Fusarium verticillioides and Aspergillus flavus Co-Occurrence Influences Plant and Fungal Transcriptional Profiles in Maize Kernels and In Vitro

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
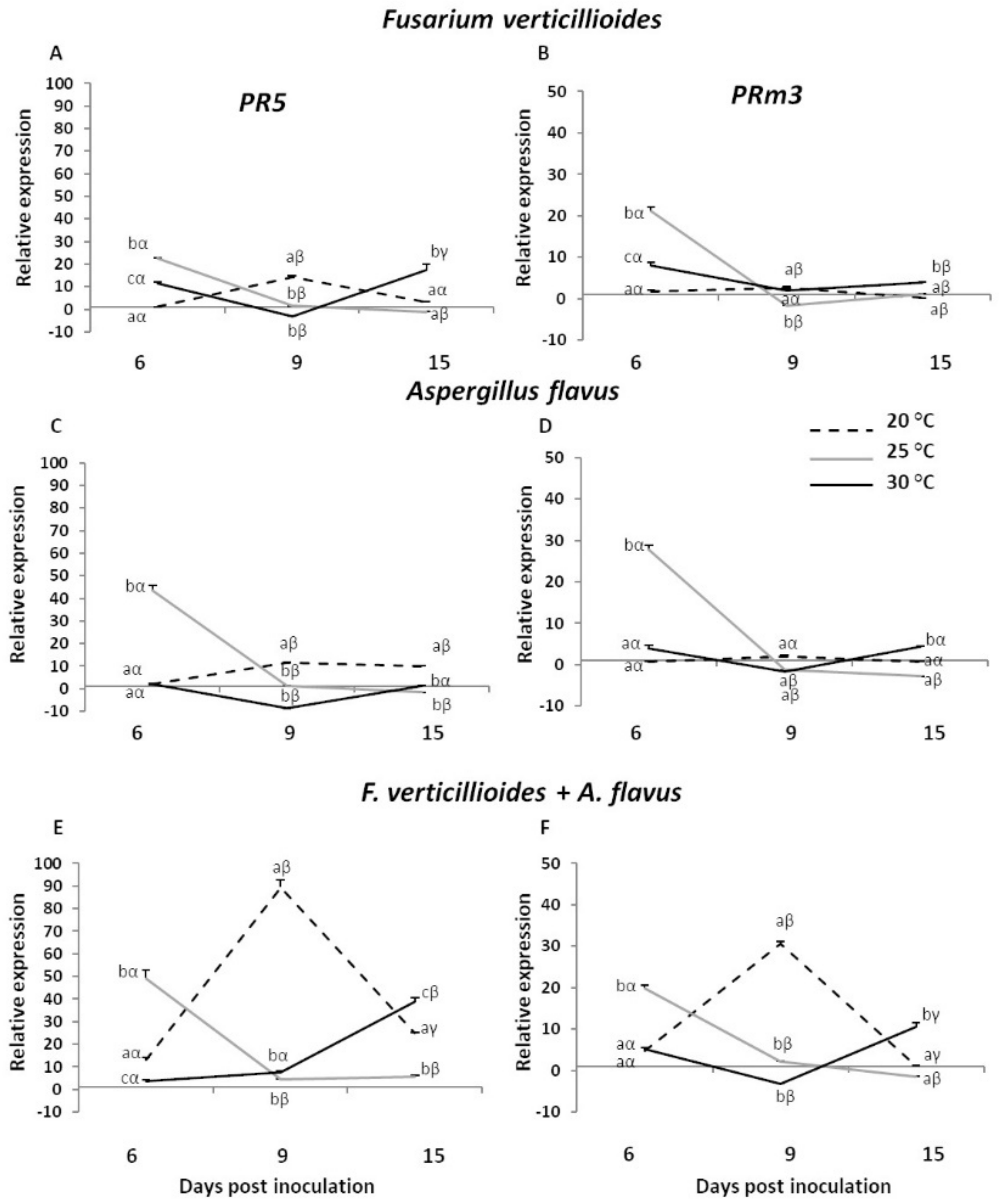
2.1. Expression Analysis of Maize Pathogenesis-Related Genes in Infected Kernels
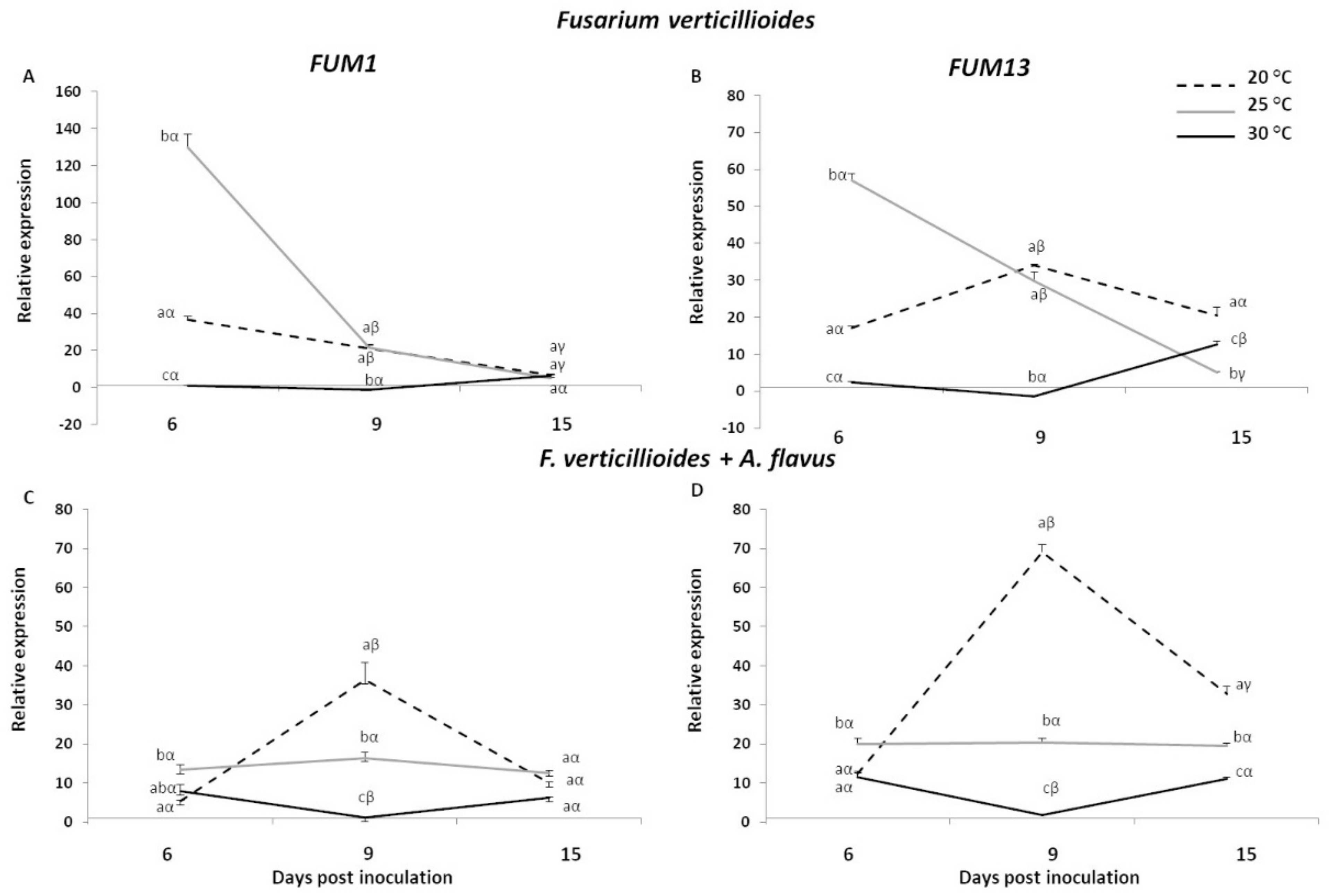
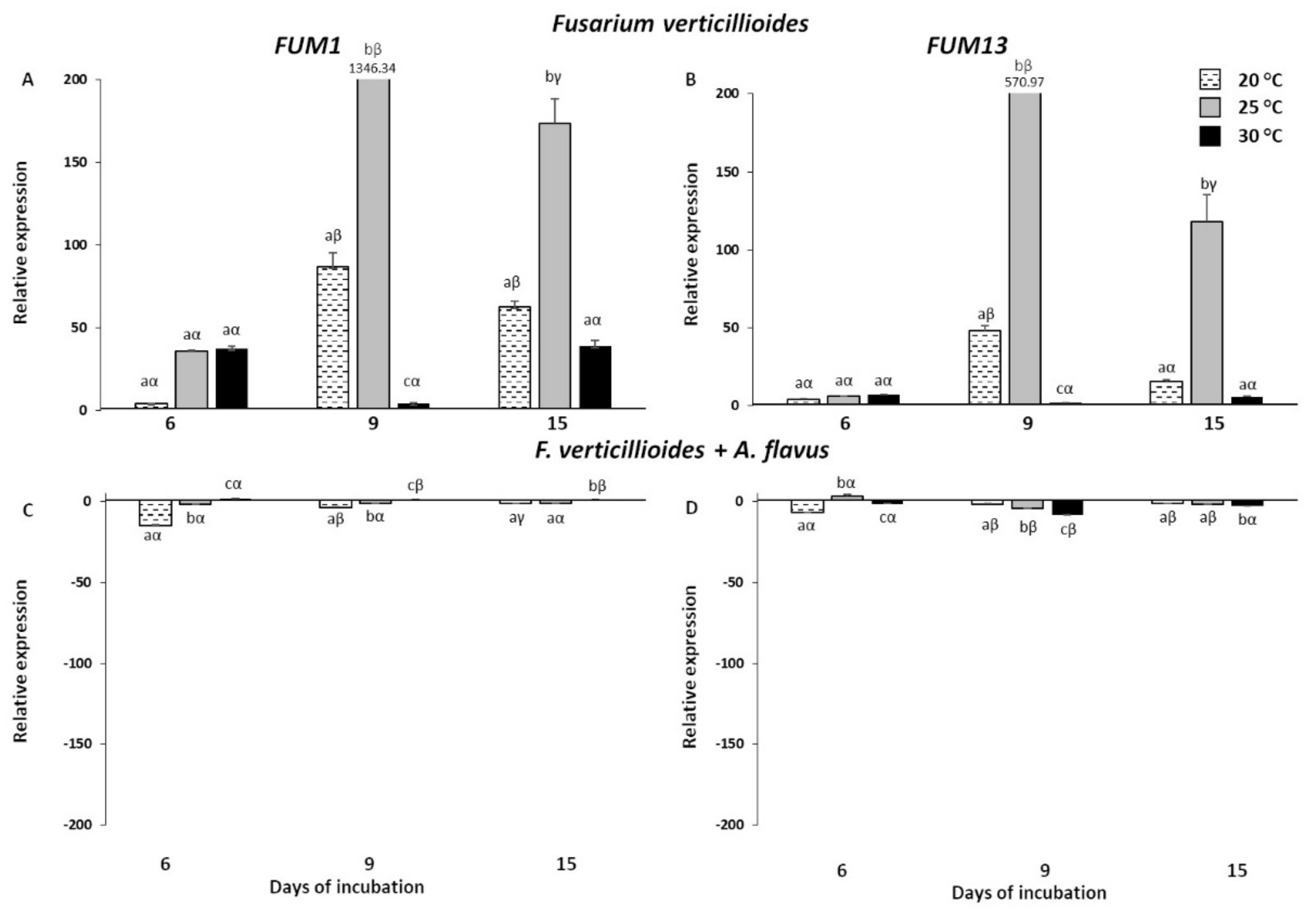
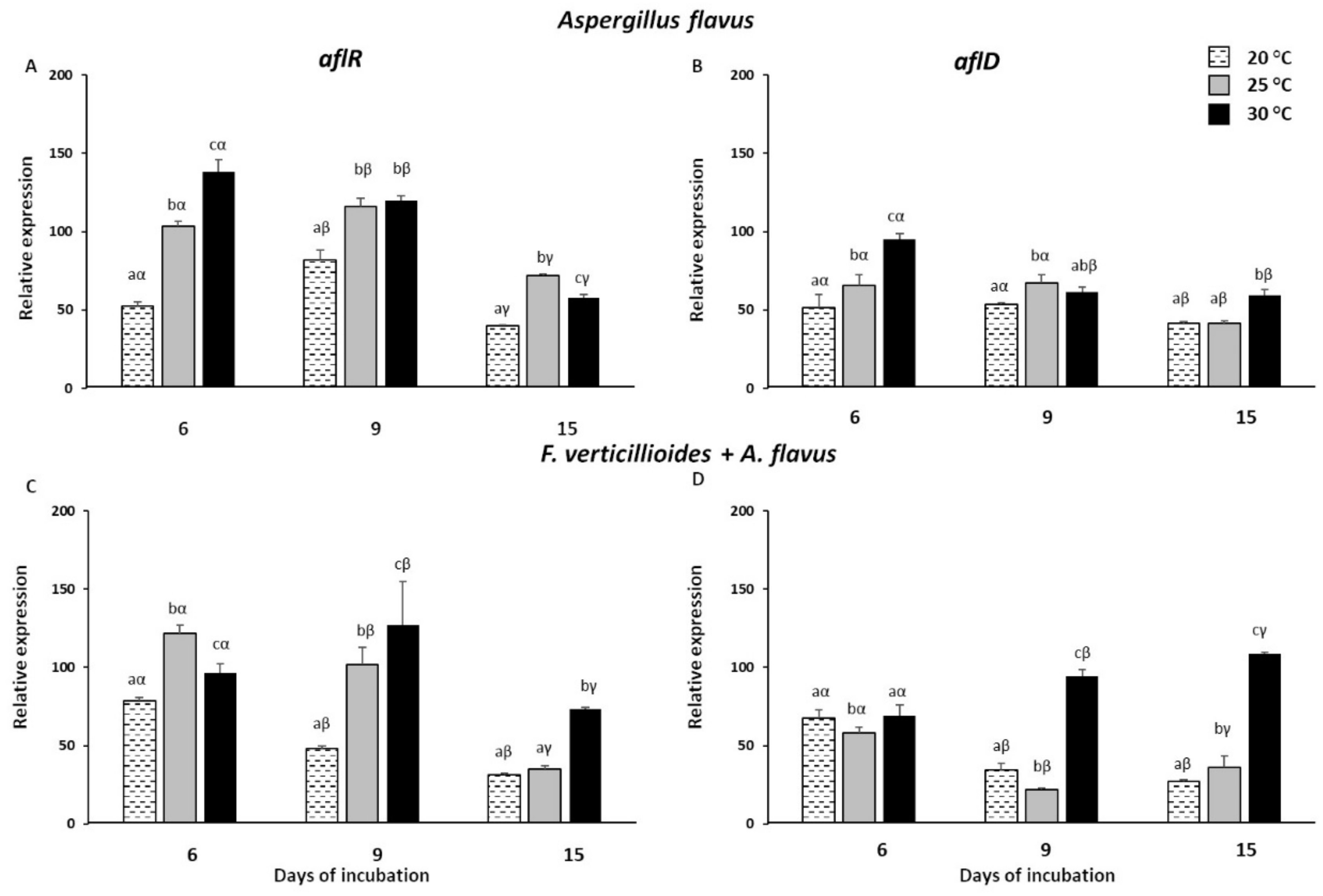
2.2. Expression Analysis of Fusarium verticillioides and Aspergillus flavus Mycotoxin Genes in Infected Kernels
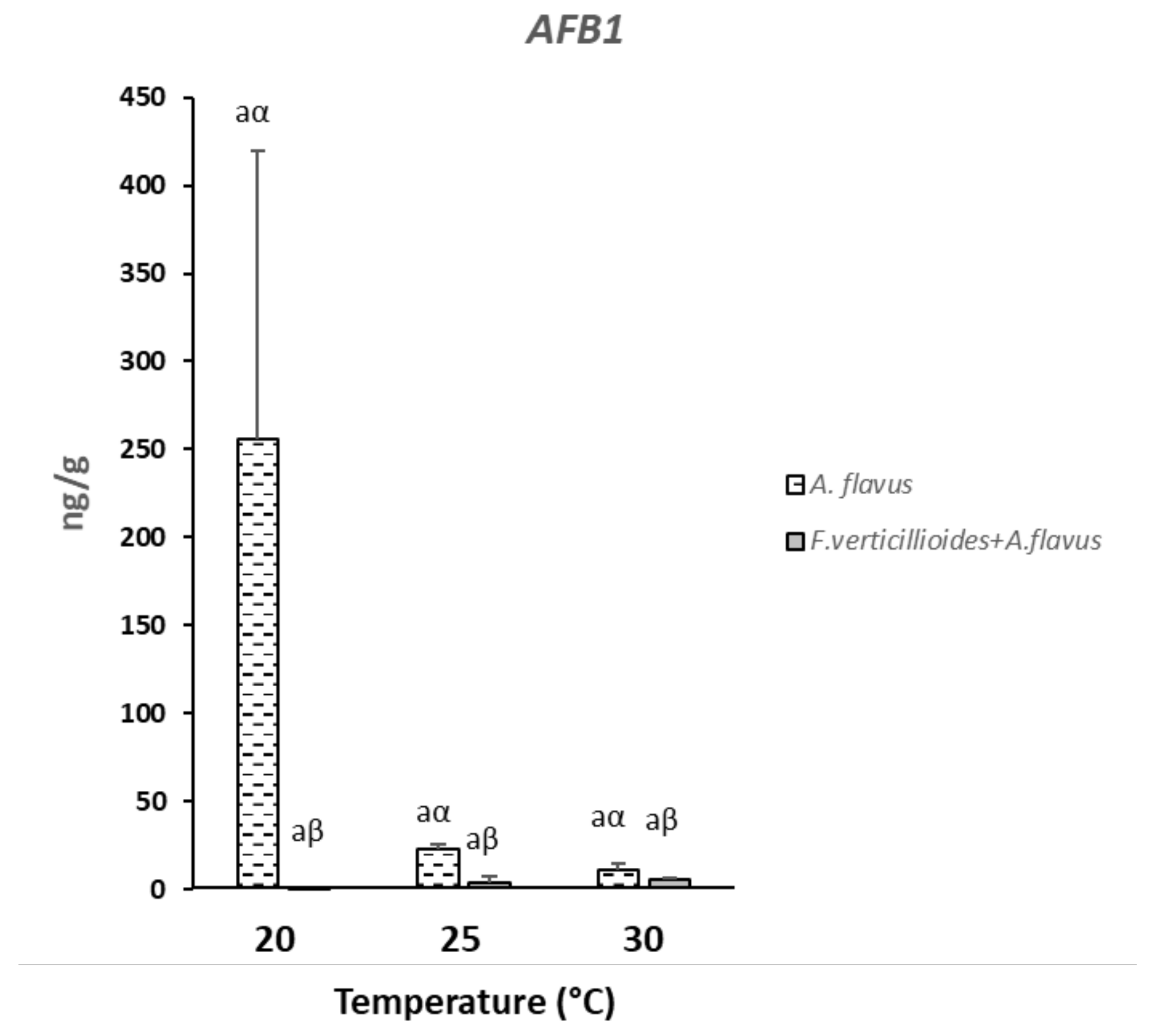
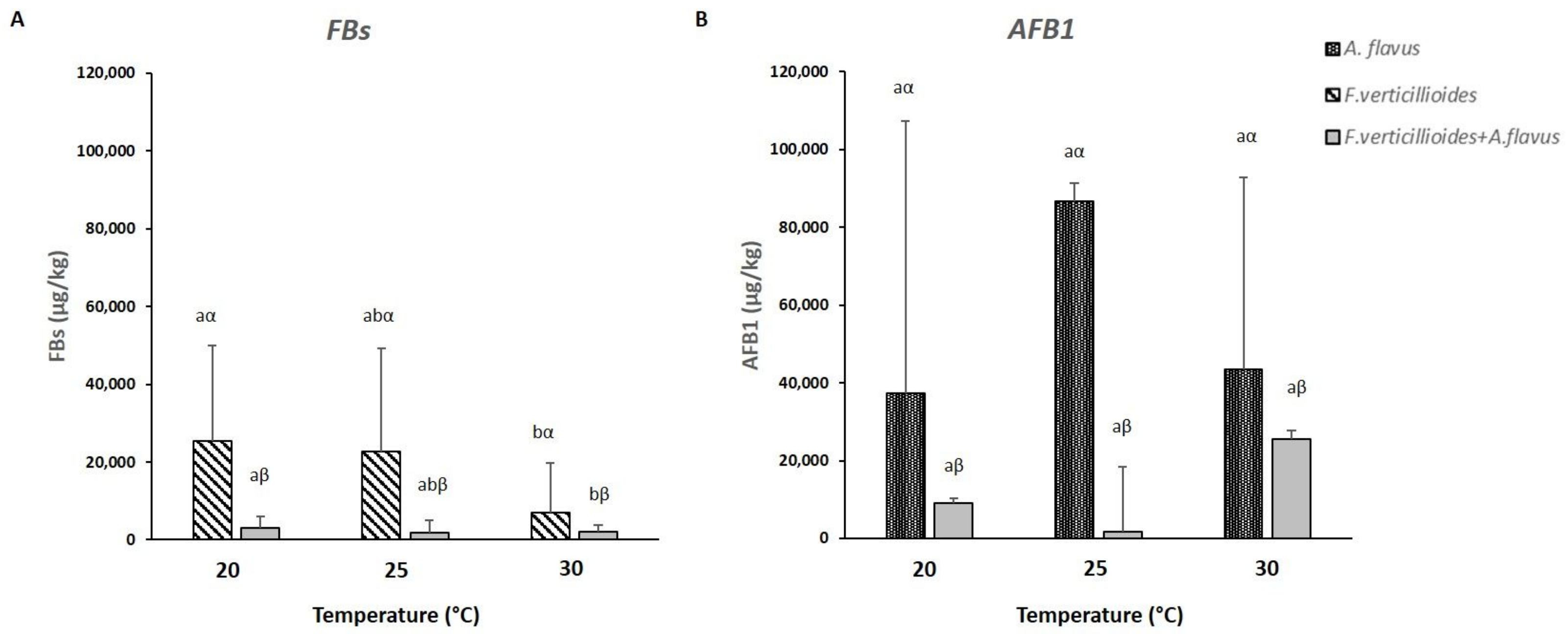
2.3. Mycotoxins Production in Infected Kernels
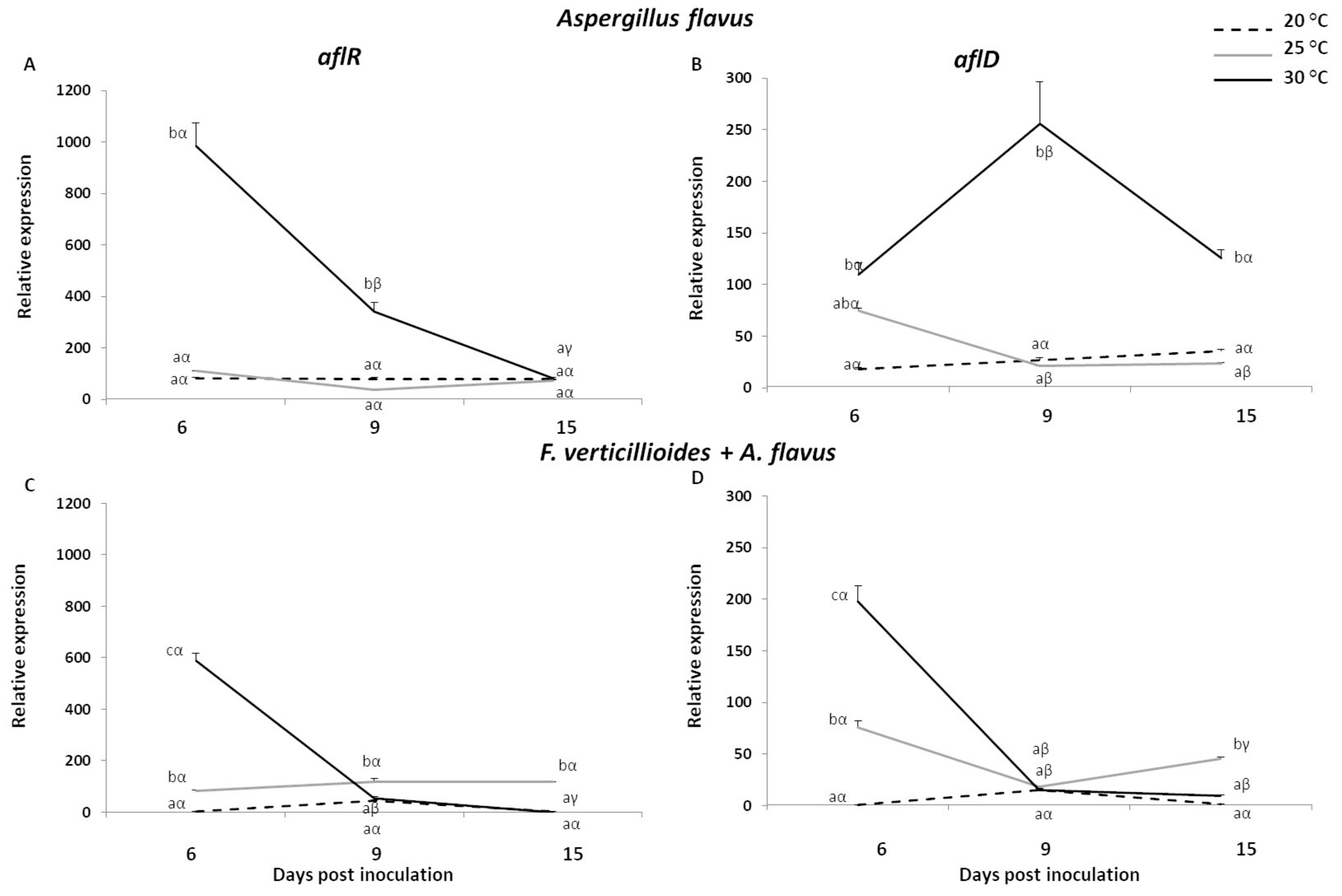
2.4. Expression Analysis of Mycotoxin Genes in Fusarium verticillioides and Aspergillus flavus Grown In Vitro
2.5. Mycotoxins Production by Fusarium verticillioides and Aspergillus flavus Grown In Vitro
3. Discussion
4. Materials and Methods
4.1. Plant Material, Fungal Strains and Inoculation Assay
4.2. RNA Isolation and Real-Time RT-qPCR Expression Analysis
4.3. Mycotoxin Analyses
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Logrieco, A.F.; Moretti, A. Between emerging and historical problems: An overview of the main toxigenic fungi and mycotoxin concerns in Europe. In Mycotoxins: Detection Methods, Management, Public Health and Agricultural Trade; CABI: Wallingford, UK, 2008; pp. 139–153. [Google Scholar]
- Nielsen, L.K.; Jensen, J.D.; Nielsen, G.C.; Jensen, J.E.; Spliid, N.H.; Thomsen, I.K.; Justesen, A.F.; Collinge, D.B.; Jorgensen, L.N. Fusarium head blight of cereals in Denmark: Species complex and related mycotoxins. Phytopathology 2011, 101, 960–969. [Google Scholar] [CrossRef] [Green Version]
- Van der Lee, T.; Zhang, H.; van Diepeningen, A.; Waalwijk, C. Biogeography of Fusarium graminearum species complex and chemotypes: A review. Food Addit. Contam. Part A 2015, 32, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Valverde-Bogantes, E.; Bianchini, A.; Herr, J.R.; Rose, D.J.; Wegulo, S.N.; Hallen-Adams, H.E. Recent population changes of Fusarium head blight pathogens: Drivers and implications. Can. J. Plant Pathol. 2020, 42, 315–329. [Google Scholar] [CrossRef]
- Piva, G.; Battilani, P.; Pietri, A. Emerging issues in Southern Europe: Aflatoxins in Italy. In The Mycotoxin Factbook. Food and Feed Topics; Barug, D., Bhatnagar, D., van Egmond, H.P., van der Kamp, J.W., van Osenbruggen, W.A., Visconti, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; pp. 139–153. [Google Scholar]
