Exploration on the Enhancement of Detoxification Ability of Zearalenone and Its Degradation Products of Aspergillus niger FS10 under Directional Stress of Zearalenone

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result and Discussion
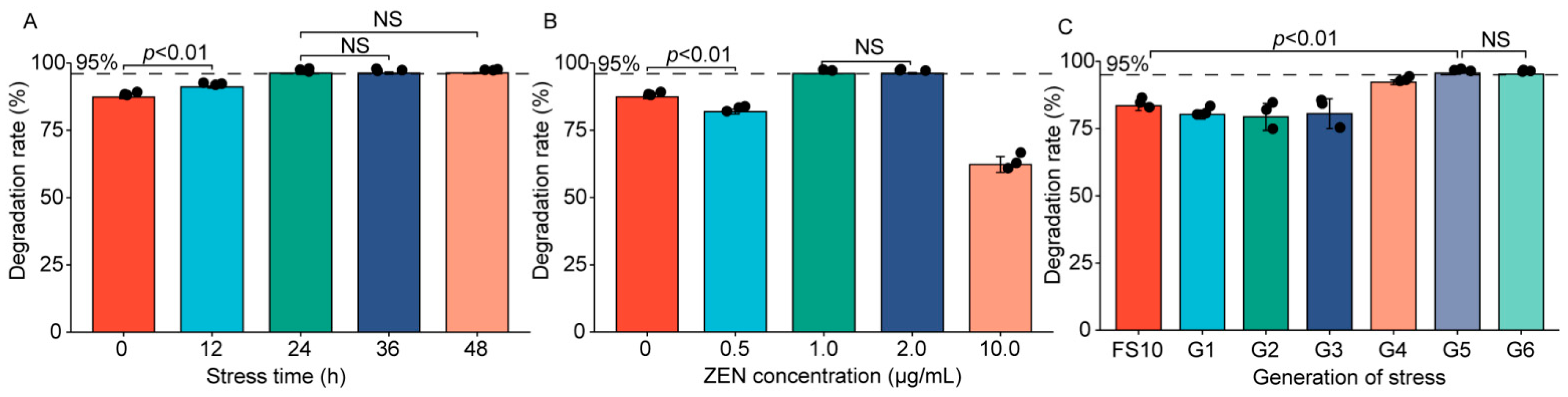
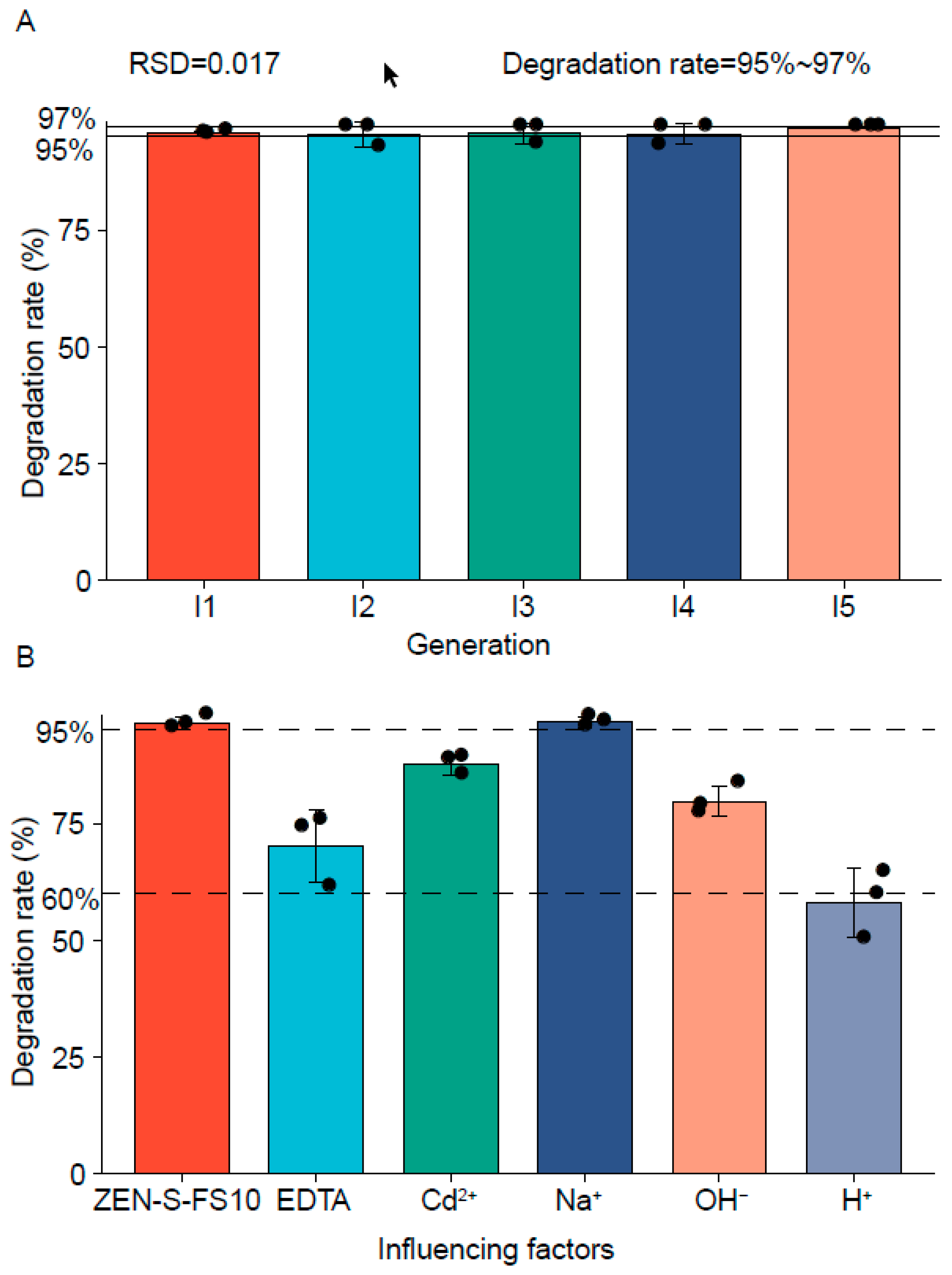
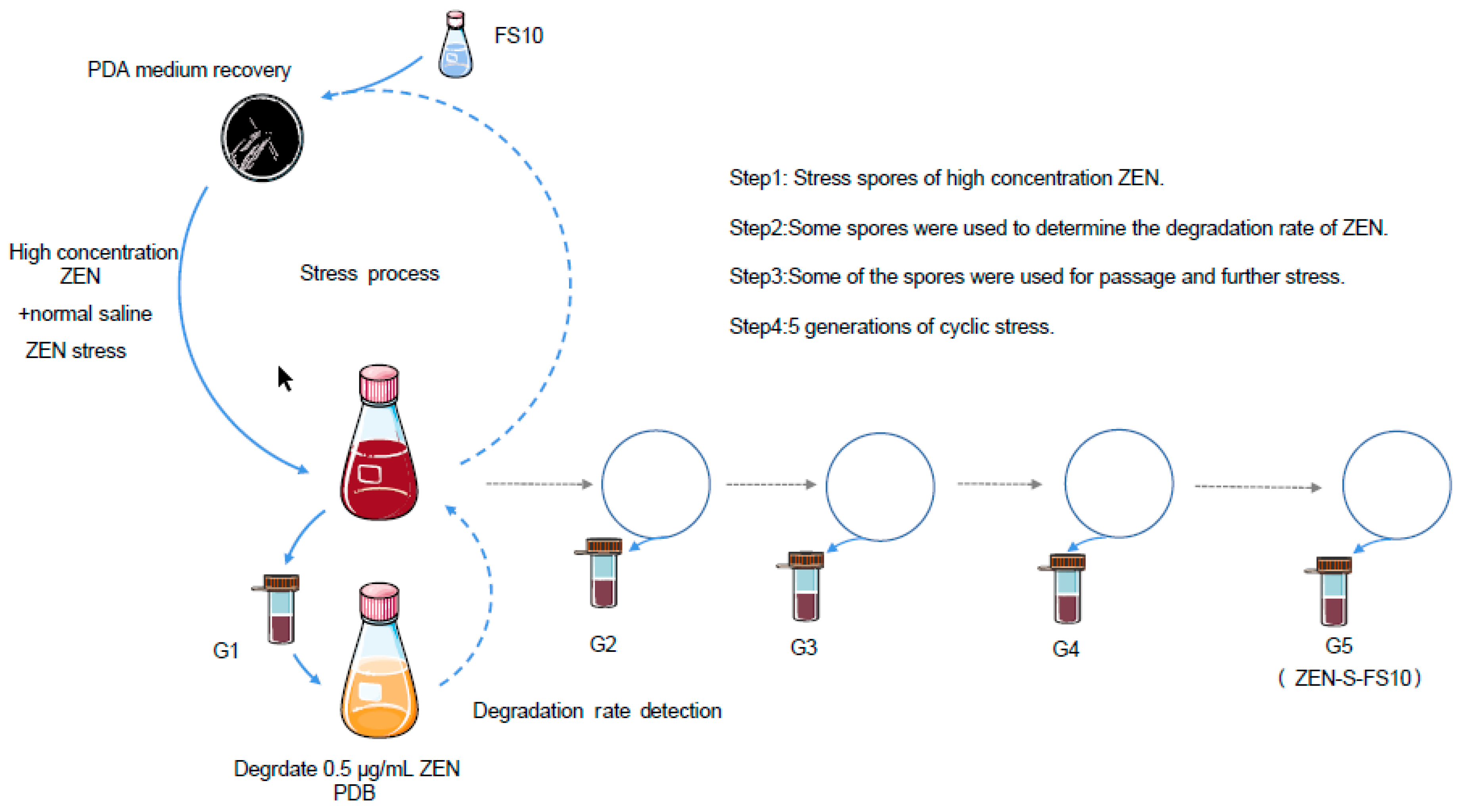
2.1. Analysis of the Strain Information and Degradation Rate
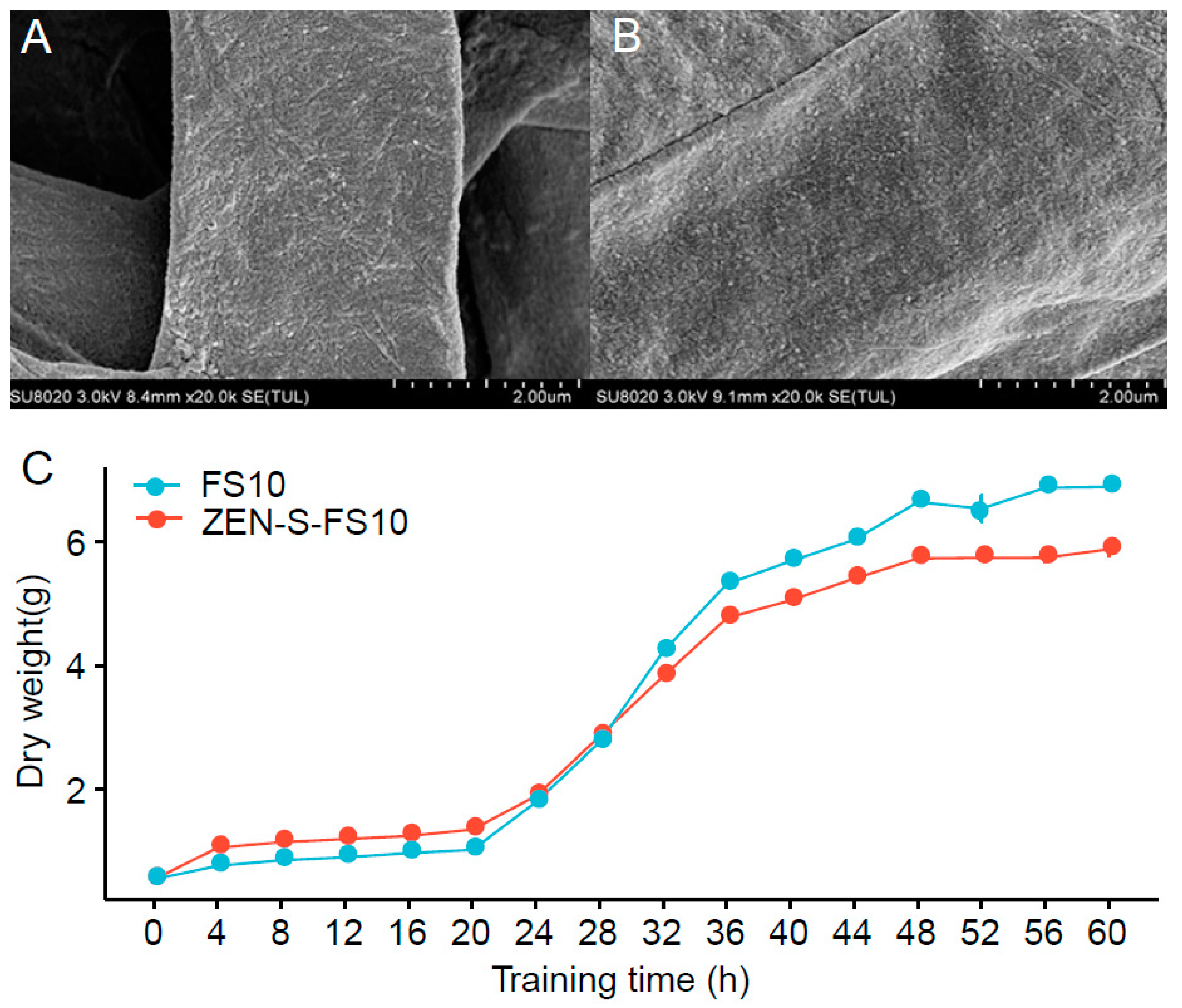
2.2. Effect of Stress on the Myceliume and Growth of Myceliume
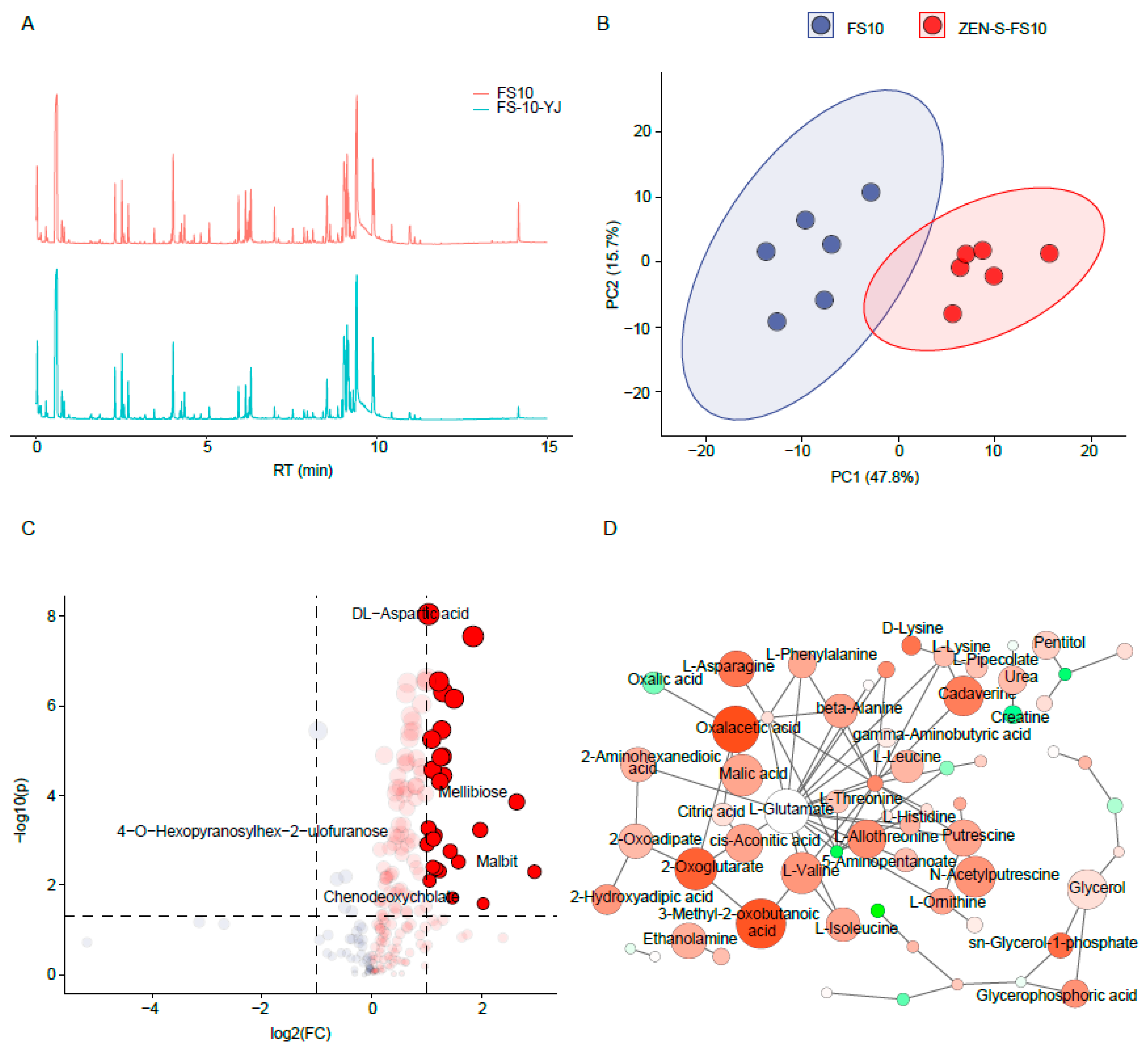
2.3. Mycelial Metabolomics Analysis
2.4. Analysis of the Optimization of Degradation Conditions
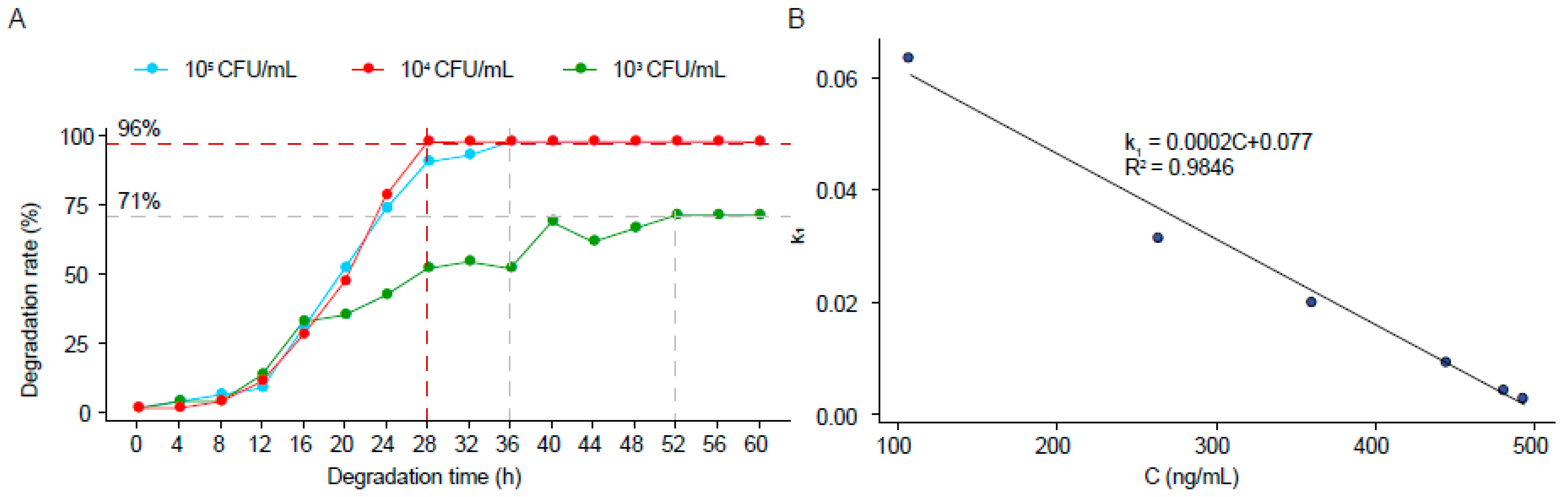
2.5. Kinetic Study of ZEN Degradation
2.6. Stability of Strain Degradability Analysis
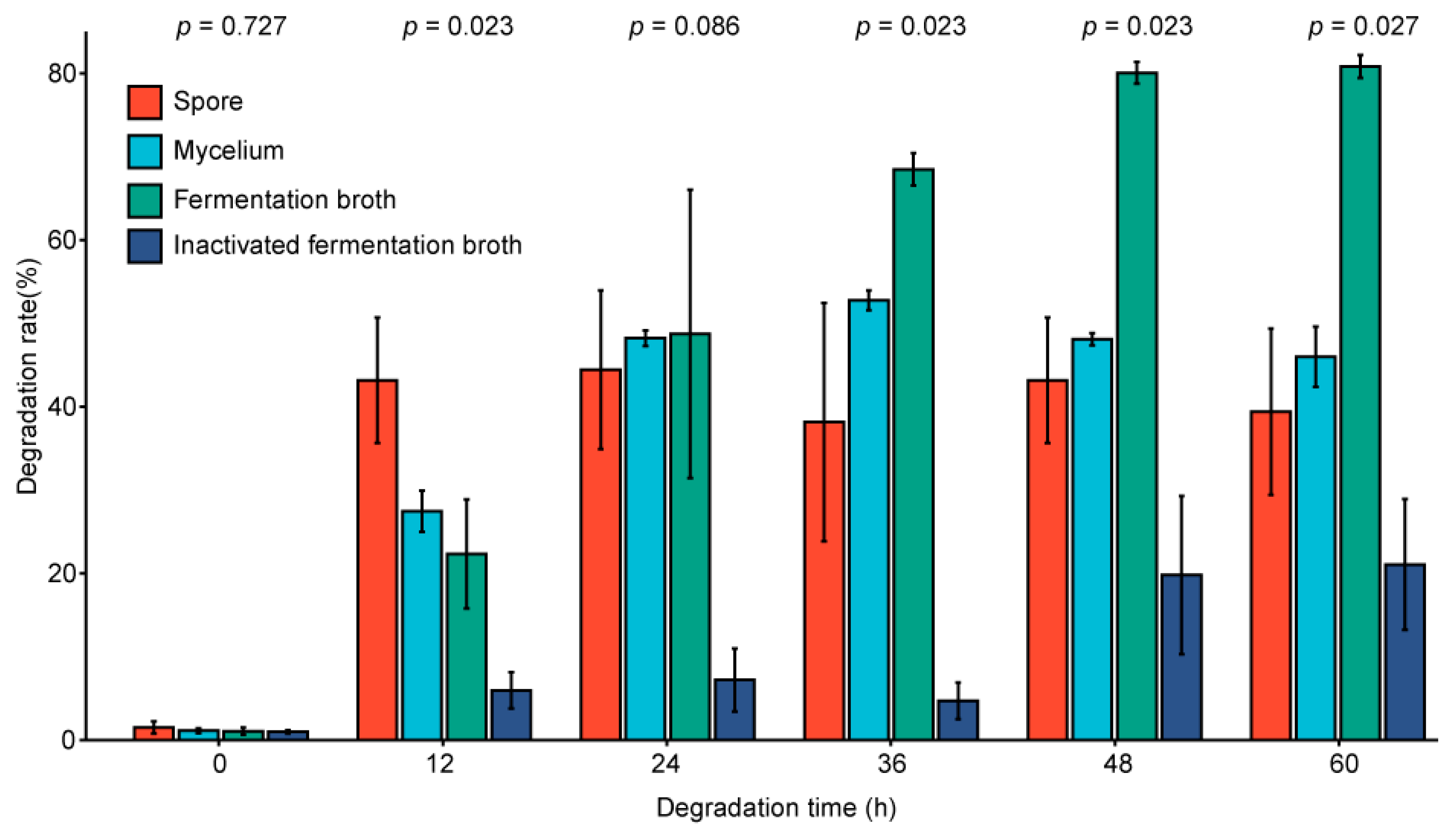
2.7. Analysis of the Influence of Fermentation Environment Changes in Degradation Ability
2.8. Analysis of the Degradation Site of the Mycelium
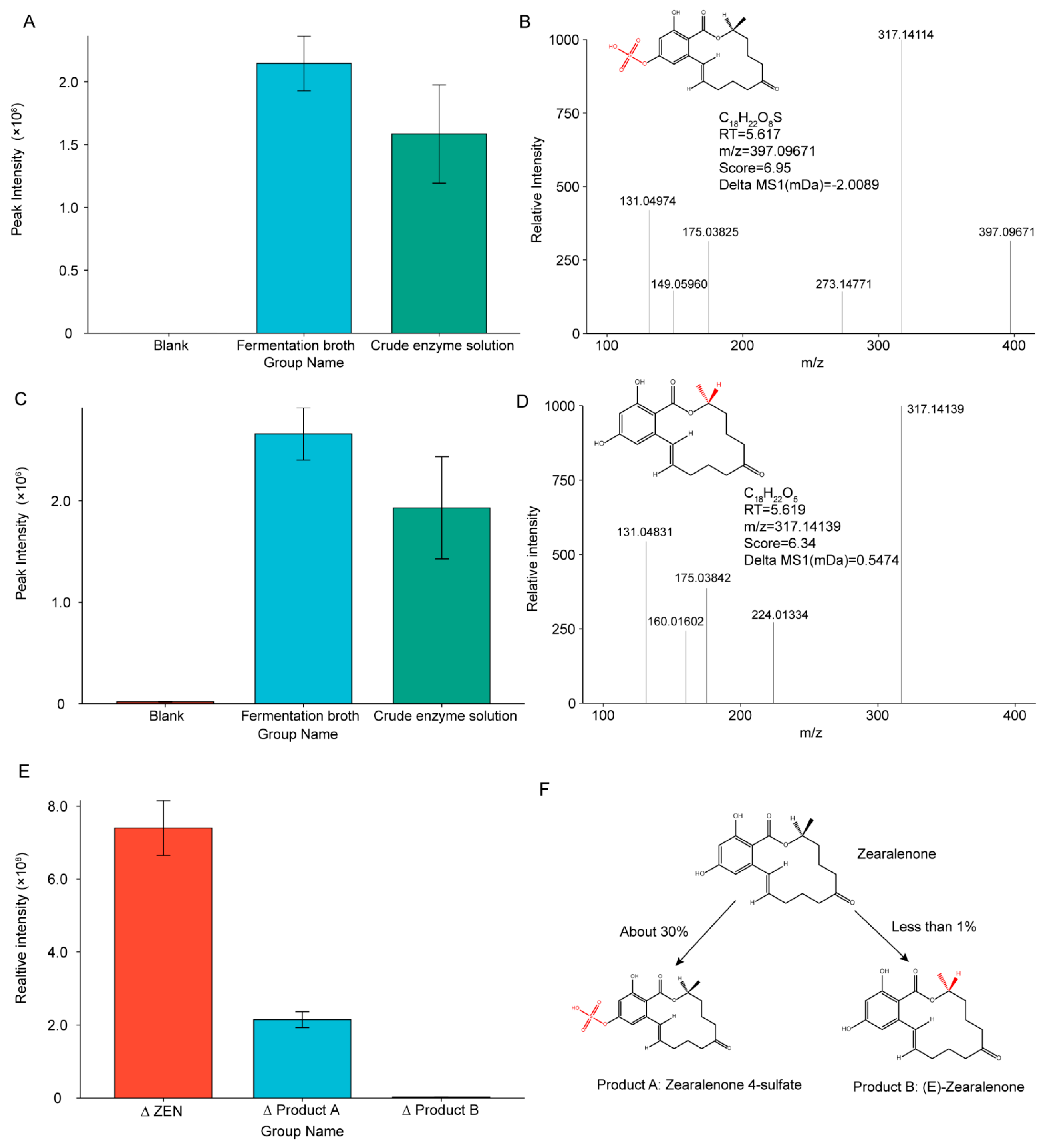
2.9. Analysis of Probable Degradation Products and Degradation Pathways
3. Conclusions and Prospect
4. Materials and Methods
4.1. FS10 and Chemicals
4.2. Coercion Method and Condition Optimization
4.3. Evaluation of the ZEN Degradation Rate
4.4. Effect of Stress on the Surface Structure and Growth of the Mycelium
4.5. Metabolome Analysis of Strains
4.6. Optimization of Degradation Conditions
4.7. Stability of Strain Degradability
4.8. Influence of Fermentation Environment on Degradation Ability
4.9. Study on the Degradation Site
4.10. Evaluation of ZEN Degradation Products
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Garcia, S.D.O.; Sibaja, K.V.M.; Nogueira, W.V.; Feltrin, A.C.P.; Pinheiro, D.F.A.; Cerqueira, M.B.R.; Furlong, E.B.; Garda-Buffon, J. Peroxidase as a simultaneous degradation agent of ochratoxin A and zearalenone applied to model solution and beer. Food Res. Int. 2020, 131, 109039. [Google Scholar] [CrossRef]
- Schaarschmidt, S.; Fauhl-Hassek, C. The fate of mycotoxins during secondary food processing of maize for human consumption. Compr. Rev. Food Sci. Food Saf. 2021, 20, 91–148. [Google Scholar] [CrossRef] [PubMed]
