Peripherally Administered Botulinum Toxin Type A Localizes Bilaterally in Trigeminal Ganglia of Animal Model


 ,
,
Abstract
:1. Introduction
2. Results
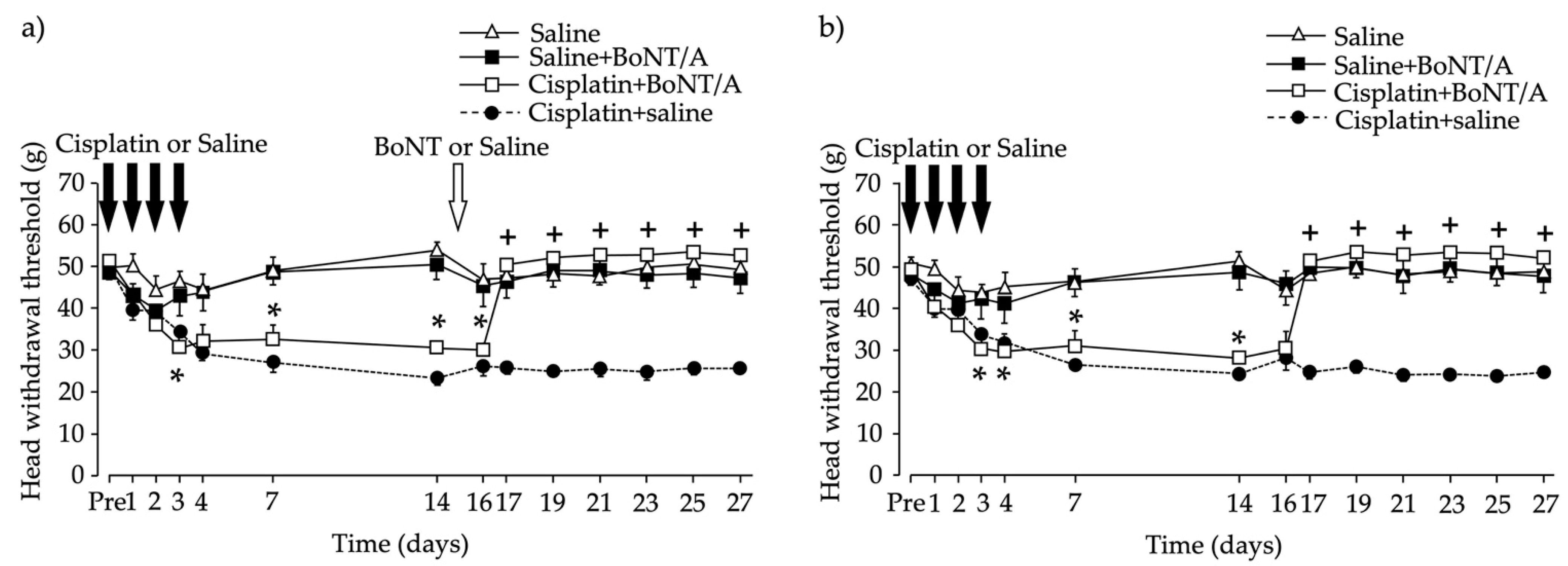
2.1. Head Withdrawal Threshold after Chemotherapy-Induced Bilateral Neuropathy and BoNT/A Application
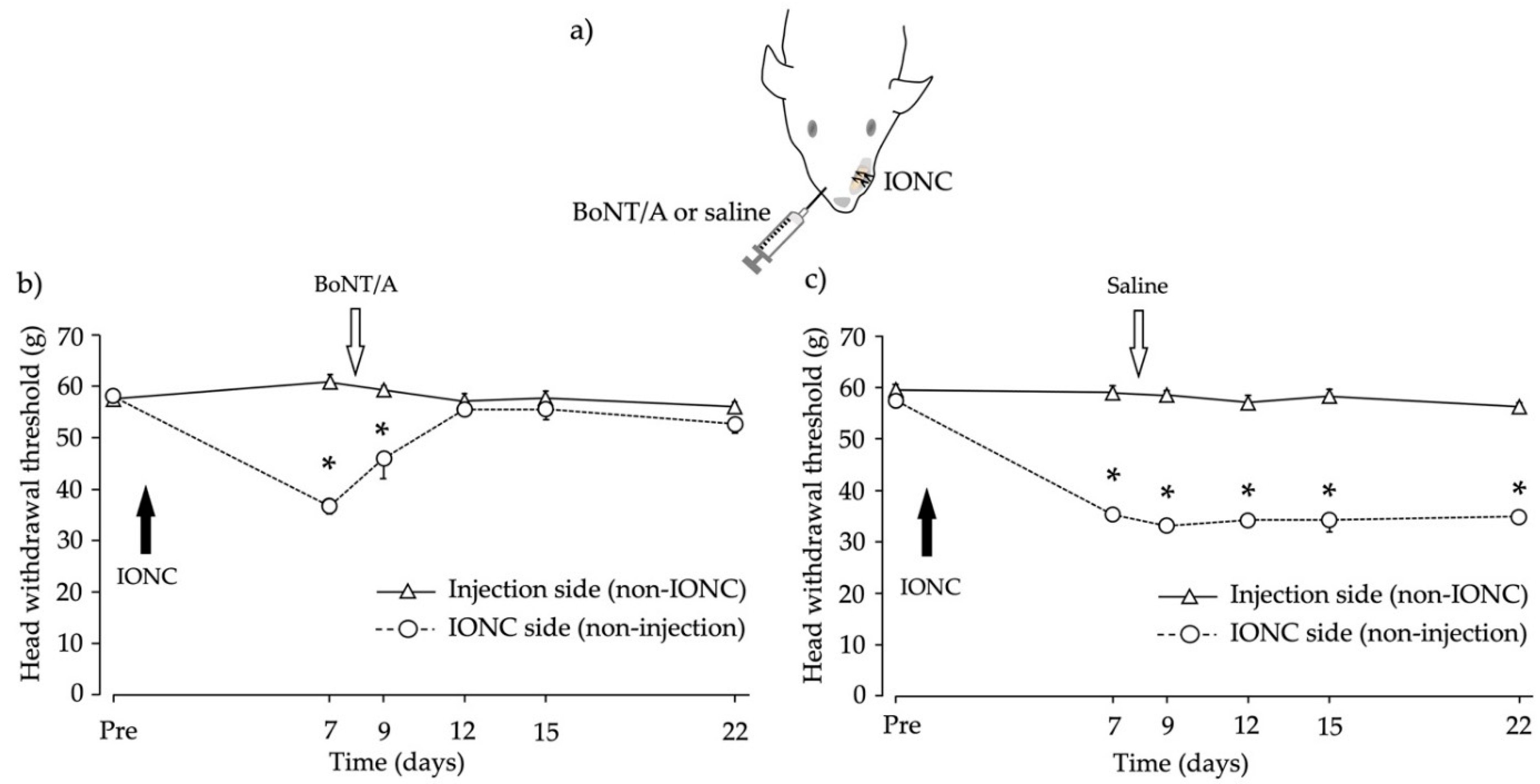
2.2. Head Withdrawal Threshold after Infraorbital Nerve Constriction (IONC) and Contralateral BoNT/A Application
2.3. Mouse Bioassay to Detect BoNT/A Concentrations in the Blood
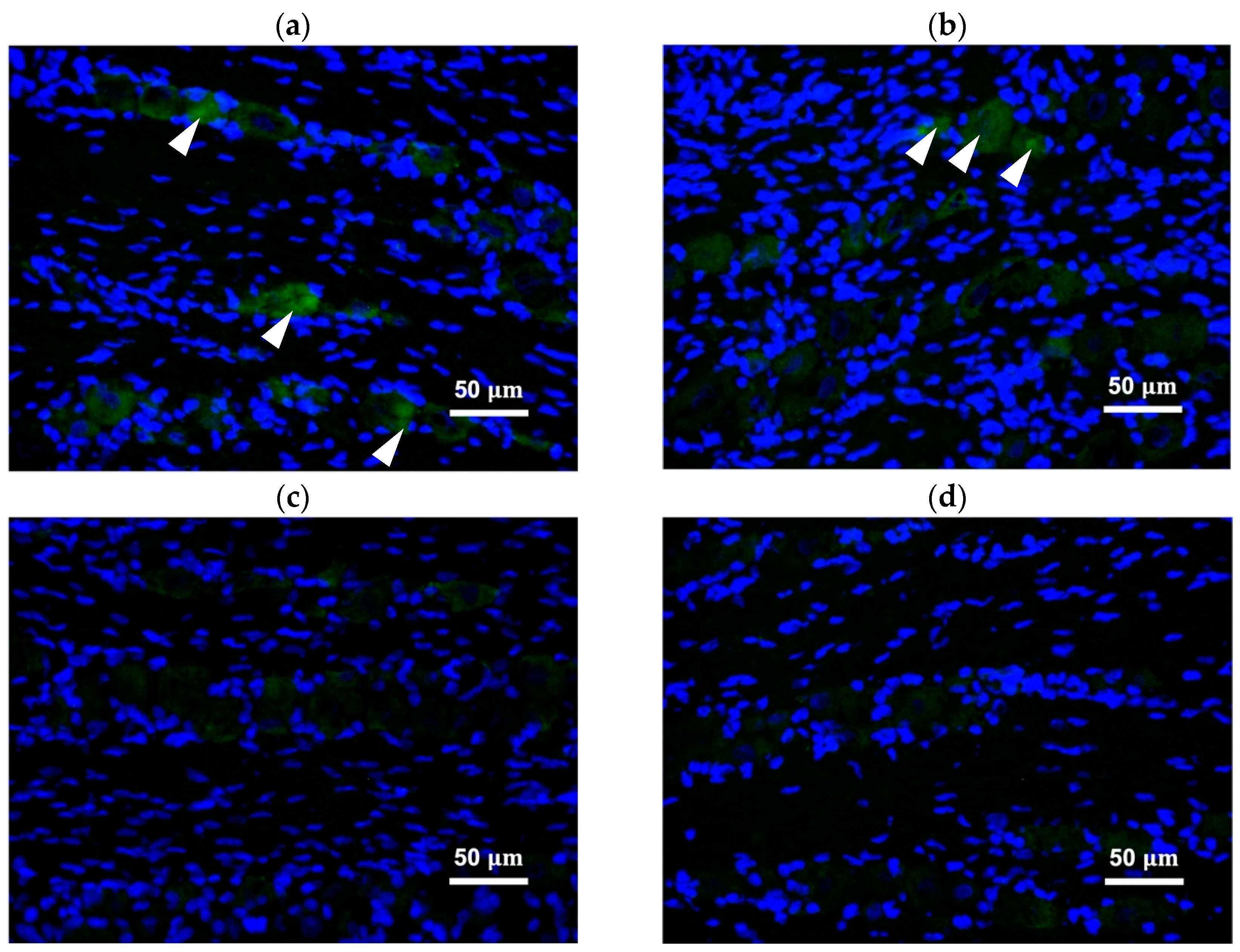
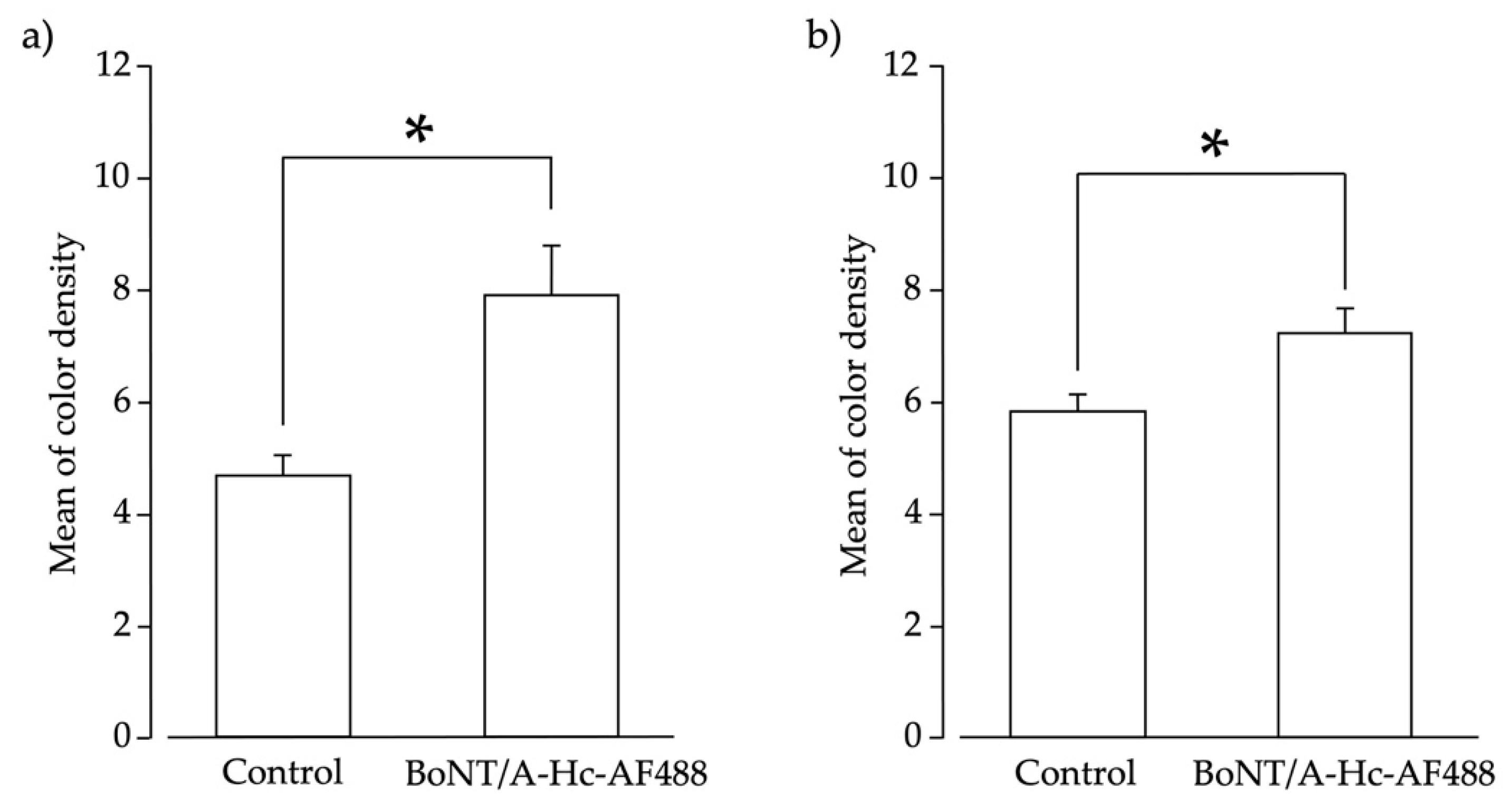
2.4. Peripherally Administered BoNT/A-Hc Localized in the TG
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
5.2. Chemotherapy-Induced Bilateral Neuropathic Pain
5.3. IONC
5.4. Drug Administration
5.5. Behavioral Testing
5.6. Mouse Bioassay
5.7. Immunohistochemistry
5.8. Densitometric Analysis
5.9. Statistical Analysis
Author Contributions
Funding
Institutional Board Review
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shinoda, M.; Kubo, A.; Hayashi, Y.; Iwata, K. Peripheral and Central Mechanisms of Persistent Orofacial Pain. Front. Neurosci. 2019, 13, 1227. [Google Scholar] [CrossRef]
- Ma, C.; Shu, Y.; Zheng, Z.; Chen, Y.; Yao, H.; Greenquist, K.W.; White, F.A.; LaMotte, R.H. Similar Electrophysiological Changes in Axotomized and Neighboring Intact Dorsal Root Ganglion Neurons. J. Neurophysiol. 2003, 89, 1588–1602. [Google Scholar] [CrossRef] [Green Version]
