Pyrrolizidine Alkaloid-Induced Hepatotoxicity Associated with the Formation of Reactive Metabolite-Derived Pyrrole–Protein Adducts


Abstract
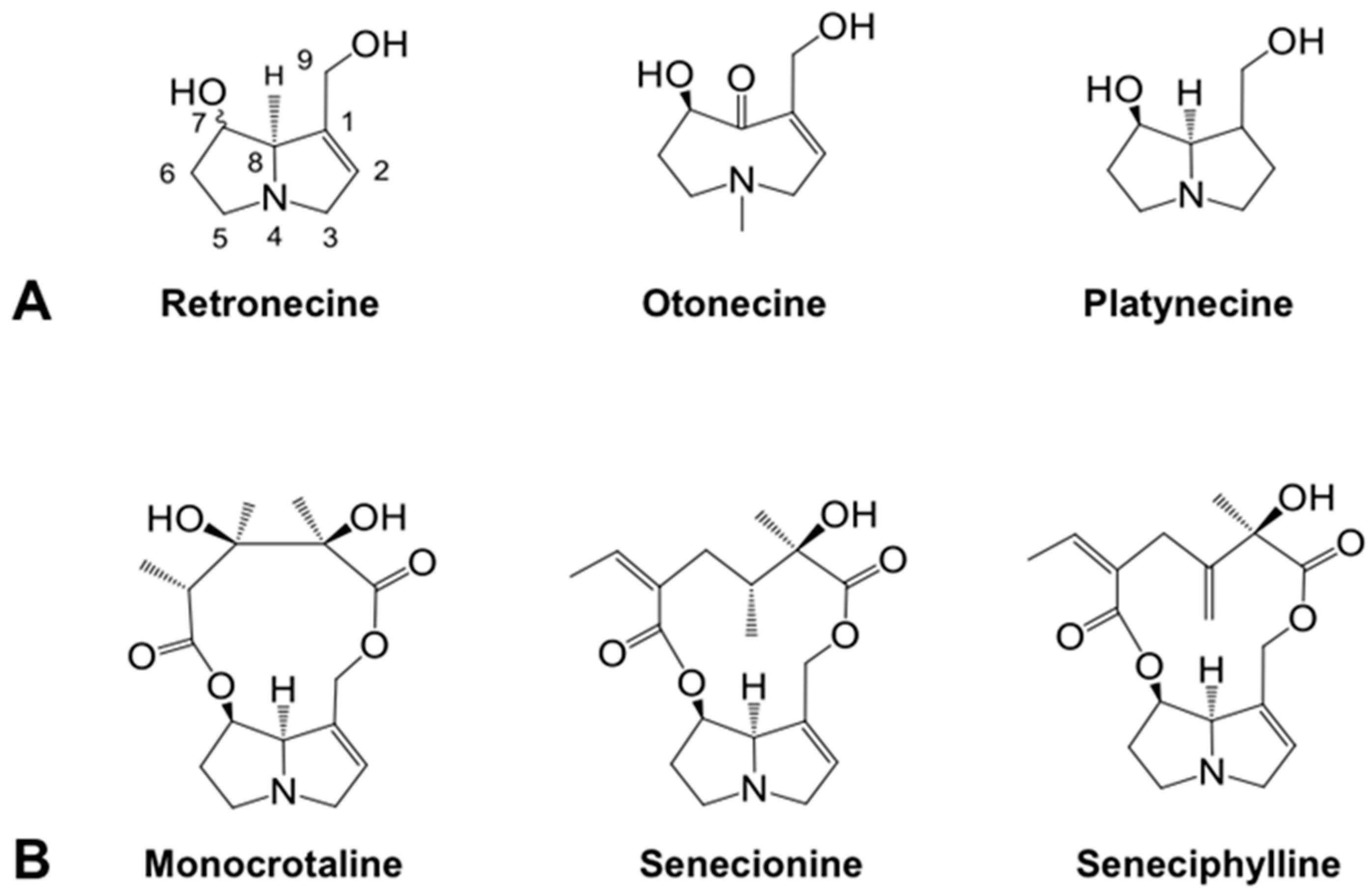
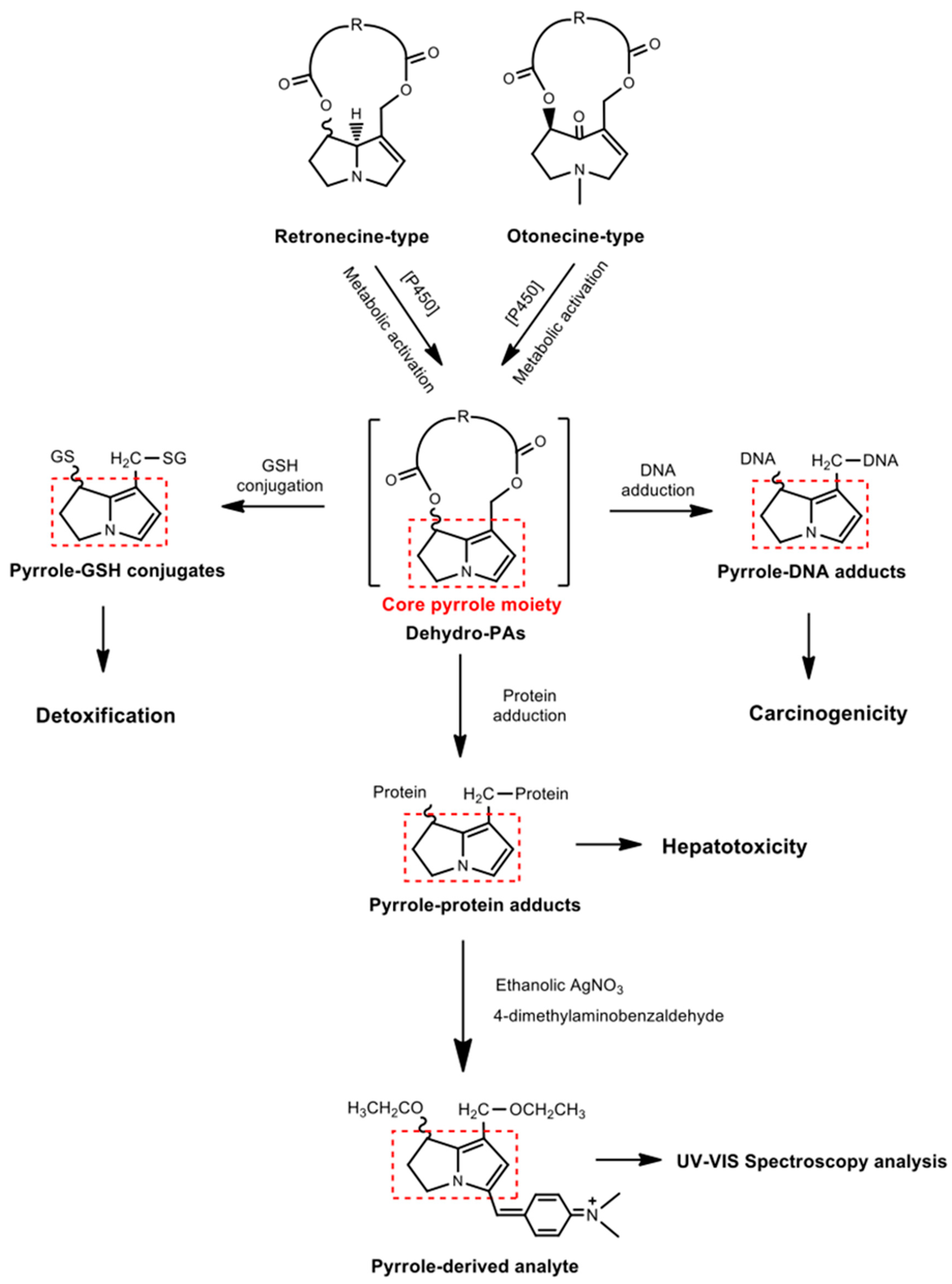
:1. Introduction
2. Results
2.1. Effects of C. sessiliflora Extract on Rats
2.1.1. Effect on Serum ALT Activity
2.1.2. Effect on Hepatic GSH Level
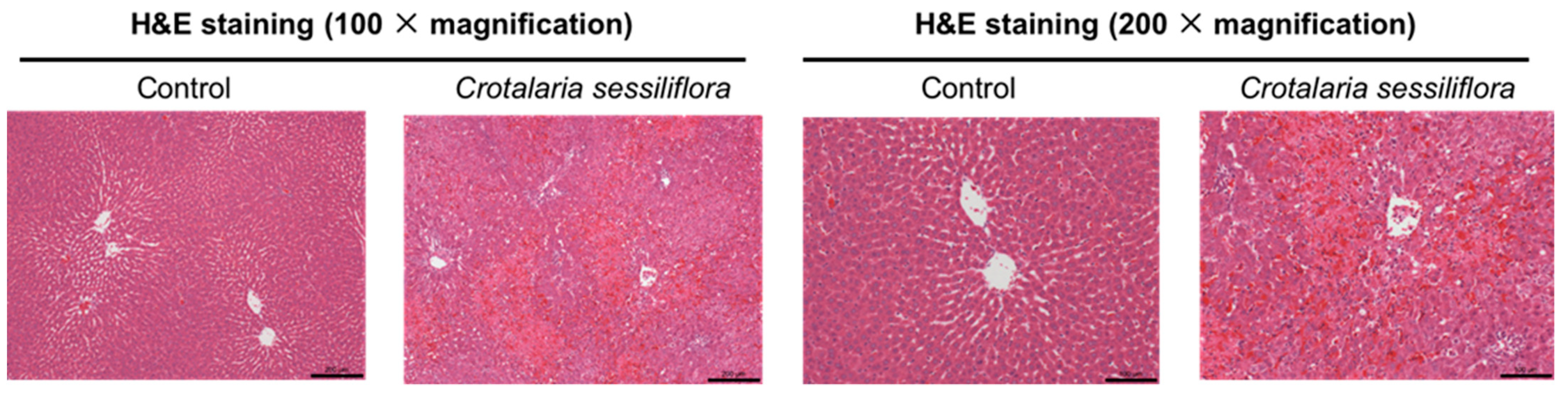
2.1.3. Effect on Liver Histological Changes
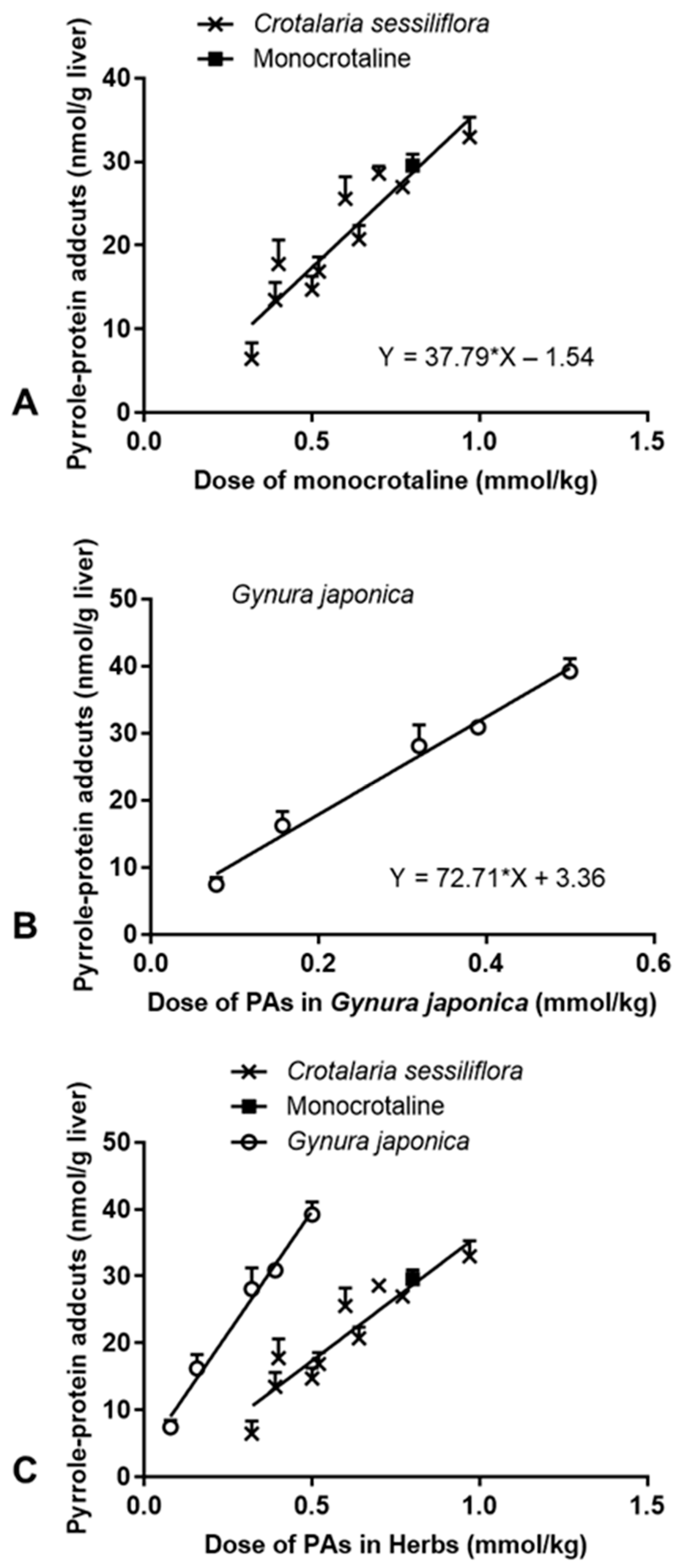
2.1.4. Effect on Formation of Hepatic Pyrrole–Protein Adducts
2.2. Effects of G. japonica Extract on Rats
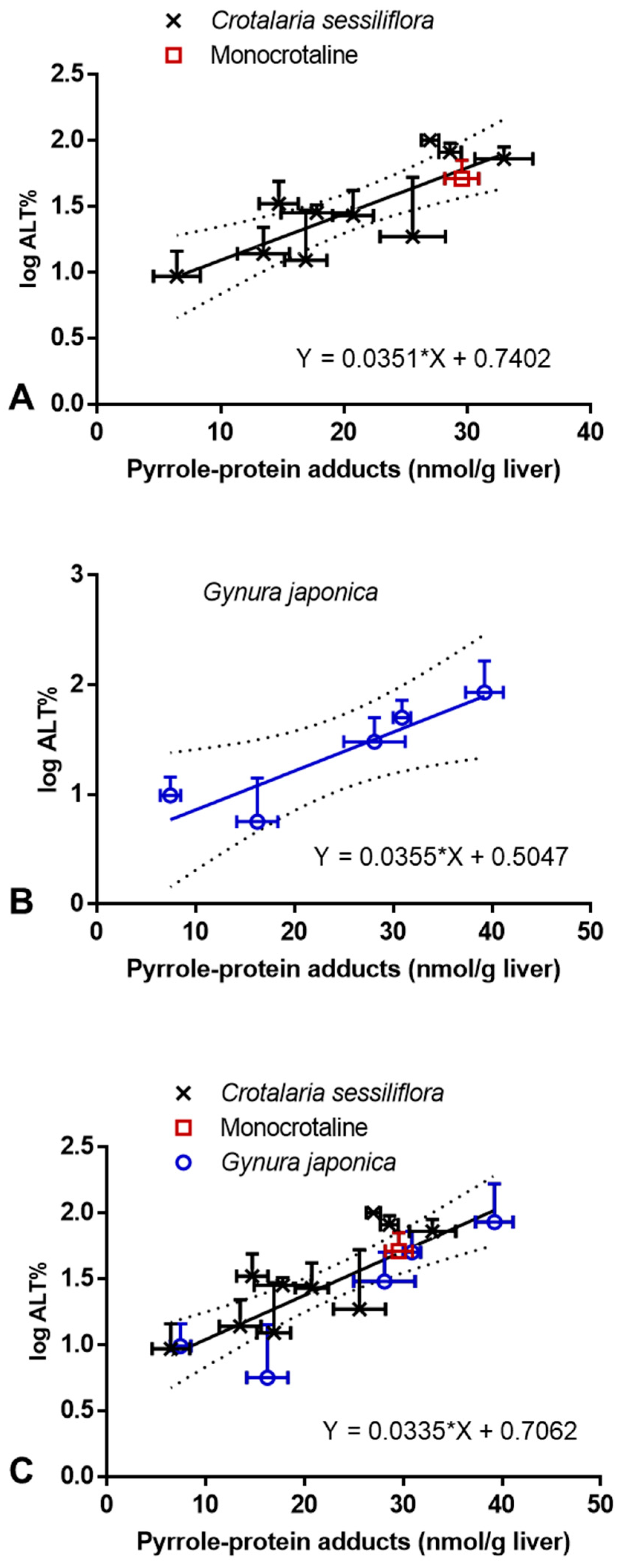
2.3. Correlation between PA-ILI and Formation of Hepatic Pyrrole–Protein Adducts
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Materials
4.3. Treatment of Rats with Herbal Extracts
4.4. Biochemical Assays and Histological Determination
4.5. Detection of Pyrrole–Protein Adducts in Liver Samples
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Schrenk, D.; Gao, L.; Lin, G.; Mahony, C.; Mulder, P.P.J.; Peijnenburg, A.; Pfuhler, S.; Rietjens, I.M.; Rutz, L.; Steinhoff, B.; et al. Pyrrolizidine alkaloids in food and phytomedicine: Occurrence, exposure, toxicity, mechanisms, and risk assessment—A review. Food Chem. Toxicol. 2020, 136, 111107. [Google Scholar] [CrossRef]
- Ma, J.; Xia, Q.; Fu, P.P.; Lin, G. Pyrrole-protein adducts—A biomarker of pyrrolizidine alkaloid-induced hepatotoxicity. J. Food Drug Anal. 2018, 26, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Moreira, R.; Pereira, D.M.; Valentão, P.; Andrade, P.B. Pyrrolizidine alkaloids: Chemistry, pharmacology, toxicology and food safety. Int. J. Mol. Sci. 2018, 19, 1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, J.A.; Molyneux, R.J.; Colegate, S.M. Pyrrolizidine alkaloids: Potential role in the etiology of cancers, pulmonary hypertension, congenital anomalies, and liver disease. Chem. Res. Toxicol. 2015, 28, 4–20. [Google Scholar] [CrossRef]
