Populations of Helicoverpa zea (Boddie) in the Southeastern United States are Commonly Resistant to Cry1Ab, but Still Susceptible to Vip3Aa20 Expressed in MIR 162 Corn

 ,
,
Abstracts
1. Introduction
2. Results
2.1. Resistance to Cry1Ab in Helicoverpa zea is Common Across the Southeastern United States
2.2. Field Populations of Helicoverpa zea in the Southeastern United States Are Still Susceptible to Vip3Aa20
2.3. There Are No Significant Relationships Between the Susceptibilities of Helicoverpa Zea Populations to Cry1Ab and Vip3Aa20 Proteins
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insect Sources
5.2. Insect Rearing
5.3. Diet-Overlay Bioassays
5.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- US EPA. White Paper on Resistance in Lepidopteran pests of Bacillus thuringiensis (Bt) Plant Incorporated Protectants in the United States. 2018. Available online: https://www.epa.gov/sites/production/files/2018-07/documents/position_paper_07132018.pdf (accessed on 22 December 2020).
- Reay-Jones, F.P.F. Pest status and management of corn earworm (Lepidoptera: Noctuidae) in field corn in the United States. J. Integr. Pest Manag. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- National Agricultural Statistics Service (NASS). Acreage; USDA: Washington, DC, USA, 2020; ISBN 1949-1522. Available online: https://www.nass.usda.gov/Publications/Todays_Reports/reports/acrg0620.pdf (accessed on 20 December 2020).
- FIFRA Scientific Advisory Panel. Report of Subpanel on Bacillus thuringiensis (Bt) Plant-Pesticides and Resistance Management. EPA SAP Report; 1998. Available online: https://archive.epa.gov/scipoly/sap/meetings/web/pdf/finalfeb.pdf (accessed on 22 December 2020).
- Sandstrom, M.A.; Changnon, D.; Flood, B.R. Improving our understanding of Helicoverpa zea migration in the Midwest: Assessment of source populations. Plant Health Prog. 2007, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Dively, G.P.; Venugopal, P.D.; Finkenbinder, C. Field-evolved resistance in corn earworm to Cry proteins expressed by transgenic sweet corn. PLoS ONE 2016, 11, e0169115. [Google Scholar] [CrossRef] [PubMed]
- Reisig, D.D.; Huseth, A.S.; Bacheler, J.S.; Aghaee, M.A.; Braswell, L.; Burrack, H.J.; Flanders, K.; Greene, J.K.; Herbert, D.A.; Jacobson, A. Long-term empirical and observational evidence of practical Helicoverpa zea resistance to cotton with pyramided Bt toxins. J. Econ. Entomol. 2018, 111, 1824–1833. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Guo, J.; Brown, S.; Head, G.P.; Price, P.A.; Paula-Moraes, S.; Ni, X.; Dimase, M.; Huang, F. Field-evolved resistance of Helicoverpa zea (Boddie) to transgenic maize expressing pyramided Cry1A. 105/Cry2Ab2 proteins in northeast Louisiana, the United States. J. Invertebr. Pathol. 2019, 163, 11–20. [Google Scholar] [CrossRef]