- Battilani, P.; Toscano, P.; Van der Fels-Klerx, H.; Moretti, A.; Leggieri, M.C.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [Green Version]
- Dobolyi, C.; Sebok, F.; Varga, J.; Kocsube, S.; Szigeti, G.; Baranyi, N.; Szecsi, A.; Toth, B.; Varga, M.; Kriszt, B.; et al. Occurrence of aflatoxin producing Aspergillus flavus isolates in maize kernel in Hungary. Acta Aliment. 2013, 42, 451–459. [Google Scholar] [CrossRef]
- Levic, J.; Gosic-Dondo, S.; Ivanovic, D.; Stankovic, S.; Krnjaja, V.; Bocarov-Stancic, A.; Stepanic, A. An outbreak of Aspergillus species in response to environmental conditions in Serbia. Pestic. Fitomed. 2013, 28, 167–179. [Google Scholar] [CrossRef]
- Leggieri, M.C.; Toscano, P.; Battilani, P. Predicted aflatoxin b1 increase in europe due to climate change: Actions and reactions at global level. Toxins 2021, 13, 292. [Google Scholar] [CrossRef] [PubMed]
- Krnjaja, V.; Levic, J.; Stankovic, S.; Petrovic, T.; Tomic, Z.; Mandic, V.; Bijelic, Z. Moulds and mycotoxins in stored maize grains. Biotech. Anim. Husb. 2013, 29, 527–536. [Google Scholar] [CrossRef]
- Smith, M.C.; Madec, S.; Coton, E.; Hymery, N. Natural co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, L.; Liu, M.; Su, Y.T.; Xie, W.M.; Zhang, N.Y.; Dai, J.F.; Wang, Y.; Rajput, S.A.; Qi, D.S.; et al. Individual and combined occurrence of mycotoxins in feed ingredients and complete feeds in China. Toxins 2018, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Palumbo, R.; Crisci, A.; Venâncio, A.; Abrahantes, J.C.; Dorne, J.-L.; Battilani, P.; Toscano, P. Occurrence and co-occurrence of mycotoxins in cereal-based feed and food. Microorganisms 2020, 8, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zhang, L.; Xu, Z.; Liu, X.; Chen, L.; Dai, J.; Karrow, N.A.; Sunet, L. Occurrence of Aflatoxin B1, deoxynivalenol and zearalenone in feeds in China during 2018–2020. J. Anim. Sci. Biotechnol. 2021, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Leggieri, M.C.; Lanubile, A.; Dall’Asta, C.; Pietri, A.; Battilani, P. The impact of seasonal weather variation on mycotoxins: Maize crop in 2014 in northern Italy as a case study. World Mycotoxin J. 2020, 13, 25–36. [Google Scholar] [CrossRef]