- Mattocks, A.R. Chemistry and Toxicology of Pyrrolizidine Alkaloids; Academic Press: Orlando, FL, USA, 1986. [Google Scholar]
- Ruan, J.; Yang, M.; Fu, P.P.; Ye, Y.; Lin, G. Metabolic activation of pyrrolizidine alkaloids: Insights into the structural and enzymatic basis. Chem. Res. Toxicol. 2014, 27, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Liao, C.; Ye, Y.; Lin, G. Lack of metabolic activation and predominant formation of an excreted metabolite of nontoxic platynecine-type pyrrolizidine alkaloids. Chem. Res. Toxicol. 2014, 27, 7–16. [Google Scholar] [CrossRef]
- Lu, Y.; Ma, J.; Song, Z.; Ye, Y.; Fu, P.P.; Lin, G. The role of formation of pyrrole-ATP synthase subunit beta adduct in pyrrolizidine alkaloid-induced hepatotoxicity. Arch. Toxicol. 2018, 92, 3403–3414. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Ruan, J.; Chen, X.; Li, D.; Yao, S.; Fu, P.P.; Ye, Y.; Gao, H.; Wang, J.; Lin, G. Pyrrole-hemoglobin adducts, a more feasible potential biomarker of pyrrolizidine alkaloid exposure. Chem. Res. Toxicol. 2019, 32, 1027–1039. [Google Scholar] [CrossRef]
- Yang, X.Q.; Ye, J.; Li, X.; Li, Q.; Song, Y.H. Pyrrolizidine alkaloids-induced hepatic sinusoidal obstruction syndrome: Pathogenesis, clinical manifestations, diagnosis, treatment, and outcomes. World J. Gastroenterol. 2019, 25, 3753–3763. [Google Scholar] [CrossRef]
- Hessel-Pras, S.; Braeuning, A.; Guenther, G.; Adawy, A.; Enge, A.M.; Ebmeyer, J.; Henderson, C.J.; Hengstler, J.G.; Lampen, A.; Reif, R. The pyrrolizidine alkaloid senecionine induces CYP-dependent destruction of sinusoidal endothelial cells and cholestasis in mice. Arch. Toxicol. 2020, 94, 219–229. [Google Scholar] [CrossRef]
- Fu, P.P. Pyrrolizidine alkaloids: Metabolic activation pathways leading to liver tumor initiation. Chem. Res. Toxicol. 2017, 30, 81–93. [Google Scholar] [CrossRef]