- Yang, F.; González, J.C.; Williams, J.; Cook, D.C.; Gilreath, R.T.; Kerns, D.L. Occurrence and ear damage of Helicoverpa zea on transgenic Bacillus thuringiensis maize in the field in Texas, US and its susceptibility to Vip3A protein. Toxins 2019, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Storer, N.P.; Van Duyn, J.W.; Kennedy, G.G. Life history traits of Helicoverpa zea (Lepidoptera: Noctuidae) on non-Bt and Bt transgenic corn hybrids in eastern North Carolina. J. Econ. Entomol. 2001, 94, 1268–1279. [Google Scholar] [CrossRef]
- Buntin, D.G.; All, J.N.; Lee, R.D.; Wilson, D.M. Plant-incorporated Bacillus thuringiensis resistance for control of fall armyworm and corn earworm (Lepidoptera: Noctuidae) in corn. J. Econ. Entomol. 2004, 97, 1603–1611. [Google Scholar] [CrossRef]
- Reisig, D.D.; Reay-Jones, F.P.F. Inhibition of Helicoverpa zea growth by transgenic corn expressing Bt toxins. Environ. Entomol. 2015, 44, 1275–1285. [Google Scholar] [CrossRef]
- Estruch, J.J.; Warren, G.W.; Mullins, M.A.; Nye, G.J.; Craig, J.A.; Koziel, M.G. Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proc. Natl. Acad. Sci. USA 1996, 93, 5389–5394. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Walters, F.S.; Hart, H.; Palekar, N.; Chen, J.S. The mode of action of the Bacillus thuringiensis vegetative insecticidal protein Vip3A differs from that of Cry1Ab δ-endotoxin. Appl. Environ. Microbiol. 2003, 69, 4648–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, R.E.; Marcus, M.A.; Gould, F.; Bradley, J.R., Jr.; Van Duyn, J.W. Cross-resistance responses of Cry1Ac-selected Heliothis virescens (Lepidoptera: Noctuidae) to the Bacillus thuringiensis protein Vip3A. J. Econ. Entomol. 2007, 100, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Yang, F.; Dangal, V.; Huang, F. Larval survival and plant injury of Cry1F-susceptible, -resistant, and -heterozygous fall armyworm (Lepidoptera: Noctuidae) on non-Bt and Bt corn containing single or pyramided genes. Crop Prot. 2014, 59, 22–28. [Google Scholar] [CrossRef]
- Santos-Amaya, O.F.; Rodrigues, J.V.; Souza, T.C.; Tavares, C.S.; Campos, S.O.; Guedes, R.N.; Pereira, E.J. Resistance to dual-gene Bt maize in Spodoptera frugiperda: Selection, inheritance, and cross-resistance to other transgenic events. Sci. Rep. 2015, 5, 18243. [Google Scholar] [CrossRef] [PubMed]
- Chakroun, M.; Banyuls, N.; Bel, Y.; Escriche, B.; Ferré, J. Bacterial vegetative insecticidal proteins (VIP) from entomopathogenic bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 329–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- US-EPA (US Environmental Protection Agency). Biopesticides Registration Action Document: Bacillus thuringiensis Vip3Aa20 Insecticidal Protein and the Genetic Material Necessary for Its Production (via Elements of Vector pNOV1300) in Event MIR162 Maize. 2009. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/decision_PC-006599_3-Apr-09.pdf (accessed on 24 April 2020).
- Ali, M.I.; Luttrell, R.G. Susceptibility of Helicoverpa zea and Heliothis virescens (Lepidoptera: Noctuidae) to Vip3A insecticidal toxin expressed in VipCot™ cotton. J. Invertebr. Pathol. 2011, 108, 76–84. [Google Scholar] [CrossRef]
