Occurrence and Characterization of Penicillium Species Isolated from Post-Harvest Apples in Lebanon

 , , , , , , ,
, , , , , , ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Frequency of Post-Harvest Fungal Pathogens
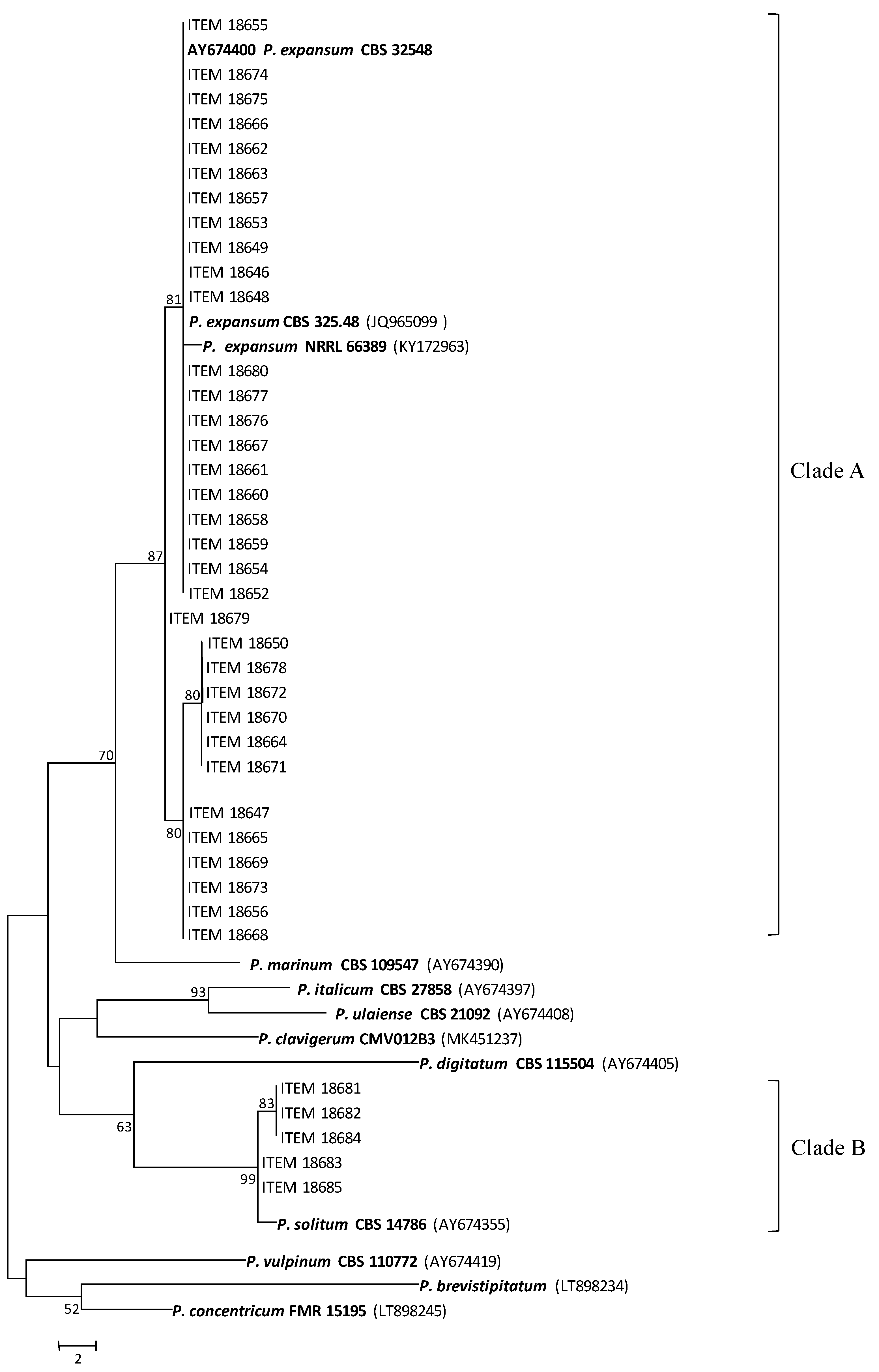
2.2. Identification of Penicillium Isolates
2.3. Patulin Production In Vitro
3. Discussion
4. Materials and Methods
4.1. Apple Sampling
4.2. Isolation of Fungi
4.3. Characterization of Penicillium Isolates
4.3.1. Morphological Characterization
4.3.2. Molecular Identification
4.4. Patulin Determination In Vitro
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bondonno, N.P.; Bondonno, C.P.; Ward, N.C.; Hodgson, J.M.; Croft, K.D. The cardiovascular health benefits of apples: Whole fruit vs. isolated compounds. Trends Food Sci. Technol. 2017, 69, 243–256. [Google Scholar] [CrossRef]
- Hyson, D.A. A comprehensive review of apples and apple components and their relationship to human health. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef]
- Xiao, C.L.; Kim, Y.K. Postharvest fruit rots in apples caused by Botrytis cinerea, Phacidiopycnis washingtonensis, and Sphaeropsis pyriputrescens. Plant Health Prog. 2008, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Belete, T.; Boyraz, N. Critical review on apple scab (Venturia inaequalis) biology, epidemiology, economic importance, management and defense mechanisms to the causal agent. J. Plant Physiol. Pathol. 2017, 5, 2–11. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, G.-L.; Li, B.-H.; Xu, X.-M.; Dong, X.-L.; Wang, C.-X.; Li, G.-F. Seasonal dynamics of Botryosphaeria dothidea infections and symptom development on apple fruits and shoots in China. Eur. J. Plant Pathol. 2016, 146, 507–518. [Google Scholar] [CrossRef]
- Michailides, T.J.; Spotts, R.A. Postharvest diseases of pome and stone fruits caused by Mucor piriformis in the Pacific Northwest and California. Plant Dis. 1990, 74, 537–543. [Google Scholar] [CrossRef]
- Xiao, C.L.; Rogers, J.D.; Boal, R.J. First report of a new postharvest fruit rot on apple caused by Sphaeropsis pyriputrescens. Plant Dis. 2004, 88, 223. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.L.; Rogers, J.; Kim, Y.; Liu, Q. Phacidiopycnis washingtonensis a new species associated with pome fruits from Washington State. Mycologia 2005, 97, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Louw, J.; Korsten, L. Pathogenic Penicillium spp. on apple and pear. Plant Dis. 2014, 98, 590–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberger, D.A. Blue mold. In Compendium of Apple and Pear Diseases; Jones, A.L., Aldwinckle, H.S., Eds.; APS Press: Minneapolis, MI, USA, 1990; pp. 54–55. [Google Scholar]
- Sanzani, S.; Susca, A.; Mastrorosa, S.; Solfrizzo, M. Patulin risk associated with blue mould of pome fruit marketed in southern Italy. Qual. Assur. Saf. Crop. Foods 2017, 9, 23–29. [Google Scholar] [CrossRef]