- Neubert, J.K.; Maidment, N.T.; Matsuka, Y.; Adelson, D.W.; Kruger, L.; Spigelman, I. Inflammation induced changes in primary afferent-evoked release of substance P within trigeminal ganglia in vivo. Brain Res. 2000, 871, 181–191. [Google Scholar] [CrossRef]
- Matsuka, Y.; Neubert, J.K.; Maidment, N.T.; Spigelman, I. Concurrent release of ATP and substance P within guinea pig trigeminal ganglia in vivo. Brain Res. 2001, 915, 248–255. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.M.; Flores, C.M.; Harding-Rose, C.A.; Goodis, H.E.; Hargreaves, K.M. Capsaicin-evoked release of immunoreactive calcitonin gene-related peptide from rat trigeminal ganglion: Evidence for intraganglionic neurotransmission. Pain 2001, 91, 219–226. [Google Scholar] [CrossRef]
- Omoto, K.; Maruhama, K.; Terayama, R.; Yamamoto, Y.; Matsushita, O.; Sugimoto, T.; Oguma, K.; Matsuka, Y. Cross-excitation in peripheral sensory ganglia associated with pain transmission. Toxins 2015, 7, 2906–2917. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.; Matsuka, Y.; Spigelman, I.; Ishihara, Y.; Yamamoto, Y.; Sonoyama, W.; Kuboki, T.; Oguma, K. Botulinum toxin type a (150 kDa) decreases exaggerated neurotransmitter release from trigeminal ganglion neurons and relieves neuropathy behaviors induced by infraorbital nerve constriction. Neuroscience 2009, 159, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Kumada, A.; Matsuka, Y.; Spigelman, I.; Maruhama, K.; Yamamoto, Y.; Neubert, J.K.; Nolan, T.A.; Watanabe, K.; Maekawa, K.; Kamioka, H.; et al. Intradermal injection of Botulinum toxin type A alleviates infraorbital nerve constriction-induced thermal hyperalgesia in an operant assay. J. Oral Rehabil. 