- Magan, N.; Medina, A. Integrating gene expression, ecology and mycotoxin production by Fusarium and Aspergillus species in relation to interacting environmental factors. World Mycotoxin J. 2016, 9, 863–874. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Landschoot, S.; Detavernier, C.; De Saeger, S.; Rajkovic, A.; Audenaert, K. Cross-talk between Fusarium verticillioides and Aspergillus flavus in vitro and in planta. Mycotoxin Res. 2021, 37, 229–240. [Google Scholar] [CrossRef]
- Abdel-Hadi, A.; Schmidt-Heydt, M.; Parra, R.; Geisen, R.; Magan, N. A systems approach to model the relationship between aflatoxin gene cluster expression, environmental factors, growth and toxin production by Aspergillus flavus. J. R. Soc. Interface 2012, 9, 757–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, A.; Schmidt-Heydt, M.; Cárdenas-Chávez, D.L.; Parra, R.; Geisen, R.; Magan, N. Integrating toxin gene expression, growth and fumonisin B1 and B2 production by a strain of Fusarium verticillioides under different environmental factors. J. R. Soc. Interface 2013, 10, 20130320. [Google Scholar] [CrossRef] [Green Version]
- Giorni, P.; Bertuzzi, T.; Battilani, P. Impact of fungi co-occurrence on mycotoxin contamination in maize during the growing season. Front. Microbiol. 2019, 10, 1265. [Google Scholar] [CrossRef]
- Zummo, N.; Scott, G.E. Interaction of Fusarium verticillioides and Aspergillus flavus on kernel infection and aflatoxin contamination in maize ears. Plant Dis. 1992, 76, 771–773. [Google Scholar] [CrossRef]
- Leggieri, M.C.; Giorni, P.; Pietri, A.; Battilani, P. Aspergillus flavus and Fusarium verticillioides interaction: Modeling the impact on mycotoxin production. Front. Microbiol. 2019, 10, 2653. [Google Scholar]
- Ferreira-Castro, F.L.; Potenza, M.R.; Rocha, L.O.; Correa, B. Interaction between toxigenic fungi and weevils in corn grain samples. Food Control 2012, 26, 594–600. [Google Scholar] [CrossRef]
- Majumdar, R.; Rajasekaran, K.; Sickler, C.; Lebar, M.; Musungu, B.M.; Fakhoury, A.M.; Payne, G.A.; Geisler, M.; Carter-Wientjes, C.; Wei, Q.; et al. The pathogenesis-related maize seed (PRms) gene plays a role in resistance to Aspergillus flavus infection and aflatoxin contamination. Front. Plant Sci. 2017, 8, 1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maschietto, V.; Lanubile, A.; De Leonardis, S.; Marocco, A.; Paciolla, C. Constitutive expression of pathogenesis-related proteins and antioxydant enzyme activities triggers maize resistance towards Fusarium verticillioides. J. Plant Physiol. 2016, 200, 53–61. [Google Scholar] [CrossRef]