- Burkness, E.C.; Dively, G.; Patton, T.; Morey, A.C.; Hutchison, W.D. Novel Vip3A Bacillus thuringiensis (Bt) maize approaches high-dose efficacy against Helicoverpa zea (Lepidoptera: Noctuidae) under field conditions. GM Crop. 2010, 1, 1–7. [Google Scholar]
- Palekar, N.; Kurtz, R.W.; Walters, F.S.; O’Reilly, D. Relative efficacy of cotton events expressing Cry1Ab and Vip3A against cotton bollworm and tobacco budworm (Lepidoptera: Noctuidae). J. Cotton Sci. 2011, 15, 100–108. [Google Scholar]
- Yang, F.; Kerns, D.L.; Leonard, B.R.; Oyediran, I.; Burd, T.; Niu, Y.; Huang, F. Performance of Agrisure® Viptera™ 3111 corn against Helicoverpa zea (Lepidoptera: Noctuidae) in seed mixed plantings. Crop Prot. 2015, 69, 77–82. [Google Scholar] [CrossRef]
- Dimase, M.; Oyediran, I.; Brown, S.; Walker, W.; Guo, J.; Yu, W.; Zhang, Y.; Chen, J.; Wen, Z.; Huang, F. Larval movement and survival of Helicoverpa zea (Boddie) in seed blends of non-Bt and Bt maize containing Agrisure Viptera® trait: Implications for resistance management. Crop Prot. 2020. [Google Scholar] [CrossRef]
- Guo, J.; Oyediran, I.; Rice, M.E.; Brown, S.; Dimase, M.; Lin, S.; Walker, W.; Yu, W.; Niu, Y.; Huang, F. Seed blends of pyramided Cry/Vip maize reduce Helicoverpa zea populations from refuge ears. J. Pest Sci. 2020. [Google Scholar] [CrossRef]
- Huang, F. Resistance of the fall armyworm, Spodoptera frugiperda (J. E. Smith), to transgenic Bacillus thuringiensis Cry1F corn in the Americas: Lessons and implications for Bt corn IRM in China. Insect Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; Santos, A.C.; Omoto, C. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Huang, F.; Qureshi, J.A.; Meagher, R.L., Jr.; Reisig, D.D.; Head, G.P.; Andow, D.A.; Ni, X.; Kerns, D.; Buntin, G.D.; Niu, Y.; et al. Cry1F resistance in fall armyworm Spodoptera frugiperda: Single gene versus pyramided Bt maize. PLoS ONE 2014, 9, e112958. [Google Scholar] [CrossRef] [Green Version]
- Chandrasena, D.; Signorini, A.M.; Abratti, G.; Storer, N.G.; Olaciregui, M.L.; Alves, A.P.; Pilcher, C.D. Characterization of field-evolved resistance to Bacillus thuringiensis derived Cry1Fδ endotoxin in Spodoptera frugiperda populations from Argentina. Pest Manag. Sci. 2018, 74, 746–754. [Google Scholar] [CrossRef] [Green Version]
- Leite, N.A.; Pereira, R.M.; Durigan, M.R.; Amado, D.; Fatoretto, J.; Medeiros, F.C.L.; Omoto, C. Susceptibility of Brazilian populations of Helicoverpa armigera and Helicoverpa zea (Lepidoptera: Noctuidae) to Vip3Aa20. J. Econ. Entomol. 2018, 111, 399–404. [Google Scholar] [CrossRef]
- Huang, F. Detection and monitoring of insect resistance to transgenic Bt crops. Insect Sci. 2006, 13, 73–84. [Google Scholar] [CrossRef]