2012, 39, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Azarnia Tehran, D.; Zanetti, G.; Leka, O.; Lista, F.; Fillo, S.; Binz, T.; Shone, C.C.; Rosseto, O.; Montecucco, C.; Paradisi, C.; et al. A Novel Inhibitor Prevents the Peripheral Neuroparalysis of Botulinum Neurotoxins. Sci. Rep. 2015, 5, 17513. [Google Scholar] [CrossRef] [Green Version]
- Pellett, S.; Yaksh, T.L.; Ramachandran, R. Current Status and Future Directions of Botulinum Neurotoxins for Targeting Pain Processing. Toxins 2015, 7, 4519–4563. [Google Scholar] [CrossRef]
- Matak, I.; Bölcskei, K.; Bach-Rojecky, L.; Helyes, Z. Mechanisms of Botulinum Toxin Type A Action on Pain. Toxins 2019, 11, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welch, M.J.; Purkiss, J.R.; Foster, K.A. Sensitivity of embryonic rat dorsal root ganglia neurons to Clostridium botulinum neurotoxins. Toxicon 2000, 38, 245–258. [Google Scholar] [CrossRef]
- McMahon, H.T.; Foran, P.; Dolly, J.O.; Verhage, M.; Wiegant, V.M.; Nicholls, D.G. Tetanus toxin and botulinum toxins type A and B inhibit glutamate, gamma-aminobutyric acid, aspartate, and met-enkephalin release from synaptosomes. Clues to the locus of action. J. Biol. Chem. 1992, 267, 21338–21343. [Google Scholar] [CrossRef]
- Durham, P.L.; Cady, R.; Cady, R. Regulation of calcitonin gene-related peptide secretion from trigeminal nerve cells by botulinum toxin type A: Implications for migraine therapy. Headache 2004, 44, 35–43. [Google Scholar] [CrossRef]