- Yan, C.C.; Huxtable, R.J. Relationship between glutathione concentration and metabolism of the pyrrolizidine alkaloid, monocrotaline, in the isolated, perfused liver. Toxicol. Appl. Pharm. 1995, 130, 132–139. [Google Scholar] [CrossRef]
- Chen, M.X.; Li, L.; Zhong, D.F.; Shen, S.J.; Zheng, J.; Chen, X.Y. 9-Glutathiony1-6,7-dihydro-1-hydroxymethy1-5H-pyrrolizine is the major pyrrolic glutathione conjugate of retronecine-type pyrrolizidine alkaloids in liver microsomes and in rats. Chem. Res. Toxicol. 2016, 29, 180–189. [Google Scholar] [CrossRef]
- He, X.; Xia, Q.; Fu, P.P. 7-Glutathione-pyrrole and 7-cysteine-pyrrole are potential carcinogenic metabolites of pyrrolizidine alkaloids. J. Environ. Sci. Health C 2017, 35, 69–83. [Google Scholar] [CrossRef]
- Lin, G.; Cui, Y.Y.; Hawes, E.M. Microsomal formation of a pyrrolic alcohol glutathione conjugate of clivorine firm evidence for the formation of a pyrrolic metabolite of an otonecine-type pyrrolizidine alkaloid. Drug Metab. Dispos. 1998, 26, 181–184. [Google Scholar]
- Yang, M.; Ruan, J.; Fu, P.P.; Lin, G. Cytotoxicity of pyrrolizidine alkaloid in human hepatic parenchymal and sinusoidal endothelial cells: Firm evidence for the reactive metabolites mediated pyrrolizidine alkaloid-induced hepatotoxicity. Chem. Biol. Interact. 2016, 243, 119–126. [Google Scholar] [CrossRef] [PubMed]
- DeLeve, L.D.; McCuskey, R.S.; Wang, X.; Hu, L.; McCuskey, M.K.; Epstein, R.B.; Kanel, G.C. Characterization of a reproducible rat model of hepatic veno-occlusive disease. Hepatology 1999, 29, 1779–1791. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Wang, J.; Li, N.; Li, M.; Gao, H.; Ji, Y.; Zhang, F.; Wang, H.; Zhou, Y.; Ye, Y.; et al. Hepatic sinusoidal obstruction syndrome associated with consumption of Gynura segetum. J. Hepatol. 2011, 54, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Chojkier, M. Hepatic sinusoidal-obstruction syndrome: Toxicity of pyrrolizidine alkaloids. J. Hepatol. 2003, 39, 437–446. [Google Scholar] [CrossRef]
