In Vitro Neurotoxicity of Chinese Krait (Bungarus multicinctus) Venom and Neutralization by Antivenoms

 and
and
Abstract
:1. Introduction
2. Results
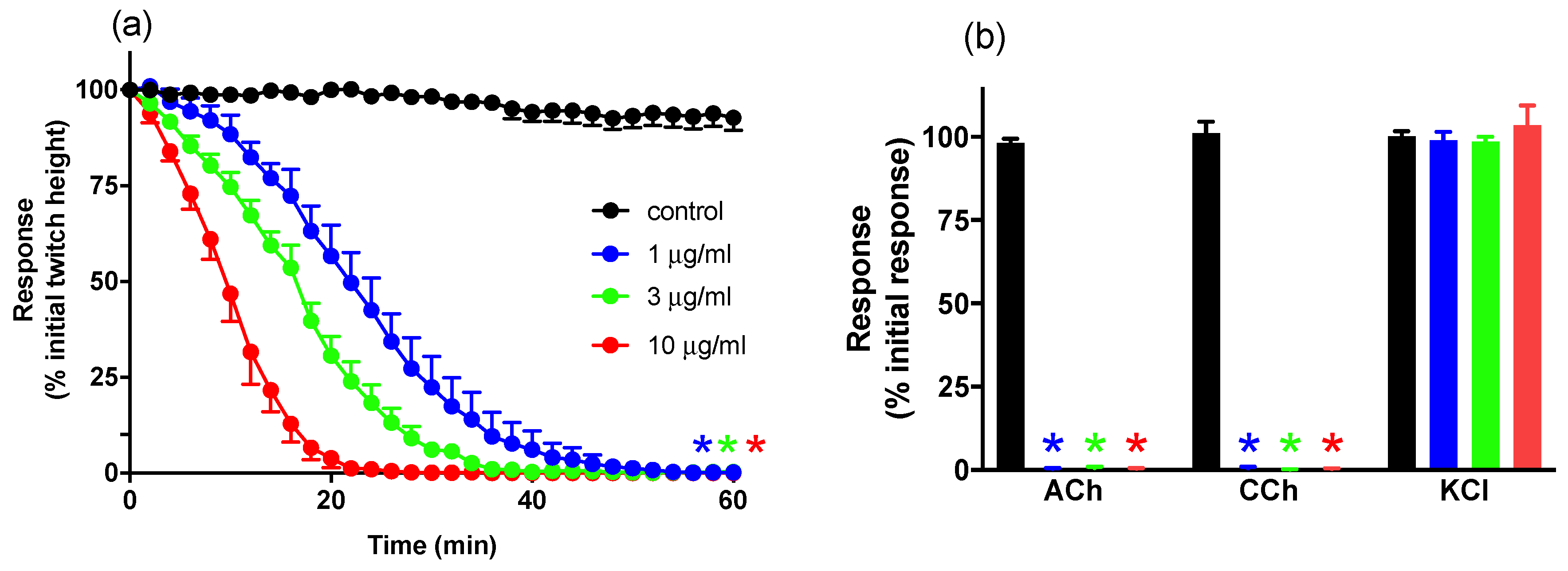
2.1. In Vitro Neurotoxicity
2.1.1. Concentration-Dependent Inhibition of Twitches and Exogenous Agonists Responses
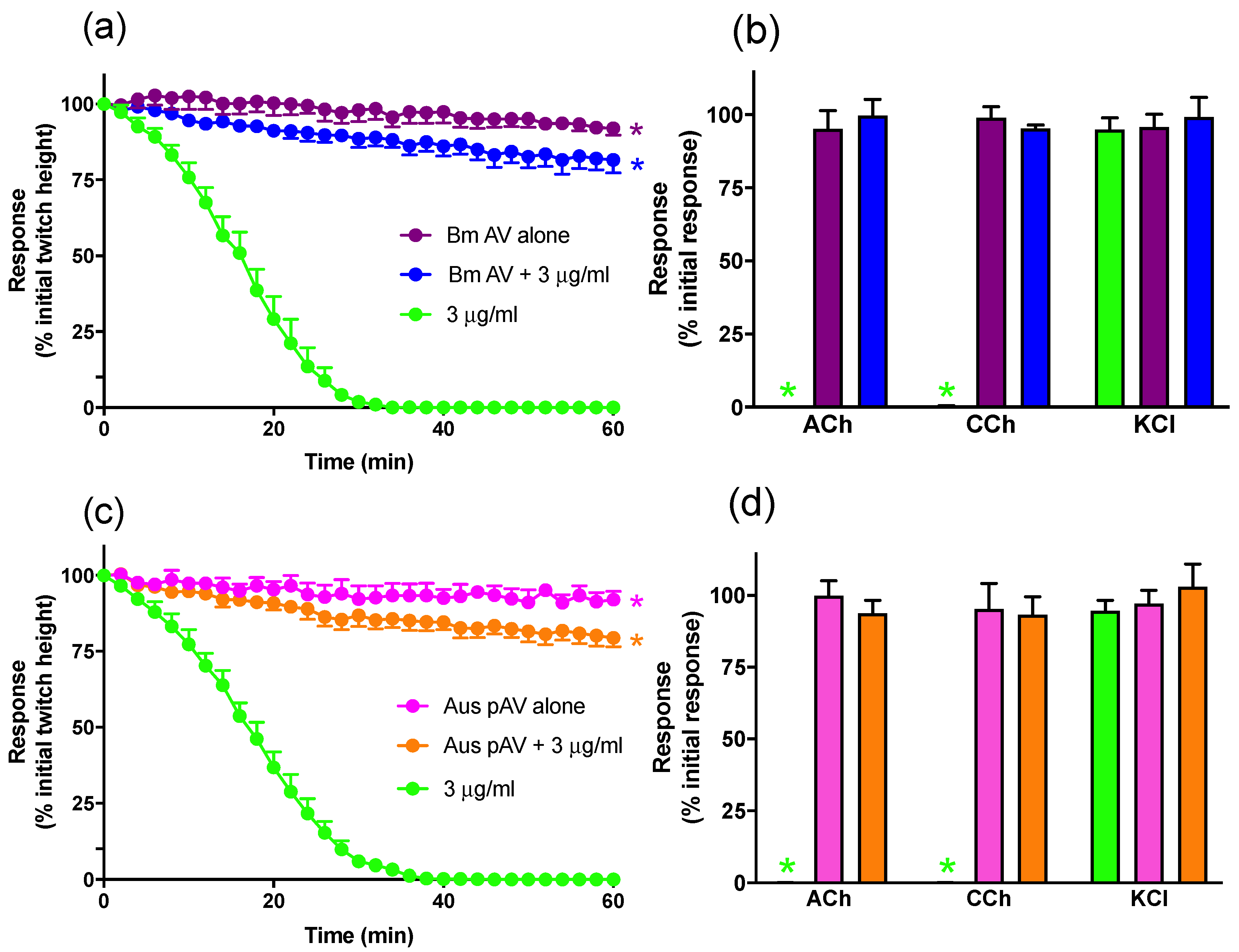
2.1.2. Prevention of In Vitro Neurotoxicity with Antivenom