- Koizumi, H.; Goto, S.; Okita, S.; Morigaki, R.; Akaike, N.; Torii, Y.; Harakawa, T.; Ginnaga, A.; Kaji, R. Spinal Central Effects of Peripherally Applied Botulinum Neurotoxin A in Comparison between Its Subtypes A1 and A2. Front. Neurol. 2014, 5, 98. [Google Scholar] [CrossRef] [Green Version]
- Antonucci, F.; Rossi, C.; Gianfranceschi, L.; Rossetto, O.; Caleo, M. Long-distance retrograde effects of botulinum neurotoxin A. J. Neurosci. 2008, 28, 3689–3696. [Google Scholar] [CrossRef]
- Lee, H.B.; Blaufox, M.D. Blood Volume in the Rat. J. Nucl. Med. 1985, 26, 72–76. [Google Scholar]
- Oh, H.M.; Chung, M.E. Botulinum toxin for neuropathic pain: A review of the literature. Toxins 2015, 7, 3127–3154. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga, C.; Díaz, S.; Piedimonte, F.; Micheli, F. Beneficial effects of botulinum toxin type A in trigeminal neuralgia. Arq. Neuropsiquiatr. 2008, 66, 500–503. [Google Scholar] [CrossRef] [Green Version]
- Ngeow, W.C.; Nair, R. Injection of botulinum toxin type A (BOTOX) into trigger zone of trigeminal neuralgia as a means to control pain. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2010, 109, 47–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.J.; Lian, Y.J.; Zheng, Y.K.; Zhang, H.F.; Chen, Y.; Xie, N.C.; Wang, L.J. Botulinum toxin type A for the treatment of trigeminal neuralgia: Results from a randomized, double-blind, placebo-controlled trial. Cephalalgia 2012, 32, 443–450. [Google Scholar] [CrossRef]
- Li, S.; Lian, Y.J.; Chen, Y.; Zhang, H.F.; Ma, Y.Q.; He, C.H.; Wu, C.J.; Xie, N.C.; Zheng, Y.K.; Zhang, Y. Therapeutic effect of botulinum toxin-a in 88 patients with trigeminal neuralgia with 14-month follow-up. J. Headache Pain 2014, 15, 43. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Khanijou, S.; Rubino, J.; Aoki, K.R. Subcutaneous administration of botulinum toxin A reduces formalin-induced pain. Pain 2004, 107, 125–133. [Google Scholar] [CrossRef]