- Afsah-Hejri, L.; Hajeb, P.; Ehsani, R.J. Application of ozone for degradation of mycotoxins in food: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1777–1808. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Xu, J.; Jiang, K.; Liu, X.; Meng, J.; Di Mavungu, J.D.; Guo, W.; Zhang, Z.; Jing, J.; Li, H.; et al. Determination of multiple mycotoxins in paired plasma and urine samples to assess human exposure in Nanjing, China. Environ. Pollut. 2019, 248, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Sun, X.; Wan, X.; Li, H.; Yan, F.; Han, R.; Li, H.; Li, Z.; Tian, Y.; Liu, X.; et al. Identification of a Bacillus amyloliquefaciens H6 Thioesterase Involved in Zearalenone Detoxification by Transcriptomic Analysis. J. Agric. Food Chem. 2020, 68, 10071–10080. [Google Scholar] [CrossRef]
- Ponzilacqua, B.; Rottinghaus, G.; Landers, B.; Oliveira, C. Effects of medicinal herb and Brazilian traditional plant extracts on in vitro mycotoxin decontamination. Food Control 2019, 100, 24–27. [Google Scholar] [CrossRef]
- Chen, F.; Wen, X.; Lin, P.; Chen, H.; Wang, A.; Jin, Y. HERP depletion inhibits zearalenone-induced apoptosis through autophagy activation in mouse ovarian granulosa cells. Toxicol. Lett. 2019, 301, 1–10. [Google Scholar] [CrossRef]
- Wagacha, J.; Muthomi, J. Mycotoxin problem in Africa: Current status, implications to food safety and health and possible management strategies. Int. J. Food Microbiol. 2008, 124, 1–12. [Google Scholar] [CrossRef]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef]
- Gott, P.N.; Hendel, E.G.; Ramirez, S.M.; Hofstetter, U.; Murugesan, G.R. Recent Mycotoxin Contamination Trends in Us Corn Grain and Corn by-Product Feeds. J. Dairy Sci. 2020, 103, 126. [Google Scholar]
