.; Pichardo, S.; Jos, A.; Moreno, F.; Cameán, A.M. Biochemical and pathological toxic effects induced by the cyanotoxin cylindrospermopsin on the human cell line Caco-2. Water Res. 2012, 46, 1566–1575. [Google Scholar] [CrossRef]
- Liebel, S.; Oliveira Ribeiro, C.A.; Magalhaes, V.F.; Silva, C.R.; Rossi, S.C.; Randi, M.A.F.; Neto, F.F. Low concentrations of cylindrospermopsin induce increases of reactive oxygen species levels, metabolism and proliferation in human hepatoma cells (HepG2). Toxicol. In Vitro 2015, 29, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Scarlett, K.R.; Kima, S.; Lovin, L.M.; Chatterjee, S.; Scott, J.T.; Brooks, B.W. Global scanning of cylindrospermopsin: Critical review and analysis of aquatic occurrence, bioaccumulation, toxicity and health hazards. Sci. Total Environ. 2020, 139807. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.P.; Jaja-Chimedza, A.; Davalos-Lind, L.; Lind, O. Apparent bioaccumulation of cylindrospermopsin and paralytic shellfish toxins by finfish in Lake Catemaco (Veracruz, Mexico). Food Addit. Contam. Part A Chem. 2012, 29, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Thomas, T.J. Polyamines in cell growth and cell death: Molecular mechanisms and therapeutic applications. Cell Mol. Life Sci. 2001, 58, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A. Gastrointestinal toxicity biomarkers. In Biomarkers in Toxicology; Gupta, R.C., Ed.; Elsevier: San Diego, CA, USA, 2014; pp. 269–277. [Google Scholar]
- Boyce, S.; Tipton, K.F.; O’Sullivan, M.I.; Davey, G.P.; Gildea, M.M.; McDonald, A.G.; Olivieri, A.; O’Sullivan, J. Nomenclature and potential functions of copper amine oxidase. In Copper Amine Oxidases Structures. Catalytic Mechanisms, and Role in Pathophysiology; Floris, G., Mondovì, B., Eds.; CRC Press: Boca Raton, FL, USA, 2009; pp. 5–13. [Google Scholar]
- Hinojosa, M.G.; Gutierrez-Praena, D.; Prieto, A.I.; Guzman-Guillen, R.; Jos, A.; Camean, A.M. Neurotoxicity induced by microcystins and cylindrospermopsin: A review. Sci. Total Environ. 2019, 668, 547–565. [Google Scholar] [CrossRef]
- Kiss, T.; Vehovszky, Á.; Hiripi, L.; Kovács, A.; Vörös, L. Membrane effects of toxins isolated from a cyanobacterium, Cylindrospermopsis raciborskii, on identified molluscan neurones. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 131, 167–176. [Google Scholar] [CrossRef]
- Kubickova, B.; Babica, P.; Hilscherová, K.; Sindlerova, L. Effects of cyanobacterial toxins on the human gastrointestinal tract and the mucosal innate immune system. Environ. Sci. Eur. 2019, 31, 31. [Google Scholar] [CrossRef] [Green Version]
- Seawright, A.A.; Nolan, C.C.; Shaw, G.R.; Chiswell, R.K.; Norris, R.L.; Moore, M.R.; Smith, M.J. The oral toxicity for mice of the tropical cyanobacterium Cylindrospermopsis raciborskii (Woloszynska). Environ. Toxicol. 1999, 14, 135–142. [Google Scholar] [CrossRef]
- Pichardo, S.; Devesa, V.; Puerto, M.; Cameán, A.M. Intestinal transport of cylindrospermopsin using the Caco-2 cell line. Toxicol. In Vitro 2017, 38, 142–149. [Google Scholar] [CrossRef]
- Fernandez, D.A.; Louzao, M.C.; Vilarico, N.; Fraga, M.; Espiña, B.; Vieytes, M.C.; Botana, L.M. Evaluation of the intestinal permeability and cytotoxic effects of cylindrospermopsin. Toxicon 2014, 91, 23–34. [Google Scholar] [CrossRef]
- Stefanova, K.; Radkova, M.; Uzunov, B.; Gärtner, G.; Stoyneva-Gärtner, M. Pilot search for cylindrospermopsin-producers in nine shallow Bulgarian waterbodies reveals nontoxic strains of Raphidiopsis raciborskii, R. mediterranea and Chrysosporum bergii. Biotech. Biotech. Equip. 2020, 34, 384–394. [Google Scholar] [CrossRef]