- Wu, C.; Xie, N.; Lian, Y.; Xu, H.; Chen, C.; Zheng, Y.; Chen, Y.; Zhang, H. Central antinociceptive activity of peripherally applied botulinum toxin type A in lab rat model of trigeminal neuralgia. Springerplus 2016, 5, 431. [Google Scholar] [CrossRef] [Green Version]
- Filipović, B.; Matak, I.; Bach-Rojecky, L.; Lacković, Z. Central action of peripherally applied botulinum toxin type A on pain and dural protein extravasation in rat model of trigeminal neuropathy. PLoS One 2012, 7, e29803. [Google Scholar] [CrossRef] [Green Version]
- Mittal, S.O.; Jabbari, B. Botulinum neurotoxins and cancer—A review of the literature. Toxins 2020, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Bhaskar, A. Chemotherapy-induced peripheral neuropathic pain. BJA Educ. 2016, 16, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Jordan, B.; Margulies, A.; Cardoso, F.; Cavaletti, G.; Haugnes, H.S.; Jahn, P.; Le Rhun, E.; Preusser, M.; Scotté, F.; Taphoorn, M.J.; et al. Systemic anticancer therapy-induced peripheral and central neurotoxicity: ESMO–EONS–EANO Clinical Practice Guidelines for diagnosis, prevention, treatment, and follow-up. Ann. Oncol. 2020, 31, 1306–1319. [Google Scholar] [CrossRef]