- Luttrell, R.G.; Wan, L.; Knighten, K. Variation in susceptibility of noctuid (Lepidoptera) larvae attacking cotton and soybean to purified endotoxin proteins and commercial formulations of Bacillus thuringiensis. J. Econ. Entomol. 1999, 92, 21–32. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Spencer, T.; Nearman, J. Baseline susceptibility of the corn earworm (Lepidoptera: Noctuidae) to the Cry1Ab toxin from Bacillus thuringiensis. J. Econ. Entomol. 2000, 93, 1265–1268. [Google Scholar] [CrossRef] [Green Version]
- Anilkumar, K.J.; Rodrigo-Simón, A.; Ferré, J.; Pusztai-Carey, M.; Sivasupramaniam, S.; Moar, W.J. Production and characterization of Bacillus thuringiensis Cry1Ac-resistant cotton bollworm Helicoverpa zea (Boddie). Appl. Environ. Microbiol. 2008, 74, 462–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welch, K.L.; Unnithan, G.C.; Degain, B.A.; Wei, J.; Zhang, J.; Li, X.; Tabashnik, B.E.; Carrière, Y. Cross-resistance to toxins used in pyramided Bt crops and resistance to Bt sprays in Helicoverpa zea. J. Invertebr. Pathol. 2015, 132, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Carrière, Y. Evaluating Cross-resistance Between Vip and Cry Toxins of Bacillus thuringiensis. J. Econ. Entomol. 2020, 113, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, R.G.; Jackson, R.E. Helicoverpa zea and Bt cotton in the United States. GM Crops Food 2012, 3, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Siebert, M.W.; Nolting, S.P.; Hendrix, W.; Dhavala, S.; Craig, C.; Leonard, B.R.; Stewart, S.D.; All, J.; Musser, F.R.; Buntin, G.D.; et al. Evaluation of corn hybrids expressing Cry1F, Cry1A. 105, Cry2Ab2, Cry34Ab1/Cry35Ab1, and Cry3Bb1 against southern United States insect pests. J. Econ. Entomol. 2012, 105, 1825–1834. [Google Scholar] [CrossRef]
- Rule, D.M.; Nolting, S.P.; Prasifka, P.L.; Storer, N.P.; Hopkins, B.W.; Scherder, E.F.; Siebert, M.W.; Hendrix, W.H., III. Efficacy of pyramided Bt proteins Cry1F, Cry1A.105, and Cry2Ab2 expressed in SmartStax corn hybrids against lepidopteran insect pests in the northern United States. J. Econ. Entomol. 2014, 107, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Bilbo, T.R.; Reay-Jones, F.P.; Reisig, D.D.; Greene, J.K. Susceptibility of corn earworm (Lepidoptera: Noctuidae) to Cry1A. 105 and Cry2Ab2 in North and South Carolina. J. Econ. Entomol. 2019, 112, 1845–1857. [Google Scholar] [CrossRef]
- Yu, W.; Lin, S.; Dimase, M.; Niu, Y.; Brown, S.; Head, G.P.; Price, P.A.; Reay-Jones, F.P.F.; Cook, D.; Reisig, D.; et al. Extended investigation of the field-evolved resistance of the corn earworm (Lepidoptera: Noctuidae) to Bacillus thuringiensis Cry1A.105 and Cry2Ab2 proteins in the southeastern United States. J. Invertebr. Path. 2021. submitted. [Google Scholar]
- Adamczyk, J.J.; Mahaffey, J.S. Efficacy of Vip3A and Cry1Ab transgenic traits in cotton against various lepidopteran pests. Fla. Entomol. 2008, 91, 570–575. [Google Scholar]
- Bommireddy, P.L.; Leonard, B.R.; Temple, J.; Price, P.; Emfinger, K.; Cook, D.; Hardke, J.T. Field performance and seasonal efficacy profiles of transgenic cotton lines expressing Vip3A and VipCot against Helicoverpa zea (Boddie) and Heliothis virescens (F.). J. Cotton Sci. 2011, 15, 251–259. [Google Scholar]