- Willmot, F.C.; Robertson, G.W. Senecio disease, or cirrhosis of the liver due to senecio poisoning. Lancet 1920, 2, 848–849. [Google Scholar] [CrossRef] [Green Version]
- Goulding, R. Pyrrolizidine alkaloids—Environmental-health criteria-80. J. R. Soc. Health 1989, 109, 152. [Google Scholar]
- Kakar, F.; Akbarian, Z.; Leslie, T.; LaisMustafa, M.; Watson, J.; van Egmond, H.P.; Omar, M.F.; Mofleh, J. An outbreak of hepatic veno-occlusive disease in Western Afghanistan associated with exposure to wheat flour contaminated with pyrrolizidine alkaloids. J. Toxicol. 2010, 2010, 313280. [Google Scholar] [CrossRef]
- Ruan, J.; Gao, H.; Li, N.; Xue, J.; Chen, J.; Ke, C.; Ye, Y.; Fu, P.P.; Zheng, J.; Wang, J.; et al. Blood pyrrole-protein adducts-A biomarker of pyrrolizidine alkaloid-induced liver injury in humans. J. Environ. Sci. Health C 2015, 33, 404–421. [Google Scholar] [CrossRef]
- Wang, X.X.; Qi, X.S.; Guo, X.Z. Tusanqi-related sinusoidal obstruction syndrome in China a systematic review of the literatures. Medicine 2015, 94, e942. [Google Scholar] [CrossRef]
- Yang, M.; Ma, J.; Ruan, J.; Ye, Y.; Fu, P.P.; Lin, G. Intestinal and hepatic biotransformation of pyrrolizidine alkaloid N-oxides to toxic pyrrolizidine alkaloids. Arch. Toxicol. 2019, 93, 2197–2209. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhang, W.; He, Y.; Zhu, L.; Zhang, C.; Liu, J.; Ye, Y.; Zhuge, Y.; Lin, G. Clinical application of pyrrole-hemoglobin adducts as a biomarker of pyrrolizidine alkaloid exposure in humans. Arch. Toxicol. 2020, 95, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, C.; Li, D.; Chen, H.; Ma, J.; Gao, H.; Ye, Y.; Wang, J.; Fu, P.P.; Lin, G. Tu-San-Qi (Gynura japonica): The culprit behind pyrrolizidine alkaloid-induced liver injury in China. Acta Pharmacol. Sin. 2020, 42, 1212–1222. [Google Scholar] [CrossRef]