- Favre-Guilmard, C.; Auguet, M.; Chabrier, P.-E. Different antinociceptive effects of botulinum toxin type A in inflammatory and peripheral polyneuropathic rat models. Eur. J. Pharmacol. 2009, 617, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Cobianchi, S.; Jaramillo, J.; Luvisetto, S.; Pavone, F.; Navarro, X. Botulinum neurotoxin A promotes functional recovery after peripheral nerve injury by increasing regeneration of myelinated fibers. Neuroscience 2017, 359, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, S.; Luvisetto, S.; Cobianchi, S.; Makuch, W.; Obara, I.; Mezzaroma, E.; Caruso, M.; Straface, E.; Przewlocka, B.; Pavone, F. Botulinum neurotoxin type A counteracts neuropathic pain and facilitates functional recovery after peripheral nerve injury in animal models. Neuroscience 2010, 171, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Bach-Rojecky, L.; Šalković-Petrišić, M.; Lacković, Z. Botulinum toxin type A reduces pain supersensitivity in experimental diabetic neuropathy: Bilateral effect after unilateral injection. Eur. J. Pharmacol. 2010, 633, 10–14. [Google Scholar] [CrossRef] [Green Version]
- Torii, Y.; Akaike, N.; Harakawa, T.; Kato, K.; Sugimoto, N.; Goto, Y.; Nakahira, S.; Kohda, T.; Kozaki, S.; Kaji, R.; et al. Type A1 but not type A2 botulinum toxin decreases the grip strength of the contralateral foreleg through axonal transport from the toxin-treated foreleg of rats. J. Pharmacol. Sci. 2011, 117, 275–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. Botulism in the United States, 1899–1996, Handbook for Epidemiologists, Clinicians and Laboratory Workers; The Centers: Atlanta, GA, USA, 1998; pp. 15–21. Available online: https://www.cdc.gov/botulism/pdf/bot-manual.pdf (accessed on 20 July 2020).