- Hernández-Rodríguez, C.S.; Hernández-Martínez, P.; Van Rie, J.; Escriche, B.; Ferré, J. Shared midgut binding sites for Cry1A.105, Cry1Aa, Cry1Ab, Cry1Ac and Cry1Fa proteins from Bacillus thuringiensis in two important corn pests, Ostrinia nubilalis and Spodoptera frugiperda. PLoS ONE 2013, 8, e68164. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, D.; Salmeron, E.; Horikoshi, R.J.; Bernardi, O.; Dourado, P.M.; Carvalho, R.A.; Martinelli, S.; Head, G.P.; Omoto, C. Cross-resistance between Cry1 proteins in fall armyworm (Spodoptera frugiperda) may affect the durability of current pyramided Bt maize hybrids in Brazil. PLoS ONE 2015, 10, e0140130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Y.; Head, G.P.; Price, P.A.; Huang, F. Performance of Cry1A.105-selected fall armyworm (Lepidoptera: Noctuidae) on transgenic maize plants containing single or pyramided Bt genes. Crop Prot. 2016, 88, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Vyavhare, S.S.; Kerns, D. Bt cotton. Texas AgriLife Extension. ENTO-067. 2017. Available online: http://lubbock.tamu.edu/files/2017/03/Bt-Cotton_ENTO067.pdf (accessed on 24 April 2020).
- DiFonzo, C. The Handy Bt Trait Table for U.S. Corn Production. 2020. Available online: https://agrilife.org/lubbock/files/2020/02/BtTraitTable_FEB_2020.pdf (accessed on 22 December 2020).
- Huang, F.; Leonard, B.R.; Gable, R.H. Comparative susceptibility of European corn borer, southwestern corn borer, and sugarcane borer (Lepidoptera: Crambidae) to Cry1Ab protein in a commercial Bt-corn hybrid. J. Econ. Entomol. 2006, 99, 194–202. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- SAS Institute. SAS/STAT User’s, 3rd ed.; SAS Institute Inc.: Cary, NC, USA, 2010. [Google Scholar]


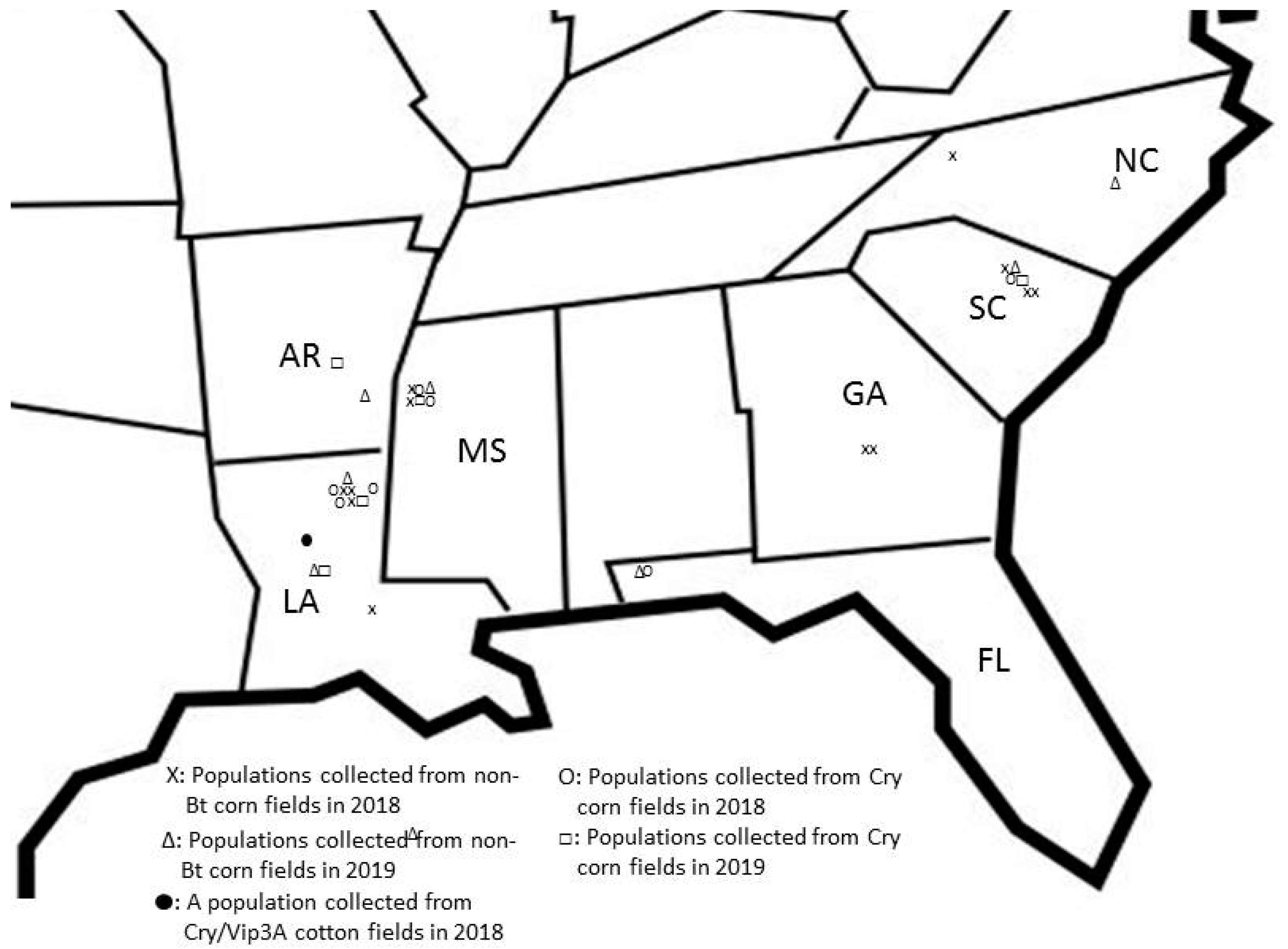{kind=link}
| Insect Populations a | Collection Locations: County/Parish, State b | Host Sources c | No. Larvae Assayed | Slope ± SE | LC50 (95% CI) d (µg/cm2) | χ2 | p-Value | Resistance ratio e | % Larval Survivorship at 10 µg/cm2 |
|---|---|---|---|---|---|---|---|---|---|
| BZ | Benzon Research Inc. | Laboratory | 1699 | 1.29 ± 0.12 | 0.21 (0.15, 0.29) | 304.09 | <0.0001 | n/a | 0.0 ± 0.0 |
| Insect populations collected during 2018 | |||||||||
| LA-BR-NBt | East Baton Rouge, LA | Non-Bt corn | 324 | 1.30 ± 0.28 | 3.51 (2.36, 5.78) | 9.20 | 0.5128 | 17 | 26.7 ± 3.7 |
| LA-WB-NBt-1 | Franklin, LA | Non-Bt corn | 983 | 0.99 ± 0.18 | 9.10 (4.78, 29.8) | 57.30 | <0.0001 | 43 | 50.8 ± 8.1 |
| LA-WB-NBt-2 | Franklin, LA | Non-Bt corn | 368 | 1.85 ± 0.32 | 9.10 (6.74, 14.56) | 13.00 | 0.2239 | 43 | 50.2 ± 3.1 |
| LA-WB-NBt-3 | Franklin, LA | Non-Bt corn | 414 | 1.48 ± 0.30 | 7.71 (5.30, 10.5) | 9.23 | 0.5106 | 37 | 18.6 ± 1.7 |
| LA-WB-VT2P-1 | Franklin, LA | VT-2P corn | 419 | 0.93 ± 0.20 | 14.4 (7.4, 49.0) | 36.18 | 0.0043 | 69 | 56.7 ± 4.3 |