- Stoyneva-Gärtner, M.P.; Uzunov, B.A.; Descy, J.-P.; Gärtner, G.; Draganova, P.H.; Borisova, C.I.; Pavlova, V.; Mitreva, M. Pilot application of drone-observations and pigment marker detection by HPLC in the studies of CyanoHABs in Bulgarian inland waters. Mar. Freshw. Res. 2019, 71, 606–616. [Google Scholar] [CrossRef]
- Durašević, S.; Dorđević, J.; Drenca, T.; Jasnić, N.; Cvijić, G. Influence of vitamin C supplementation on the oxidative status of rat liver. Arch. Biol. Sci. 2008, 60, 169–173. [Google Scholar] [CrossRef]
- Wiederkehr, A.; Wollheim, C.B. Mitochondrial signals drive insulin secretion in the pancreatic b-cell. Mol. Cell. Endocrinol. 2012, 353, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Terao, K.; Ohmori, S.; Igarashi, K.; Ohtani, I.; Watanabe, M.F.; Harada, K.I.; Ito, E.; Watanabe, M. Electron microscopic studies on experimental poisoning in mice induced by cylindrospermopsin isolated from blue-green alga Umezakia natans. Toxicon 1994, 32, 32–843. [Google Scholar] [CrossRef]
- Sazdova, I.; Ilieva, B.; Minkov, I.; Schubert, R.; Gagov, H. Obestatin as contractile mediator of excised frog heart. Cent. Eur. J. Biol. 2009, 4, 327–334. [Google Scholar] [CrossRef]
- Froscio, S.M.; Fanok, S.; Humpage, A.R. Cytotoxicity screening for the cyanobacterial toxin cylindrospermopsin. J. Toxicol. Environ. Health Part A 2009, 72, 345–349. [Google Scholar] [CrossRef]
- Humpage, A.R.; Fontaine, F.; Froscio, S.; Burcham, P.; Falconer, I.R. Cylindrospermopsin genotoxicity and cytotoxicity: Role of cytochrome P-450 and oxidative stress. J. Toxicol. Environ. Health A 2005, 68, 739–753. [Google Scholar] [CrossRef]
- Johnson, D.R.; Lardy, H.A. Isolation of liver or kidney mitochondria. In Methods of Enzymology; Academic Press: New York, NY, USA, 1967; Volume 10, pp. 94–95. [Google Scholar]
- Chichova, M.; Shkodrova, M.; Vasileva, P.; Kirilova, K.; Doncheva-Stoimenova, D. Influence of silver nanoparticles on the activity of rat liver mitochondrial ATPase. J. Nanopart. Res. 2014, 16, 2243. [Google Scholar] [CrossRef]
- Dimitrov, O.; Pavlov, V.; Jotova, I. Effects of female sex hormones on polyamine-oxidizing enzyme activities and polyamine concentrations in immature rat uterus and liver. J. Exp. 1996, 52, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Straub, W. Mechanismus der Muskarinwirkung am Herzen und des Antagonismus Atropin-Muskarin. Zentralblatt für Physiologie 1905, 19, 302–304. [Google Scholar]
- Mosmann, T. Rapid Colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assay. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time after Application | Time Control Ringer, ±SEM | CYN, ±SEM | p |
|---|---|---|---|
| 2 min | 93% ± 14.28% (n = 7) | 125% ± 4.74% (n = 6) | 0.091 |
| 4 min | 90% ± 15.86% (n = 7) | 101% ± 7.09% (n = 6) | 0.552 |
| 6 min | 75% ± 14.98% (n = 7) | 90% ± 7.03% (n = 6) | 0.388 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chichova, M.; Tasinov, O.; Shkodrova, M.; Mishonova, M.; Sazdova, I.; Ilieva, B.; Doncheva-Stoimenova, D.; Kiselova-Kaneva, Y.; Raikova, N.; Uzunov, B.; et al. New Data on Cylindrospermopsin Toxicity. Toxins 2021, 13, 41. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010041
Chichova M, Tasinov O, Shkodrova M, Mishonova M, Sazdova I, Ilieva B, Doncheva-Stoimenova D, Kiselova-Kaneva Y, Raikova N, Uzunov B, et al. New Data on Cylindrospermopsin Toxicity. Toxins. 2021; 13(1):41. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010041
Chicago/Turabian StyleChichova, Mariela, Oskan Tasinov, Milena Shkodrova, Milena Mishonova, Iliyana Sazdova, Bilyana Ilieva, Dilyana Doncheva-Stoimenova, Yoana Kiselova-Kaneva, Neli Raikova, Blagoy Uzunov, and et al. 2021. "New Data on Cylindrospermopsin Toxicity" Toxins 13, no. 1: 41. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010041