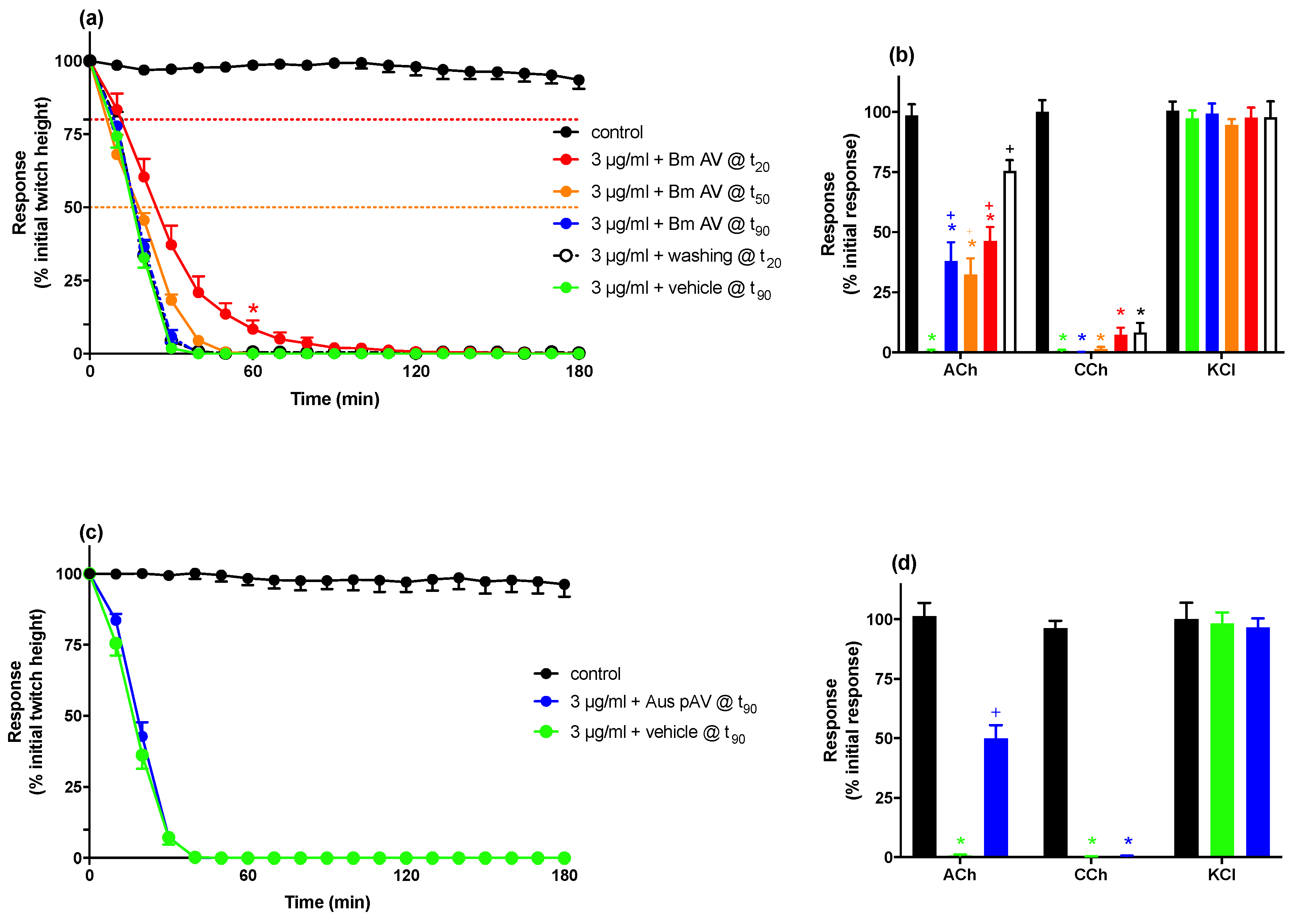
2.1.3. Reversal of In Vitro Neurotoxicity with Antivenom and Washings
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Venom and Antivenoms
5.2. Chemicals and Reagents
5.3. Chick Biventer Cervicis Nerve-Muscle Preparation
5.4. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Zhao, E.M. Snakes in China, 1st ed.; Anhui Science and Technology Publishing House: Hefei, China, 2006; pp. 290–293. [Google Scholar]
- Lan, H.; Chen, Y.C. Chinese Venomous Snake and Treatment of Snakebite, 1st ed.; Shanghai Scientific & Technical Publishers: Shanghai, China, 2008; pp. 535–540. [Google Scholar]
- Uetz, P.; Freed, P.; Hošek, J. (Eds.) 2020. Available online: http://www.reptile-database.org (accessed on 20 August 2020).