- Luo, X.; Zhai, Y.; Qi, L.; Pan, L.; Wang, J.; Xing, J.; Wang, R.; Wang, L.; Zhang, Q.; Yang, K.; et al. Influences of Electron Beam Irradiation on the Physical and Chemical Properties of Zearalenone- and Ochratoxin A-Contaminated Corn and In Vivo Toxicity Assessment. Foods 2020, 9, 376. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Li, K.; Pan, L.; Luo, X.; Xing, J.; Wang, J.; Wang, L.; Wang, R.; Zhai, Y.; Chen, Z. Effect of Ozone and Electron Beam Irradiation on Degradation of Zearalenone and Ochratoxin A. Toxins 2020, 12, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.J.; Kang, J.S.; Cho, W.T.; Lee, C.H.; Ha, J.K.; Bin Song, K. In vitro degradation of zearalenone by Bacillus subtilis. Biotechnol. Lett. 2010, 32, 1921–1924. [Google Scholar] [CrossRef] [PubMed]
- Kosawang, C.; Karlsson, M.; Vélëz, H.; Rasmussen, P.H.; Collinge, D.B.; Jensen, B.; Jensen, D. Zearalenone detoxification by zearalenone hydrolase is important for the antagonistic ability of Clonostachys rosea against mycotoxigenic Fusarium graminearum. Fungal Biol. 2014, 118, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Vekiru, E.; Frühauf, S.; Hametner, C.; Schatzmayr, G.; Krska, R.; Moll, W.; Schuhmacher, R. Isolation and characterisation of enzymatic zearalenone hydrolysis reaction products. World Mycotoxin J. 2016, 9, 353–363. [Google Scholar] [CrossRef]
- Wang, H.; Silong, S.; Wenyang, G.; Lanfei, Z.; Bingqian, H.; Kai, W.; Zhongfan, L.; Liyang, C.; Shoushen, X.; Jun, G.; et al. Horizontal Gene Transfer of Fhb7 from Fungus Underlies Fusarium Head Blight Resistance in Wheat. Science 2020, 368, 6493. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Liu, X.; Li, J. Updating techniques on controlling mycotoxins—A review. Food Control 2018, 89, 123–132. [Google Scholar] [CrossRef]
- Kim, S.H.; Vujanovic, V. Biodegradation and Biodetoxification of Fusarium Mycotoxins by Sphaerodes Mycoparasitica. AMB Express 2017, 7, 145. [Google Scholar] [CrossRef]
- Xu, D.; Wang, H.; Zhang, Y.; Yang, Z.; Sun, X. Inhibition of non-toxigenic Aspergillus niger FS10 isolated from Chinese fermented soybean on growth and aflatoxin B1 production by Aspergillus flavus. Food Control 2013, 32, 359–365. [Google Scholar] [CrossRef]
- He, M.; Li, Y.; Pi, F.; Ji, J.; He, X.; Zhang, Y.; Sun, X. A novel detoxifying agent: Using rice husk carriers to immobilize zearalenone-degrading enzyme from Aspergillus niger FS10. Food Control 2016, 68, 271–279. [Google Scholar] [CrossRef]