- Ozer, J.; Ratner, M.; Shaw, M.; Bailey, W.; Schomaker, S. The current state of serum biomarkers of hepatotoxicity. Toxicology 2008, 245, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Nnane, I.P.; Cheng, T.Y. The effects of pretreatment with glycyrrhizin and glycyrrhetinic acid on the retrorsine-induced hepatotoxicity in rats. Toxicon 1999, 37, 1259–1270. [Google Scholar] [CrossRef]
- Yan, C.C.; Huxtable, R.J. Effects of monocrotaline, a pyrrolizidine alkaloid, on glutathione metabolism in the rat. Biochem. Pharmacol. 1996, 51, 375–379. [Google Scholar] [CrossRef]
- Ho, V.T.; Revta, C.; Richardson, P.G. Hepatic veno-occlusive disease after hematopoietic stem cell transplantation: Update on defibrotide and other current investigational therapies. Bone Marrow Transplant. 2008, 41, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Dusemund, B.; Nowak, N.; Sommerfeld, C.; Lindtner, O.; Schafer, B.; Lampen, A. Risk assessment of pyrrolizidine alkaloids in food of plant and animal origin. Food Chem. Toxicol. 2018, 115, 63–72. [Google Scholar] [CrossRef]
- Merz, K.H.; Schrenk, D. Interim relative potency factors for the toxicological risk assessment of pyrrolizidine alkaloids in food and herbal medicines. Toxicol. Lett. 2016, 263, 44–57. [Google Scholar] [CrossRef]
- Ning, J.; Chen, L.; Strikwold, M.; Louisse, J.; Wesseling, S.; Rietjens, I.M. Use of an in vitro-in silico testing strategy to predict inter-species and inter-ethnic human differences in liver toxicity of the pyrrolizidine alkaloids lasiocarpine and riddelliine. Arch. Toxicol. 2019, 93, 801–818. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Zhu, L.; Ma, J.; Lin, G. Metabolism-mediated cytotoxicity and genotoxicity of pyrrolizidine alkaloids. Arch. Toxicol. 2021, 95, 1917–1942. [Google Scholar] [CrossRef] [PubMed]