- Jurado, M.; Marin, P.; Magan, N.; Gonzalez-Jaen, M.T. Relationship between solute and matric potential stress, temperature, growth, and FUM1 gene expression in two Fusarium verticillioides strains from Spain. Appl. Environ. Microbiol. 2008, 74, 2032–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, S.A.; Borrego, E.; Shim, W.; Isakeit, T.; Kolomiets, M. Quantification of fungal colonization, sporogenesis, and production of mycotoxins using kernel bioassays. J. Vis. Exp. 2012, 23, 3727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battilani, P.; Lanubile, A.; Scala, V.; Reverberi, M.; Gregori, R.; Falavigna, C.; Dall’asta, C.; Park, Y.S.; Bennett, J.; Borrego, E.J.; et al. Oxylipins from both pathogen and host antagonize jasmonic acid-mediated defence via the 9-lipoxygenase pathway in Fusarium verticillioides infection of maize. Mol. Plant Pathol. 2018, 19, 2162–2176. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.H.; Plattner, R.D.; Desjardins, A.E.; Busman, M.; Butchko, R.A.E. Fumonisin production in the maize pathogen Fusarium verticillioides: Genetic basis of naturally occurring chemical variation. J. Agric. Food Chem. 2006, 54, 2424–2430. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, A.E.; Proctor, R.H. Molecular biology of Fusarium mycotoxins. Int. J. Food Microbiol. 2007, 119, 47–50. [Google Scholar] [CrossRef]
- Caceres, I.; Al Khoury, A.; El Khoury, R.; Lorber, S.; Oswald, I.P.; El Khoury, A.; Atoui, A.; Puel, O.; Bailly, J.-D. Aflatoxin biosynthesis and genetic regulation: A review. Toxins 2020, 12, 150. [Google Scholar] [CrossRef] [Green Version]
- Juroszek, P.; von Tiedemann, A. Climatic changes and the potential future importance of maize diseases: A short review. J. Plant Dis. Prot. 2013, 120, 49–56. [Google Scholar] [CrossRef]
- Medina, A.; Akbar, A.; Baazeem, A.; Rodriguez, A.; Magan, N. Climate change, food security and mycotoxins: Do we know enough? Fungal Biol. Rev. 2017, 31, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Logrieco, A.; Battilani, P.; Leggieri, M.C.; Jiang, Y.; Haesaert, G.; Lanubile, A.; Mahuku, G.; Mesterhazy, A.; Ortega-Beltran, A.; Pasti, M.; et al. Perspectives on global mycotoxin issues and management from the MycoKey Maize Working Group. Plant Dis. 2021, 105, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Vandicke, J.; De Visschere, K.; Croubels, S.; De Saeger, S.; Audenaert, K.; Haesaert, G. Mycotoxins in Flanders’ fields: Occurrence and correlations with Fusarium species in whole-plant harvested maize. Microorganisms 2019, 7, 571. [Google Scholar] [CrossRef] [Green Version]