| LA-SJ-VT2P | Tensas, LA | VT-2P corn | 394 | 2.81 ± 0.35 | 12.4 (10.0, 15.3) | 15.57 | 0.1126 | 59 | 55.9 ± 5.7 |
| LA-WB-SMT | Franklin, LA | SmartStax corn | 255 | n/a | >31.6 | n/a | n/a | >150 | 73.0 ± 4.1 |
| LA-GT-WS3 | Grant, LA | WideStrike-3 cotton | 485 | n/a | >10 | n/a | n/a | >48 | 51.8 ± 1.0 |
| MS-ST-NBt-1 | Washington, MS | Non-Bt corn | 374 | 0.94 ± 0.12 | 1.84 (1.27, 2.82) | 25.73 | 0.1062 | 9 | 24.2 ± 4.8 |
| MS-LL-NBt | Washington, MS | Non-Bt corn | 360 | 0.78 ± 0.14 | 2.77 (1.50, 6.88) | 28.59 | 0.0536 | 13 | 24.7 ± 5.8 |
| MS-ST-VT2P-1 | Washington, MS | VT-2P corn | 371 | n/a | >10 | n/a | n/a | >48 | 54.7 ± 6.9 |
| MS-LL-Intra | Washington, MS | Intrasect corn | 287 | 0.82 ± 0.24 | 10.75 (5.76, 64.5) | 15.17 | 0.1261 | 51 | 51.8 ± 10.6 |
| GA-TF-NBt-1 | Tift, GA | Non-Bt corn | 228 | n/a | >10 | n/a | n/a | >48 | 74.4 ± 10.2 |
| GA-TF-NBt-2 | Tift, GA | Non-Bt corn | 439 | n/a | >10 | n/a | n/a | >48 | 73.5 ± 6.5 |
| SC-FR-NBt-1 | Florence, SC | Non-Bt corn | 362 | n/a | >10 | n/a | n/a | >48 | 66.9 ± 7.6 |
| SC-FR-NBt-2 | Florence, SC | Non-Bt corn | 342 | n/a | >10 | n/a | n/a | >48 | 82.9 ± 1.6 |
| SC-DL-NBt-1 | Darlington, SC | Non-Bt corn | 384 | n/a | >10 | n/a | n/a | >48 | 59.4 ± 11.0 |
| SC-DL-VT2P-1 | Darlington, SC | VT-2P corn | 341 | 0.67 ± 0.19 | 4.61 (1.84, 38.1) | 45.00 | 0.0004 | 22 | 44.1 ± 15.6 |
| NC-CD-NBt | Caldwell, NC | Non-Bt corn | 349 | 1.25 ± 0.20 | 3.49 (2.39, 5.79) | 16.68 | 0.2736 | 17 | 44.2 ± 8.7 |
| FL-SR-VT2P | Jay, FL | VT-2P corn | 377 | 0.79 ± 0.12 | 1.84 (1.09, 2.97) | 16.77 | 0.2687 | 9 | n/a |
| Insect populations collected during 2019 | |||||||||
| LA-WB-NBt-4 | Franklin, LA | Non-Bt corn | 437 | n/a | >10 | n/a | n/a | >48 | 68.7 ± 6.8 |
| LA-AX-NBt | Rapides, LA | Non-Bt corn | 441 | n/a | >10 | n/a | n/a | >48 | 79.4 ± 6.1 |
| LA-WB-VT2P-2 | Franklin, LA | VT-2P corn | 478 | n/a | >10 | n/a | n/a | >48 | 80.6 ± 5.6 |
| LA-AX-VT2P | Rapides, LA | VT-2P corn | 474 | 0.79 ± 0.11 | 1.00 (0.60, 1.77) | 31.79 | 0.0810 | 5 | 27.0 ± 1.6 |
| MS-ST-NBt-2 | Tensas, LA | Non-Bt corn | 512 | 1.02 ± 0.16 | 1.03 (0.56, 2.09) | 84.87 | <0.0001 | 5 | 14.6 ± 4.0 |
| MS-ST-VT2P-2 | Tensas, LA | VT-2P corn | 447 | 0.75 ± 0.11 | 2.32 (1.48, 4.11) | 24.70 | 0.1334 | 11 | 37.9 ± 8.2 |
| AR-TR-NBt | Desha, AR | Non-Bt corn | 510 | 0.83 ± 0.09 | 0.46 (0.31, 0.68) | 19.87 | 0.5913 | 2 | 9.7 ± 1.9 |
| SC-DL-VT2P-2 | Darlington, SC | VT-2P corn | 511 | 0.65 ± 0.11 | 5.99 (3.24, 16.54) | 21.74 | 0.2438 | 29 | 34.9 ± 6.6 |
| NC-HA-NBt | Harnett, NC | Non-Bt corn | 506 | 1.10 ± 0.18 | 8.36 (5.43, 16.59) | 13.36 | 0.4981 | 40 | 50.8 ± 9.3 |
| FL-SR-NBt | Jay, FL | Non-Bt corn | 502 | 1.15 ± 0.25 | 0.40 (0.19, 0.72) | 33.50 | 0.0024 | 2 | 3.9 ± 3.9 |
| Insect Population Groups | Mean Survivorship ± SEM% | t-Tests |
|---|---|---|
| 12 populations collected from non-Bt fields in 2018 | 49.7 ± 6.4 | |
| 7 populations collected from Bt fields in 2018 | 55.4 ± 3.3 | t17 = −0.64, p = 0.5284 |
| 6 populations collected from non-Bt fields in 2019 | 37.9 ± 13.3 | |