- Chan, J.C.; Cockram, C.S.; Buckley, T.; Young, K.; Kay, R.; Tomlinson, B. Evenoming by Bungarus multicinctus (many-banded krait) in Hong Kong. J. Trop. Med. Hyg. 1995, 98, 457–460. [Google Scholar] [PubMed]
- Mao, Y.C.; Liu, P.Y.; Chiang, L.C.; Liao, S.C.; Su, H.Y.; Hsieh, S.Y.; Yang, C.C. Bungarus multicinctus multicinctus snakebite in Taiwan. Am. J. Trop. Med. Hyg. 2017, 96, 1497–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- 2018 Expert Consensus on China Snake-bites Rescue and Treatment. Chin. J. Emerg. Med. 2018, 27, 1315–1322. [CrossRef]
- Nirthanan, S.; Gwee, M.C. Three-finger alpha-neurotoxins and the nicotinic acetylcholine receptor, forty years on. J. Pharmacol. Sci. 2004, 94, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowan, E.G. What does beta-bungarotoxin do at the neuromuscular junction? Toxicon 2001, 39, 107–118. [Google Scholar] [CrossRef]
- Shan, L.L.; Gao, J.F.; Zhang, Y.X.; Shen, S.S.; He, Y.; Wang, J.; Ma, X.M.; Ji, X. Proteomic characterization and comparison of venoms from two elapid snakes (Bungarus multicinctus and Naja atra) from China. J. Proteom. 2016, 138, 83–94. [Google Scholar] [CrossRef]
- Chang, C.C.; Lee, C.Y. Isolation of neurotoxins from the venom of Bungarus multicinctus and their modes of neuromuscular blocking action. Arch. Int. Pharm. Ther. 1963, 144, 241–257. [Google Scholar]
- Dixon, R.W.; Harris, J.B. Nerve terminal damage by beta-bungarotoxin: Its clinical significance. Am. J. Pathol. 1999, 154, 447–455. [Google Scholar] [CrossRef]
- Prasarnpun, S.; Walsh, J.; Awad, S.S.; Harris, J.B. Envenoming bites by kraits: The biological basis of treatment-resistant neuromuscular paralysis. Brain 2005, 128, 2987–2996. [Google Scholar] [CrossRef] [Green Version]
- Prasarnpun, S.; Walsh, J.; Harris, J.B. Beta-bungarotoxin-induced depletion of synaptic vesicles at the mammalian neuromuscular junction. Neuropharmacology 2004, 47, 304–314. [Google Scholar] [CrossRef]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon 2013, 66, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Rusmili, M.R.; Yee, T.T.; Mustafa, M.R.; Othman, I.; Hodgson, W.C. In-vitro neurotoxicity of two Malaysian krait species (Bungarus candidus and Bungarus fasciatus) venoms: Neutralization by monovalent and polyvalent antivenoms from Thailand. Toxins 2014, 6, 1036–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Q.; Huynh, T.M.; Konstantakopoulos, N.; Isbister, G.K.; Hodgson, W.C. An examination of the neutralization of in vitro toxicity of Chinese cobra (Naja atra) venom by different antivenoms. Biomedicines 2020, 8, 377. [Google Scholar] [CrossRef]
- Tibballs, J.; Kuruppu, S.; Hodgson, W.C.; Carroll, T.; Hawdon, G.; Sourial, M.; Baker, T.; Winkel, K. Cardiovascular, haematological and neurological effects of the venom of the Papuan New Guinean small-eyed snake (Micropechis ikaheka) and their neutralisation with CSL polyvalent and black snake antivenoms. Toxicon 2003, 42, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Hodgson, W.C.; Isbister, G.K. Cross-neutralisation of in vitro neurotoxity of Asian and Australian snake neurotoxins and venoms by different antivenoms. Toxins 2016, 8, 302. [Google Scholar] [CrossRef] [Green Version]
- Leong, P.K.; Tan, N.H.; Fung, S.Y.; Sim, S.M. Cross neutralisation of Southeast Asian cobra and krait venoms by Indian polyvalent antivenoms. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 731–737. [Google Scholar] [CrossRef]
- Leong, P.K.; Sim, S.M.; Fung, S.Y.; Sumana, K.; Sitprija, V.; Tan, N.H. Cross neutralization of Afro-Asian cobra and Asian krait venoms by a Thai polyvalent snake antivenom (Neuro Polyvalent Snake Antivenom). PLoS Negl. Trop. Dis. 2012, 6, e1672. [Google Scholar] [CrossRef]
- Oh, A.M.F.; Tan, C.H.; Tan, K.Y.; Quraishi, N.H.; Tan, N.H. Venom proteome of Bungarus sindanus (Sind krait) from Pakistan and in vivo cross-neutralization of toxicity using an Indian polyvalent antivenom. J. Proteom. 2019, 193, 243–254. [Google Scholar] [CrossRef]