- Lanubile, A.; Ferrarini, A.; Maschietto, V.; Delledonne, M.; Marocco, A.; Bellin, D. Functional genomic analysis of constitutive and inducible defence responses to Fusarium verticillioides infection in maize genotypes with contrasting ear rot resistance. BMC Genom. 2014, 15, 710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanubile, A.; Maschietto, V.; Borrelli, V.M.; Stagnati, L.; Logrieco, A.F.; Marocco, A. Molecular basis of resistance to Fusarium ear rot in maize. Front. Plant Sci. 2017, 8, 1774. [Google Scholar] [CrossRef] [PubMed]
- Lanubile, A.; Borrelli, V.M.; Soccio, M.; Giorni, P.; Stagnati, L.; Busconi, M.; Marocco, A. Loss of ZmLIPOXYGENASE4 decreases Fusarium verticillioides resistance in maize seedlings. Genes 2021, 12, 335. [Google Scholar] [CrossRef] [PubMed]
- Lanubile, A.; Maschietto, V.; De Leonardis, S.; Battilani, P.; Paciolla, C.; Marocco, A. Defence responses to mycotoxin-producing fungi Fusarium proliferatum, F. subglutinans, and Aspergillus flavus in kernels of susceptible and resistant maize genotypes. Mol. Plant Microbe Interact. 2015, 28, 546–557. [Google Scholar] [CrossRef] [Green Version]
- Lanubile, A.; Maschietto, V.; Battilani, P.; Marocco, A. Infection with toxigenic and atoxigenic strains of Aspergillus flavus induces different transcriptional signatures in maize kernels. J. Plant Interact. 2017, 12, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Shu, X.; Livingston, D.P.; Franks, R.G.; Boston, R.S.; Woloshuk, C.P.; Payne, G.A. Tissue-specific gene expression in maize seeds during colonization by Aspergillus flavus and Fusarium verticillioides. Mol. Plant Pathol. 2015, 16, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Marín, S.; Sanchis, V.; Ramos, A. Environmental factors, in vitro interactions, and niche overlap between Fusarium moniliforme, F. proliferatum, and F. graminearum, Aspergillus and Penicillium species from maize. Mycol. Res. 1998, 102, 831–837. [Google Scholar] [CrossRef]
- Lazzaro, I.; Susca, A.; Mulè, G.; Ritieni, A.; Ferracane, R.; Marocco, A.; Battilani, P. Effects of temperature and water activity on FUM2 and FUM21 gene expression and fumonisin B production in Fusarium verticillioides. Eur. J. Plant Pathol. 2012, 134, 685–695. [Google Scholar] [CrossRef]