- Sebaei, A.S.; Sobhy, H.M.; Fouzy, A.; Hussain, O.A. Occurrence of zearalenone in grains and its reduction by gamma radiation. Int. J. Environ. Anal. Chem. 2020, 9, 1–9. [Google Scholar] [CrossRef]
- González-Salgado, A.; Patiño, B.; Vázquez, C.; González-Jaén, M.T. Discrimination of Aspergillus Niger and Other Aspergillus Species Belonging to Section Nigri by Pcr Assays. FEMS Microbiol. Lett. 2005, 245, 353–361. [Google Scholar] [CrossRef]
- Sun, X.; Xingxing, H.; Siyu, X.K.; Yun, L.; Dan, X.; He, Q. Biological Detoxification of Zearalenone by Aspergillus Niger Strain Fs10. Food Chem. Toxicol. 2014, 72, 76–82. [Google Scholar] [CrossRef]
- Watanabe, C.; Terashima, I.; Noguchi, K. Responses of the Respiratory Chain to Low Temperature Stress. Plant Cell Physiol. 2007, 48, S181. [Google Scholar]
- Jensen, T.; De Boevre, M.; De Saeger, S.; Preußke, N.; Sönnichsen, F.D.; Kramer, E.; Klink, H.; Verreet, J.-A.; Birr, T. Effect of ensiling duration on the fate of deoxynivalenol, zearalenone and their derivatives in maize silage. Mycotoxin Res. 2020, 36, 127–136. [Google Scholar] [CrossRef]
- Calado, T.; Abrunhosa, L.; Verde, S.C.; Alté, L.; Venâncio, A.; Fernández-Cruz, M.L. Effect of Gamma-Radiation on Zearalenone—Degradation, Cytotoxicity and Estrogenicity. Foods 2020, 9, 1687. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-J.; Zhang, R.-Q.; Zhai, Q.-Y.; Liu, J.-C.; Li, N.; Liu, W.-X.; Li, L.; Shen, W. Metagenomic analysis of gut microbiota alteration in a mouse model exposed to mycotoxin deoxynivalenol. Toxicol. Appl. Pharmacol. 2019, 372, 47–56. [Google Scholar] [CrossRef]
- Essex-Lopresti, A.; David, J.; Moore, K.; Armstrong, S.; Auton, P.; Rutter, S.; Green, C.; Hiscox, J.A.; Paszkiewicz, K.H.; Clark, G.C. Unbiased whole genome sequence analysis of the murine transcriptome in response to ricin exposure via the pulmonary route. Toxicon 2019, 158, S6. [Google Scholar] [CrossRef]