- Sakamoto, T.; Torii, Y.; Takahashi, M.; Ishida, S.; Goto, Y.; Nakano, H.; Harakawa, T.; Ginnaga, A.; Kozaki, S.; Kaji, R. Quantitative determination of the biological activity of botulinum toxin type A by measuring the compound muscle action potential (CMAP) in rats. Toxicon 2009, 54, 857–861. [Google Scholar] [CrossRef]
- Iwasa, T.; Afroz, S.; Inoue, M.; Arakaki, R.; Oshima, M.; Raju, R.; Waskitho, A.; Inoue, M.; Baba, O.; Matsuka, Y. IL-10 and CXCL2 in trigeminal ganglia in neuropathic pain. Neurosci. Lett. 2019, 703, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Imamura, Y.; Kawamoto, H.; Nakanishi, O. Characterization of heat-hyperalgesia in an experimental trigeminal neuropathy in rats. Exp. Brain Res. 1997, 116, 97–103. [Google Scholar] [CrossRef]
- Lee, J.-C.; Yokoyama, T.; Hwang, H.-J.; Arimitsu, H.; Yamamoto, Y.; Kawasaki, M.; Takigawa, T.; Takeshi, K.; Nishikawa, A.; Kumon, H.; et al. Clinical application of Clostridium botulinum type A neurotoxin purified by a simple procedure for patients with urinary incontinence caused by refractory destrusor overactivity. FEMS Immunol. Med. Microbiol. 2007, 51, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Beeton, C.; Garcia, A.; Chandy, K.G. Drawing Blood from Rats through the Saphenous Vein and by Cardiac Puncture. J. Vis. Exp. 2007, 7, e266. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BoNT/A Injection | Dilution | ||||||
|---|---|---|---|---|---|---|---|
| 2× | 5× | 10× | 20× | 50× | 100× | ||
| A | Peripheral injection | ×× | ×× | ×△× | ×× | ○△ | ○○ |
| B | BoNT/A concentration/0.5 mL = 20 MLD BoNT/A concentration/mL = 20 MLD X 2 = 40 MLD | ||||||
| C | Blood volume = 0.06 × body weight + 0.77 = 0.06 × 360 + 0.77 = 22.37 mL Total BoNT/A concentration in blood = 40 MLD X 22.37 mL = 894.8 MLD | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waskitho, A.; Yamamoto, Y.; Raman, S.; Kano, F.; Yan, H.; Raju, R.; Afroz, S.; Morita, T.; Ikutame, D.; Okura, K.; et al. Peripherally Administered Botulinum Toxin Type A Localizes Bilaterally in Trigeminal Ganglia of Animal Model. Toxins 2021, 13, 704. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100704
Waskitho A, Yamamoto Y, Raman S, Kano F, Yan H, Raju R, Afroz S, Morita T, Ikutame D, Okura K, et al. Peripherally Administered Botulinum Toxin Type A Localizes Bilaterally in Trigeminal Ganglia of Animal Model. Toxins. 2021; 13(10):704. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100704
Chicago/Turabian StyleWaskitho, Arief, Yumiko Yamamoto, Swarnalakshmi Raman, Fumiya Kano, Huijiao Yan, Resmi Raju, Shaista Afroz, Tsuyoshi Morita, Daisuke Ikutame, Kazuo Okura, and et al. 2021. "Peripherally Administered Botulinum Toxin Type A Localizes Bilaterally in Trigeminal Ganglia of Animal Model" Toxins 13, no. 10: 704. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100704