- Rocha, L.O.; Reis, G.M.; Fontes, L.C.; Piacentini, K.C.; Barroso, V.M.; Reis, T.A.; Pereira, A.A.; Corrêa, B. Association between FUM expression and fumonisin contamination in maize from silking to harvest. Crop Prot. 2017, 94, 77–82. [Google Scholar] [CrossRef]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered pathway genes in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Hua, S.S.; Beck, J.J.; Sarreal, S.B.L.; Gee, W. The major volatile compound 2-phenylethanol from the biocontrol yeast, Pichia anomala, inhibits growth and expression of aflatoxin biosynthetic genes of Aspergillus flavus. Mycotoxin Res. 2014, 30, 71–78. [Google Scholar] [CrossRef]
- Dooso Oloo, R.; Okoth, S.; Wachira, P.; Mutiga, S.; Ochieng, P.; Kago, L.; Nganga, F.; Domelevo Entfellner, J.B.; Ghimire, S. Genetic profiling of Aspergillus isolates with varying aflatoxin production potential from different maize-growing regions of Kenya. Toxins 2019, 11, 467. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Fedorova, N.D.; Montalbano, B.G.; Bhatnagar, D.; Cleveland, T.E.; Bennett, J.W.; Nierman, W.C. Tight control of mycotoxin biosynthesis gene expression in Aspergillus flavus by temperature as revealed by RNA-Seq. FEMS Microbiol. Lett. 2011, 322, 145–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, A.; Solfrizzo, M.; Epifani, F.; Panzarini, G.; Perrone, G. Effect of temperature and water activity on gene expression and aflatoxin biosynthesis in Aspergillus flavus on almond medium. Int. J. Food Microbiol. 2016, 217, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Zorzete, P.; Castro, R.S.; Pozzi, C.R.; Israel, A.L.M.; Fonseca, H.; Yanaguibashi, G.; Corrêa, B. Relative populations and toxin production by Aspergillus flavus and Fusarium verticillioides in artificially inoculated corn at various stages of development under field conditions. J. Sci. Food Agric. 2008, 88, 48–55. [Google Scholar] [CrossRef]
- Picot, A.; Hourcade-Marcolla, D.; Barreau, C.; Pinson-Gadais, L.; Caron, D.; Richard-Forget, F.; Lannou, C. Interactions between Fusarium verticillioides and Fusarium graminearum in maize ears and consequences for fungal development and mycotoxin accumulation. Plant Pathol. 2012, 61, 140–151. [Google Scholar] [CrossRef]
- Giorni, P.; Bertuzzi, T.; Battilani, P. Aflatoxin in maize, a multifaceted answer of Aspergillus flavus governed by weather, host-plant and competitor fungi. J. Cereal Sci. 2016, 70, 256–262. [Google Scholar] [CrossRef]