- Wang, J.; Leipeng, C.; Yuhuan, L.; Qi, Z.; Roger, R.; Xuan, L. Effect of Acclimatized Paddy Soil Microorganisms Using Swine Wastewater on Degradation of Rice Straw. Bioresour. Technol. 2021, 332, 125039. [Google Scholar] [CrossRef] [PubMed]
- Jahoor, F.; Jackson, A.; Golden, M. In vivo Metabolism of Nitrogen Precursors for Urea Synthesis in the Postprandial Rat. Ann. Nutr. Metab. 1988, 32, 240–244. [Google Scholar] [CrossRef]
- de Sousa, R.T.; Streck, E.L.; Forlenza, O.V.; Brunoni, A.R.; Zanetti, M.V.; Ferreira, G.K.; Diniz, B.; Portela, L.V.; Carvalho, A.F.; Zarate, C.A.; et al. Regulation of leukocyte tricarboxylic acid cycle in drug-naïve Bipolar Disorder. Neurosci. Lett. 2015, 605, 65–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Zhou, J.; Tang, Y.; Ma, Q.; Zhang, J.; Ji, C.; Zhao, L. Characterization and Genome Analysis of a Zearalenone-Degrading Bacillus velezensis Strain ANSB01E. Curr. Microbiol. 2019, 77, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Changyong, W.; Jian’e, Z.; Yuexi, Z.; Jin, Y. Residual Ozone in Microorganisms Enhanced Organics Removal and Shaped Microbial Community. Chemosphere 2021, 278, 130322. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yuhui, W.; Ying, L.; Min, S.; Wenfang, C. Microorganisms and Their Metabolic Activities Affect Seepage through Porous Media in Groundwater Artificial Recharge Systems: A Review. J. Hydrol. 2021, 598, 126256. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, W.; Wu, H.; Zhang, W.; Mu, W. Identification of a Potent Enzyme for the Detoxification of Zearalenone. J. Agric. Food Chem. 2019, 68, 376–383. [Google Scholar] [CrossRef]
- Kiseleva, M.; Chalyy, Z.; Sedova, I.; Aksenov, I. Stability of Mycotoxins in Individual Stock and Multi-Analyte Standard Solutions. Toxins 2020, 12, 94. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, M.F.; Girgin, G.; Baydar, T.; Krska, R.; Sulyok, M. Occurrence of multiple mycotoxins and other fungal metabolites in animal feed and maize samples from Egypt using LC-MS/MS. J. Sci. Food Agric. 2017, 97, 4419–4428. [Google Scholar] [CrossRef]