| 4 populations collected from Bt fields in 2019 | 45.1 ± 12.1 | t8 = −0.38, p = 0.7155 |
| 19 populations collected from non-Bt and Bt fields in 2018 | 51.8 ± 4.2 | |
| 10 populations collected from non-Bt and Bt fields in 2019 | 40.8 ± 8.9 | t27 = 1.28, p = 0.2125 |
| 18 populations collected from non-Bt fields in 2018 and 2019 | 45.8 ± 6.1 | |
| 11 populations collected from Bt fields in 2018 and 2019 | 51.7 ± 4.7 | t27 = −0.68, p = 0.5006 |
| Insect Populations a | Collection Locations: County/Parish, State b | Host Sources c | No. Larvae Assayed | Slope ± SE | LC50 (95%CI) (µg/cm2) | χ2 | p-Value | Susceptibility Ratio d |
|---|---|---|---|---|---|---|---|---|
| BZ | Benzon Research, Inc. | Laboratory | 1652 | 2.47 ± 0.39 | 0.40 (0.30, 0.51) | 113.0 | <0.0001 | - |
| Insect populations collected during 2018 | ||||||||
| LA-BR-NBt | East Baton Rouge, LA | Non-Bt corn | 365 | 4.41 ± 0.60 | 0.20 (0.17, 0.23) | 12.49 | 0.2535 | −2.0 |
| LA-WB-NBt-1 | Franklin, LA | Non-Bt corn | 1004 | 2.43 ± 0.24 | 0.96 (0.78, 1.12) | 23.68 | 0.0501 | 2.4 |
| LA-WB-NBt-2 | Franklin, LA | Non-Bt corn | 383 | 2.14 ± 0.54 | 0.13 (0.06, 0.22) | 50.73 | <0.0001 | –3.1 |
| LA-WB-NBt-3 | Franklin, LA | Non-Bt corn | 367 | 1.92 ± 0.23 | 0.21 (0.16, 0.27) | 9.85 | 0.7730 | −1.9 |
| LA-WB-SMT | Franklin, LA | SmartStax corn | 350 | 2.19 ± 0.23 | 0.37 (0.26, 0.41) | 14.19 | 0.7163 | −1.1 |
| LA-GT-WS3 | Grant, LA | WideStrike-3 cotton | 370 | 1.71 ± 0.25 | 0.92 (0.59, 1.31) | 22.11 | 0.0763 | 2.3 |
| MS-ST-NBt-1 | Washington, MS | Non-Bt corn | 362 | 1.43 ± 0.24 | 0.32 (0.17, 0.51) | 43.94 | 0.0006 | −1.3 |
| MS-LL-NBt | Washington, MS | Non-Bt corn | 343 | 1.89 ± 0.47 | 0.14 (0.05, 0.23) | 45.11 | <0.0001 | −2.9 |
| MS-ST-VT2P-1 | Washington, MS | VT-2P corn | 375 | 3.16 ± 0.48 | 0.48 (0.36, 0.65) | 30.40 | 0.0067 | 1.2 |
| MS-LL-Intra | Washington, MS | Intrasect corn | 358 | 1.96 ± 0.26 | 0.46 (0.32, 0.66) | 32.91 | 0.0116 | 1.2 |
| GA-TF-NBt-1 | Tift, GA | Non-Bt corn | 369 | 2.34 ± 0.26 | 0.42 (0.34, 0.51) | 10.86 | 0.9003 | 1.1 |
| GA-TF-NBt-2 | Tift, GA | Non-Bt corn | 446 | n/a | <0.0316 | n/a | n/a | <−11.5 |
| SC-FR-NBt-1 | Florence, SC | Non-Bt corn | 368 | 2.55 ± 0.41 | 0.30 (0.21, 0.41) | 35.75 | 0.0049 | −1.3 |
| SC-FR-NBt-2 | Florence, SC | Non-Bt corn | 365 | 2.22 ± 0.24 | 0.34 (0.27, 0.42) | 11.61 | 0.6379 | −1.2 |
| SC-DL-NBt-1 | Darlington, SC | Non-Bt corn | 438 | 4.91 ± 0.59 | 0.88 (0.78, 1.01) | 4.90 | 0.9872 | 2.2 |
| SC-DL-VT2P-1 | Darlington, SC | VT-2P corn | 367 | 2.05 ± 0.33 | 0.31 (0.21, 0.45) | 29.69 | 0.0084 | −1.3 |
| NC-CD-NBt | Caldwell, NC | Non-Bt corn | 369 | 1.79 ± 0.17 | 0.48 (0.37, 0.61) | 23.69 | 0.1656 | 1.2 |
| FL-SR-VT2P | Jay, FL | VT-2P corn | 376 | 2.46 ± 0.68 | 0.17 (0.07, 0.28) | 90.42 | <0.0001 | −2.4 |
| Insect populations collected during 2019 | ||||||||
| LA-WB-NBt-4 | Franklin, LA | Non-Bt corn | 511 | 2.27 ± 0.21 | 0.25 (0.20, 0.30) | 14.66 | 0.6853 | −1.6 |