- Garcia-Cela, E.; Kiaitsi, E.; Sulyok, M.; Krska, R.; Medina, A.; Damico, I.P.; Magan, N. Influence of storage environment on maize grain: CO2 production, dry matter losses and aflatoxins contamination. Food Addit. Contam. Part A 2019, 36, 175–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, M.K.; Medina, A.; Mack, B.M.; Lebar, M.; Rodriguez, A.; Bhatnagar, D.; Magan, N.; Obrian, G.; Payne, G. Carbon dioxide mediates the response to temperature and water activity levels in Aspergillus flavus during infection of maize kernels. Toxins 2018, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, A.; Rodriguez, A.; Magan, N. Climate change and mycotoxigenic fungi: Impacts on mycotoxin production. Curr. Opin. Food Sci. 2015, 5, 99–104. [Google Scholar] [CrossRef]
- Baazeem, A.; Rodriguez, A.; Medina, A.; Magan, N. Impacts of climate change interacting abiotic factors on growth, aflD and aflR gene expression and aflatoxin B1 production by Aspergillus flavus strains in vitro and on pistachio nuts. Toxins 2021, 13, 385. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Errasquín, E.; Vazquez, C.; Jimenez, M.; Gonzalez-Jaen, M.T. Real-Time RT-PCR assay to quantify the expression of FUM1 and FUM19 genes from the fumonisin-producing Fusarium verticillioides. J. Microbiol. Methods 2007, 68, 312–317. [Google Scholar] [CrossRef]
- Van Zyl, K.; Rose, L.J.; Viljoen, A. Fusarium verticillioides FUM1 and FUM19 gene expression in maize kernels during early infection. Physiol. Mol. Plant Pathol. 2019, 108, 101430. [Google Scholar] [CrossRef]
- Gao, X.; Brodhagen, M.; Isakeit, T.; Brown, S.H.; Göbel, C.; Betran, J.; Feussner, I.; Keller, N.P.; Kolomiets, M.V. Inactivation of the lipoxygenase ZmLOX3 increases susceptibility of maize to Aspergillus spp. Mol. Plant Microbe Interact. 2009, 22, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Lanubile, A.; Logrieco, A.; Battilani, P.; Proctor, R.H.; Marocco, A. Transcriptional changes in developing maize kernels in response to fumonisin-producing and nonproducing strains of Fusarium verticillioides. Plant Sci. 2013, 210, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.O.; Barroso, V.M.; Andrade, L.J.; Pereira, G.H.A.; Ferreira-Castro, F.L.; Duarte, A.P.; Michelotto, M.D.; Correa, B. FUM gene expression profile and fumonisin production by Fusarium verticillioides inoculated in Bt and non-Bt maize. Front. Microbiol. 2016, 6, 150. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, T.; Rastelli, S.; Mulazzi, A.; Pietri, A. Evaluation and improvement of extraction methods for the analysis of aflatoxins B1, B2, G1 and G2 from naturally contaminated maize. Food Anal. Methods 2012, 5, 512–519. [Google Scholar] [CrossRef]