- Zhuge, Y.; Liu, Y.; Xie, W.; Zou, X.; Xu, J.; Wang, J. Chinese society of gastroenterology committee of hepatobiliary disease. Expert consensus on the clinical management of pyrrolizidine alkaloid-induced hepatic sinusoidal obstruction syndrome. J. Gastroenterol. Hepatol. 2019, 34, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Danan, G.; Teschke, R. RUCAM in drug and herb induced liver injury: The update. Int. J. Mol. Sci. 2016, 17, 14. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R.; Danan, G. Worldwide use of RUCAM for causality assessment in 81,856 DILI and 14,029 HILI cases published 1993—Mid 2020, A comprehensive analysis. Medicines 2020, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.C.; Mao, Y.M.; Chen, C.W.; Chen, J.J.; Chen, J.; Cong, W.M.; Ding, Y.; Duan, Z.P.; Fu, Q.C.; Guo, X.Y.; et al. CSH guidelines for the diagnosis and treatment of drug-induced liver injury. Hepatol. Int. 2017, 11, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, C.; Zhang, W.; Xia, Q.; Ma, J.; He, X.; He, Y.; Fu, P.P.; Jia, W.; Zhuge, Y.; et al. Developing urinary pyrrole-amino acid adducts as non-invasive biomarkers for identifying pyrrolizidine alkaloids-induced liver injury in human. Arch. Toxicol. 2021, 95, 3191–3204. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, W.; Ma, J.; Xia, Q.; Song, Z.; Zhu, L.; Zhang, C.; Liu, J.; Ye, Y.; Fu, P.P.; et al. Blood pyrrole–DNA adducts define the early tumorigenic risk in patients with pyrrolizidine alkaloid-induced liver injury. Environ. Sci. Technol. Lett. 2021, 8, 551–557. [Google Scholar] [CrossRef]
- Teschke, R.; Vongdala, N.; Quan, N.V.; Quy, T.N.; Xuan, T.D. Metabolic toxification of 1,2-unsaturated pyrrolizidine alkaloids causes human hepatic sinusoidal obstruction syndrome: The update. Int. J. Mol. Sci. 2021, 22, 10419. [Google Scholar] [CrossRef]