| LA-AX-NBt | Rapides, LA | Non-Bt corn | 497 | 2.57 ± 0.26 | 1.34 (1.10, 1.63) | 17.79 | 0.2165 | 3.4 |
| LA-WB-VT2P-2 | Franklin, LA | VT-2P corn | 256 | - | <0.316 | - | - | <1.3 |
| LA-AX-VT2P | Rapides, LA | VT-2P corn | 512 | 2.98 ± 0.30 | 0.06 (0.05, 0.07) | 24.04 | 0.1539 | −6.7 |
| MS-ST-VT2P-2 | Washington, MS | VT-2P corn | 512 | 1.49 ± 0.14 | 0.21 (0.16, 0.28) | 21.70 | 0.2455 | −1.9 |
| AR-TR-NBt | Desha, AR | Non-Bt corn | 500 | 1.21 ± 0.18 | 0.04 (0.02, 0.06) | 33.71 | 0.0137 | −10.0 |
| AR-LK-VT2P | Lonoke, AR | VT-2P corn | 507 | 1.86 ± 0.21 | 3.60 (2.84, 4.69) | 6.51 | 0.9520 | 9.0 |
| SC-DL-NBt-2 | Darlington, SC | Non-Bt corn | 1015 | 1.95 ± 0.19 | 1.32 (1.03, 1.71) | 93.73 | <0.0001 | 3.3 |
| SC-DL-VT2P-2 | Darlington, SC | VT-2P corn | 511 | 1.25 ± 0.13 | 0.65 (0.49, 0.96) | 15.31 | 0.6409 | 1.6 |
| NC-HA-NBt | Harnett, NC | Non-Bt corn | 511 | 3.07 ± 0.33 | 0.08 (0.06, 0.09) | 18.20 | 0.1977 | −5.0 |
| FL-SR-NBt | Jay, FL | Non-Bt corn | 509 | 1.37 ± 0.14 | 0.12 (0.09. 0.17) | 41.34 | 0.0287 | −3.3 |
| Insect Population Groups | Mean LC50 ± SEM (µg/cm2) | t-Tests |
|---|---|---|
| 12 populations collected from non-Bt fields in 2018 | 0.37 ± 0.08 | |
| 6 populations collected from Bt fields in 2018 | 0.45 ± 0.10 | t17 = −0.60, p = 0.5540 |
| 6 populations collected from non-Bt fields in 2019 | 0.53 ± 0.26 | |
| 5 populations collected from Bt fields in 2019 | 0.97 ± 0.67 | t8 = −0.67, p = 0.5221 |
| 18 populations collected from non-Bt and Bt fields in 2018 | 0.42 ± 0.06 | |
| 11 populations collected from non-Bt and Bt fields in 2019 | 0.73 ± 0.32 | t27 = −1.26, p = 0.2175 |
| 18 populations collected from non-Bt fields in 2018 and 2019 | 0.42 ± 0.10 | |
| 11 populations collected from Bt fields in 2018 and 2019 | 0.69 ± 0.30 | t27 = −1.01, p = 0.3234 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Y.; Oyediran, I.; Yu, W.; Lin, S.; Dimase, M.; Brown, S.; Reay-Jones, F.P.F.; Cook, D.; Reisig, D.; Thrash, B.; et al. Populations of Helicoverpa zea (Boddie) in the Southeastern United States are Commonly Resistant to Cry1Ab, but Still Susceptible to Vip3Aa20 Expressed in MIR 162 Corn. Toxins 2021, 13, 63. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010063
Niu Y, Oyediran I, Yu W, Lin S, Dimase M, Brown S, Reay-Jones FPF, Cook D, Reisig D, Thrash B, et al. Populations of Helicoverpa zea (Boddie) in the Southeastern United States are Commonly Resistant to Cry1Ab, but Still Susceptible to Vip3Aa20 Expressed in MIR 162 Corn. Toxins. 2021; 13(1):63. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010063
Chicago/Turabian StyleNiu, Ying, Isaac Oyediran, Wenbo Yu, Shucong Lin, Marcelo Dimase, Sebe Brown, Francis P. F. Reay-Jones, Don Cook, Dominic Reisig, Ben Thrash, and et al. 2021. "Populations of Helicoverpa zea (Boddie) in the Southeastern United States are Commonly Resistant to Cry1Ab, but Still Susceptible to Vip3Aa20 Expressed in MIR 162 Corn" Toxins 13, no. 1: 63. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010063