- Vendl, O.; Berthiller, F.; Crews, C.; Krska, R. Simultaneous determination of deoxynivalenol, zearalenone, and their major masked metabolites in cereal-based food by LC–MS–MS. Anal. Bioanal. Chem. 2009, 395, 1347–1354. [Google Scholar] [CrossRef]
- Plasencia, J.; Mirocha, C.J. Isolation and Characterization of Zearalenone Sulfate Produced by Fusarium spp. Appl. Environ. Microbiol. 1991, 57, 146–150. [Google Scholar] [CrossRef] [Green Version]
- De Boevre, M.; Di Mavungu, J.; Maene, P.; Audenaert, K.; Deforce, D.; Haesaert, G.; Eeckhout, M.; Callebaut, A.; Berthiller, F.; Van Peteghem, C.; et al. Development and validation of an LC-MS/MS method for the simultaneous determination of deoxynivalenol, zearalenone, T-2-toxin and some masked metabolites in different cereals and cereal-derived food. Food Addit. Contam. Part A-Chem. Anal. Control Expo. Risk Assess. 2012, 29, 819–835. [Google Scholar] [CrossRef]
- Wu, S.; Wang, F.; Li, Q.; Wang, J.; Zhou, Y.; Duan, N.; Niazi, S.; Wang, Z. Photocatalysis and Degradation Products Identification of Deoxynivalenol in Wheat Using Upconversion Nanoparticles@Tio2 Composite. Food Chem. 2020, 323, 9. [Google Scholar] [CrossRef]
- Tinyiro, S.E.; Wokadala, C.; Xu, D.; Yao, W. Adsorption and degradation of zearalenone by bacillus strains. Folia Microbiol. 2011, 56, 321–327. [Google Scholar] [CrossRef]
- Al-Nussairawi, M.; Risa, A.; Garai, E.; Varga, E.; Szabó, I.; Csenki-Bakos, Z.; Kriszt, B.; Cserháti, M. Mycotoxin Biodegradation Ability of the Cupriavidus Genus. Curr. Microbiol. 2020, 77, 2430–2440. [Google Scholar] [CrossRef]
- Wang, X.; Qin, X.; Hao, Z.; Luo, H.; Yao, B.; Su, X. Degradation of Four Major Mycotoxins by Eight Manganese Peroxidases in Presence of a Dicarboxylic Acid. Toxins 2019, 11, 566. [Google Scholar] [CrossRef] [Green Version]
- Fruhauf, S.; Thamhesl, M.; Pfeffer, M.; Moll, D.; Schatzmayr, G.; Binder, E.M. Polypeptide for Hydrolytic Cleavage of Zearalenone and/or Zearalenone Derivatives, Isolated Polynucleotide Thereof as Well as a Polypeptide Containing an Additive, Use of Same as Well as a Process; Erber AG: Getzersdorf, Austria, 2020. [Google Scholar]