- Chang, F.; Yu, D.; Wang, H.; Qu, S.; Wang, D.; Liu, X.; Pan, Y. Authentication of Saposhnikovia divaricata (Trucz.) Schischk and its two adulterants based on their macroscopic morphology and microscopic characteristics. Microsc. Res. Tech. 2021, 84, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Li, N.; Xia, Q.; Fu, P.P.; Peng, S.; Ye, Y.; Lin, G. Characteristic ion clusters as determinants for the identification of pyrrolizidine alkaloid N-oxides in pyrrolizidine alkaloid-containing natural products using HPLC-MS analysis. J. Mass Spectrom. 2012, 47, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.C.; Huxtable, R.J. Quantitation of the hepatic release of metabolites of the pyrrolizidine alkaloid, monocrotaline. Toxicol. Appl. Pharm. 1994, 127, 58–63. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose of Herbal Extract (g/kg) | Equivalent to PA Dose (mmol/kg) | Pyrrole–Protein Adducts (nmol/g Liver) | ALT (SF unit/mL) | GSH (µmol/g Liver) | Animal Group |
|---|---|---|---|---|---|
| Different batches of control groups | |||||
| 0 | 0 | 0 | 50.19 ± 2.21 | 4.25 ± 0.23 | A |
| 0 | 0 | 0 | 43.00 ± 2.89 | 7.70 ± 0.52 | B |
| 0 | 0 | 0 | 40.87 ± 1.28 | 5.30 ± 0.31 | C |
| 0 | 0 | 0 | 47.32 ± 1.96 | 4.15 ± 0.23 | D |
| Crotalaria sessiliflora water extract | |||||
| 0.50 | 0.32 | 6.46 ± 1.90 | 56.04 ± 3.12 | 7.91 ± 0.40 | B |
| 0.60 | 0.38 | 13.48 ± 2.11 | 57.04 ± 4.51 | 10.37 ± 0.42 ** | B |
| 0.62 | 0.40 | 17.78 ± 2.87 | 57.75 ± 6.26 | 6.05 ± 0.46 | C |
| 0.78 | 0.50 | 14.71 ± 1.59 | 56.64 ± 7.55 | 7.36 ± 0.16 ** | C |
| 0.80 | 0.51 | 16.90 ± 1.71 | 58.64 ± 8.52 | 11.14 ± 0.19 ** | B |
| 0.94 | 0.60 | 25.57 ± 2.64 | 57.54 ± 6.95 | 7.91 ± 0.26 ** | C |
| 1.00 | 0.64 | 20.74 ± 1.66 | 54.57 ± 6.05 | 8.85 ± 0.89 ** | A |
| 1.09 | 0.70 | 28.59 ± 0.91 | 71.81 ± 5.35 ** | 7.73 ± 0.43 ** | C |