- Pietri, A.; Bertuzzi, T. Simple phosphate buffer extraction for the determination of fumonisins in masa, maize and derived products. Food Anal. Methods 2012, 5, 1088–1096. [Google Scholar] [CrossRef]
- Clewer, A.G.; Scarisbrick, D.H. Practical Statistics and Experimental Design for Plant and Crop Science; John Wiley & Sons, Ltd.: Chichester, UK, 2001. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Forward (5′ → 3′) | Primer Reverse (5′ → 3′) | Source |
|---|---|---|---|
| β-actin | ATGGTCAAGGCCGGTTTCG | TCAGGATGCCTCTCTTGGCC | AY273142.1 [61] |
| PR5 | GTCATCGACGGCTACAACCT | GGGCAGAAGGTGACTTGGTA | U82201.1 [39] |
| PRm3 | GGCTCTACGCCTACGTCAAC | GATGGAGAGGAGCACCTTGA | S82314.1 [39] |
| FvCalmodulin | GATGGCCAGATTACCACCAA | CGCCATCATGGTAAGGAACT | HQ412321.1 [25] |
| FUM1 | GAGCCGAGTCAGCAAGGATT | AGGGTTCGTGAGCCAAGGA | AF155773 [57] |
| FUM13 | GCCTTTGGTCTTGTTCTCTCA | CGTCAATTATTGCCTCTTTCAA | AF155773 [62] |
| AfCalmodulin | GGCCGACTCTTTGACTGAAG | CATGTCCTGGAGTTCCGACT | XM_002374071.1 [39] |
| aflR | GAGCAAAGCACCCTGTCTTC | GTCGACTCGGCCAAGAAATC | XM_002379905 [46] |
| aflD | GCGCAAGTTCCACTTTGAGA | CCTTGGTCGCCCATATCAGT | AFLA_139390 [49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanubile, A.; Giorni, P.; Bertuzzi, T.; Marocco, A.; Battilani, P. Fusarium verticillioides and Aspergillus flavus Co-Occurrence Influences Plant and Fungal Transcriptional Profiles in Maize Kernels and In Vitro. Toxins 2021, 13, 680. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100680
Lanubile A, Giorni P, Bertuzzi T, Marocco A, Battilani P. Fusarium verticillioides and Aspergillus flavus Co-Occurrence Influences Plant and Fungal Transcriptional Profiles in Maize Kernels and In Vitro. Toxins. 2021; 13(10):680. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100680
Chicago/Turabian StyleLanubile, Alessandra, Paola Giorni, Terenzio Bertuzzi, Adriano Marocco, and Paola Battilani. 2021. "Fusarium verticillioides and Aspergillus flavus Co-Occurrence Influences Plant and Fungal Transcriptional Profiles in Maize Kernels and In Vitro" Toxins 13, no. 10: 680. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100680