- Madhushani, U.; Isbister, G.K.; Tasoulis, T.; Hodgson, W.C.; Silva, A. In-vitro neutralization of the neurotoxicity of coastal taipan venom by Australian polyvalent antivenom: The window of opportunity. Toxins 2020, 12, 690. [Google Scholar] [CrossRef]
- Kuruppu, S.; Smith, A.I.; Isbister, G.K.; Hodgson, W.C. Neurotoxins from Australo-Papuan elapids: A biochemical and pharmacological perpective. Crit. Rev. Toxicol. 2008, 38, 73–86. [Google Scholar] [CrossRef]
- Hodgson, W.C.; Wickramaratna, J.C. In vitro neuromuscular activity of snake venoms. Clin. Exp. Pharmacol. Physiol. 2002, 29, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Hodgson, W.C.; Isbister, G.K. Antivenom for neuromuscular paralysis resulting from snake envenoming. Toxins 2017, 9, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, A.L.; Barfaraz, A.; Thomson, E.; Faiz, A.; Preston, S.; Harris, J.B. Screening of snake venoms for neurotoxic and myotoxic effects using simple in vitro preparations from rodents and chicks. Toxicon 1994, 32, 257–265. [Google Scholar] [CrossRef]
- Liu, C.C.; Lin, C.C.; Hsiao, Y.C.; Wang, P.J.; Yu, J.S. Proteomic characterization of six Taiwanese snake venoms: Identification of species-specific proteins and development of a SISCAPA-MRM assay for cobra venom factors. J. Proteom. 2018, 187, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, Y.; Lee, W.; Xu, X.; Zhang, Y.; Zhao, R.; Zhang, Y.; Wang, W. Venom gland transcriptomes of two elapid snakes (Bungarus multicinctus and Naja atra) and evolution of toxin genes. BMC Genom. 2011, 12, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, H.T.; Höjer, J.; Du, N.T. Clinical features of 60 consecutive ICU-treated patients envenomed by Bungarus multicinctus. Southeast. Asian J. Trop. Med. Public Health 2009, 40, 518–524. [Google Scholar] [PubMed]
- Ha, T.H.; Höjer, J.; Trinh, X.K.; Nguyen, T.D. A controlled clinical trial of a novel antivenom in patients envenomed by Bungarus multicinctus. J. Med. Toxicol. 2010, 6, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.; Cristofori-Armstrong, B.; Rash, L.D.; Hodgson, W.C.; Isbister, G.K. Defining the role of post-synaptic α-neurotoxins in paralysis due to snake envenoming in humans. Cell Mol. Life Sci. 2018. [Google Scholar] [CrossRef]
- Hart, A.J.; Isbister, G.K.; Hodgson, W.C. In vitro neurotoxic effects of Pseudechis spp. venoms: A comparison of avian and murine skeletal muscle preparations. Toxicon 2013, 63, 112–115. [Google Scholar] [CrossRef]
- Hart, A.J.; Isbister, G.K.; O’Donnell, P.; Williamson, N.A.; Hodgson, W.C. Species differences in the neuromuscular activity of post-synaptic neurotoxins from two Australian black snakes (Pseudechis porphyriacus and Pseudechis colletti). Toxicol. Lett. 2013, 219, 262–268. [Google Scholar] [CrossRef]
- Lin, B.; Zhang, J.-R.; Lu, H.-J.; Zhao, L.; Chen, J.; Zhang, H.-F.; Wei, X.-S.; Zhang, L.-Y.; Wu, X.-B.; Lee, W.-H. Immunoreactivity and neutralization study of Chinese Bungarus multicinctus antivenin and lab-prepared anti-bungarotoxin antisera towards purified bungarotoxins and snake venoms. PLoS Negl. Trop. Dis. 2020, 14, e0008873. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Vehicle @ t90 | Bm AV @ t90 | Bm AV @ t50 | Bm AV @ t20 | Washing @ t20 | |
|---|---|---|---|---|---|
| t90 (min) | 26.2 ± 0.9 | 28.0 ± 1.2 | 34.5 ± 1.3 † | 56.2 ± 8.0 *,§ | 26.0 ± 1.0 † |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Q.; Huynh, T.M.; Ng, Y.Z.; Isbister, G.K.; Hodgson, W.C. In Vitro Neurotoxicity of Chinese Krait (Bungarus multicinctus) Venom and Neutralization by Antivenoms. Toxins 2021, 13, 49. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010049
Liang Q, Huynh TM, Ng YZ, Isbister GK, Hodgson WC. In Vitro Neurotoxicity of Chinese Krait (Bungarus multicinctus) Venom and Neutralization by Antivenoms. Toxins. 2021; 13(1):49. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010049
Chicago/Turabian StyleLiang, Qing, Tam Minh Huynh, Yen Zhi Ng, Geoffrey K. Isbister, and Wayne C. Hodgson. 2021. "In Vitro Neurotoxicity of Chinese Krait (Bungarus multicinctus) Venom and Neutralization by Antivenoms" Toxins 13, no. 1: 49. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010049