| 1.20 | 0.77 | 26.97 ± 0.71 | 75.63 ± 4.30 * | 9.85 ± 0.34 ** | A |
| 1.50 | 0.96 | 32.96 ± 2.35 | 88.59 ± 13.69 ** | 8.66 ± 0.51 ** | A |
| Monocrotaline | |||||
| 0.80 | 29.55 ± 1.38 | 81.73 ± 11.04 * | 7.52 ± 0.59 * | A | |
| Gynura japonica water extract | |||||
| 3 | 0.08 | 7.44 ± 1.05 | 52.36 ± 1.70 | 3.99 ± 0.37 | D |
| 6 | 0.16 | 16.24 ± 2.09 | 55.67 ± 3.28 | 5.66 ± 0.30 | D |
| 12 | 0.31 | 28.10 ± 3.11 | 61.98 ± 5.36 | 6.75 ± 0.74 ** | D |
| 15 | 0.39 | 30.88 ± 0.89 | 68.29 ± 7.68 | 10.19 ± 0.36 ** | D |
| 18 | 0.47 | 39.23 ± 1.90 | 83.34 ± 18.34 * | 6.24 ± 0.64 ** | D |
| Dose of Crotalaria sessiliflora Water Extract (g/kg) | Equivalent to Monocrotaline Dose (mmol/kg) | Hemorrhage | Coagulative Necrosis |
|---|---|---|---|
| 0.00 | 0.00 | - | - |
| 0.50 | 0.32 | - | + |
| 0.60 | 0.38 | - | + |
| 0.62 | 0.40 | - | + |
| 0.78 | 0.50 | + | ++ |
| 0.80 | 0.51 | + | ++ |
| 0.94 | 0.60 | ++ | +++ |
| 1.00 | 0.64 | ++ | +++ |
| 1.09 | 0.70 | +++ | +++ |
| 1.20 | 0.77 | +++ | +++ |
| MCT * | 0.80 | +++ | +++ |
| 1.50 | 0.96 | +++ | +++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Li, M.; Li, N.; Chan, W.Y.; Lin, G. Pyrrolizidine Alkaloid-Induced Hepatotoxicity Associated with the Formation of Reactive Metabolite-Derived Pyrrole–Protein Adducts. Toxins 2021, 13, 723. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100723
Ma J, Li M, Li N, Chan WY, Lin G. Pyrrolizidine Alkaloid-Induced Hepatotoxicity Associated with the Formation of Reactive Metabolite-Derived Pyrrole–Protein Adducts. Toxins. 2021; 13(10):723. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100723
Chicago/Turabian StyleMa, Jiang, Mi Li, Na Li, Wood Yee Chan, and Ge Lin. 2021. "Pyrrolizidine Alkaloid-Induced Hepatotoxicity Associated with the Formation of Reactive Metabolite-Derived Pyrrole–Protein Adducts" Toxins 13, no. 10: 723. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100723