- Janković-Tomanić, M.; Petković, B.; Todorović, D.; Vranković, J.; Perić-Mataruga, V. Physiological and behavioral effects of the mycotoxin deoxynivalenol in Tenebrio molitor larvae. J. Stored Prod. Res. 2019, 83, 236–242. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, M.; Yang, Q.; Apaliya, M.T.; Li, J.; Zhang, X. Biodegradation of zearalenone by Saccharomyces cerevisiae: Possible involvement of ZEN responsive proteins of the yeast. J. Proteom. 2016, 143, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Adebo, O.A.; Kayitesi, E.; Njobeh, P.B. Reduction of Mycotoxins during Fermentation of Whole Grain Sorghum to Whole Grain Ting (a Southern African Food). Toxins 2019, 11, 180. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Chang, J.; Wang, P.; Yin, Q.; Huang, W.; Dang, X.; Lu, F.; Gao, T. Zearalenone Biodegradation by the Combination of Probiotics with Cell-Free Extracts of Aspergillus oryzae and its Mycotoxin-Alleviating Effect on Pig Production Performance. Toxins 2019, 11, 552. [Google Scholar] [CrossRef] [Green Version]
- Qiu, T.; Haiming, W.; Yang, Y.; Jian, Y.; Jian, J.; Jiadi, S.; Shuang, Z.; Xiulan, S. Exploration of Biodegradation Mechanism by Afb1-Degrading Strain Aspergillus Niger Fs10 and Its Metabolic Feedback. Food Control. 2021, 121, 107609. [Google Scholar] [CrossRef]
- Lyagin, I.; Efremenko, E. Enzymes for Detoxification of Various Mycotoxins: Origins and Mechanisms of Catalytic Action. Molecules 2019, 24, 2362. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, C.; Zhang, D.; Zhao, M.; Peng, M.; Guo, P.; Cui, Z. Microbial Degradation of Zearalenone by a Novel Microbial Consortium, NZDC-6, and Its Application on Contaminated Corncob by Semisolid Fermentation. J. Agric. Food Chem. 2020, 68, 1634–1644. [Google Scholar] [CrossRef] [PubMed]
- Goessens, T.; De Baere, S.; De Troyer, N.; Deknock, A.; Goethals, P.; Lens, L.; Pasmans, F.; Croubels, S. Multi-residue analysis of 20 mycotoxins including major metabolites and emerging mycotoxins in freshwater using UHPLC-MS/MS and application to freshwater ponds in flanders, Belgium. Environ. Res. 2021, 196, 110366. [Google Scholar] [CrossRef] [PubMed]
- de Paula, G.S.; Oliveira, M.C.; Sales, L.S.; Boriollo, M.; Rodrigues, L.K.; Nobre-dos-Santos, M.; Steiner-Oliveira, C. Antimicrobial Photodynamic Therapy Mediated by Methylene Blue Coupled to Β-Cyclodextrin Reduces Early Colonizing Microorganisms from the Oral Biofilm. Photodiagn. Photodyn. Ther. 2021, 34, 102283. [Google Scholar] [CrossRef] [PubMed]
- Dellafiora, L.; Gonaus, C.; Streit, B.; Galaverna, G.; Moll, W.D.; Vogtentanz, G.; Schatzmayr, G.; Dall’Asta, C.; Prasad, S. An in Silico Target Fishing Approach to Identify Novel Ochratoxin a Hydrolyzing Enzyme. Toxins 2020, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, B.; Han, S.-K.; Yoon, K.-S.; Sung, J.-M. Morphological Characteristics of Conidiogenesis inCordyceps militaris. Mycobiology 2005, 33, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Gbashi, S.; Madala, N.E.; De Saeger, S.; De Boevre, M.; Njobeh, P.B. Numerical optimization of temperature-time degradation of multiple mycotoxins. Food Chem. Toxicol. 2019, 125, 289–304. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, J.; Yu, J.; Yang, Y.; Yuan, X.; Yang, J.; Zhang, Y.; Sun, J.; Sun, X. Exploration on the Enhancement of Detoxification Ability of Zearalenone and Its Degradation Products of Aspergillus niger FS10 under Directional Stress of Zearalenone. Toxins 2021, 13, 720. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100720
Ji J, Yu J, Yang Y, Yuan X, Yang J, Zhang Y, Sun J, Sun X. Exploration on the Enhancement of Detoxification Ability of Zearalenone and Its Degradation Products of Aspergillus niger FS10 under Directional Stress of Zearalenone. Toxins. 2021; 13(10):720. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100720
Chicago/Turabian StyleJi, Jian, Jian Yu, Yang Yang, Xiao Yuan, Jia Yang, Yinzhi Zhang, Jiadi Sun, and Xiulan Sun. 2021. "Exploration on the Enhancement of Detoxification Ability of Zearalenone and Its Degradation Products of Aspergillus niger FS10 under Directional Stress of Zearalenone" Toxins 13, no. 10: 720. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100720