- Vico, I.; Gaskins, V.; Duduk, N.; Vasić, M.; Yu, J.J.; Peter, K.A.; Jurick, W.M. First report of Penicillium crustosum causing blue mold on stored apple fruit in Serbia. Plant Dis. 2014, 98, 1430. [Google Scholar] [CrossRef]
- Andersen, B.; Smedsgaard, A.J.; Frisvad, J.C. Penicillium expansum: Consistent production of patulin, chaetoglobosin, and other secondary metabolites in culture and their natural occurrence in fruit products. J. Agric. Food Chem. 2004, 52, 2421–2428. [Google Scholar] [CrossRef]
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [Green Version]
- Cheraghali, A.M.; Mohammadi, H.; Amirahmadi, M.; Yazdanpanah, H.; Abouhossain, G.; Zamanian, F.; Ghazi-Khansari, M.; Afshar, M. Incidence of patulin contamination in apple juice produced in Iran. Food Control 2005, 16, 165–167. [Google Scholar] [CrossRef]
- Sant’Ana, A.D.S.; Rosenthal, A.; De Massaguer, P.R. The fate of patulin in apple juice processing: A review. Food Res. Int. 2008, 41, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Hammami, W.; Al-Thani, R.; Fiori, S.; Al-Meer, S.; Atia, F.A.; Rabah, D.; Migheli, Q.; Jaoua, S. Patulin and patulin producing Penicillium spp. occurrence in apples and apple-based products including baby food. J. Infect. Dev. Ctries. 2017, 11, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 28 August 2021).
- USAID. Pome Fruit Value Chain Assessment Report—Lebanon Industry Value Chain Development (LIVCD) Project. United States Agency for International Development, Washington; 2013. Available online: http://pdf.usaid.gov/pdf_docs/PA00JWQH.pdf (accessed on 15 December 2020).
- Letters, B. Export Value Chain Analysis–Fresh Fruit and Vegetables Lebanon. 2018. Available online: http://www.bureauleeters.nl/data/261-Kws0xYbtYkfr/vca-ffv-lebanon-2018.pdf (accessed on 28 August 2021).
- Tarabay, P.A.; Chahine, H.; Tawk, S.T.; Nemer, N.; Habib, W. Reduction of food losses in Lebanese apple through good harvesting and postharvest practices. Ann. Agric. Sci. 2018, 63, 207–213. [Google Scholar] [CrossRef]
- FAO. The State of Food and Agriculture 2019. Moving Forward on Food Loss and Waste Reduction; FAO: Rome, Italy, 2019. [Google Scholar]
- Konstantinou, S.; Karaoglanidis, G.S.; Bardas, G.A.; Minas, I.; Doukas, E.; Markoglou, A.N. Postharvest fruit rots of apple in Greece: Pathogen incidence and relationships between fruit quality parameters, cultivar susceptibility, and patulin production. Plant Dis. 2011, 95, 666–672. [Google Scholar] [CrossRef] [Green Version]
- Sholberg, P.; Haag, P. Incidence of postharvest pathogens of stored apples in British Columbia. Can. J. Plant Pathol. 1996, 18, 81–85. [Google Scholar] [CrossRef]
- Valiuškaitė, A.; Kviklienė, N.; Kviklys, D.; Lanauskas, J. Post-harvest fruit rot incidence depending on apple maturity. Agron. Res. 2006, 4, 427–431. [Google Scholar]
- Guo, Z.; Guo, C.; Sun, L.; Zuo, M.; Chen, Q.; El-Seedi, H.; Xiaobo, Z. Identification of the apple spoilage causative fungi and prediction of the spoilage degree using electronic nose. J. Food Proc. Eng. 2021, 44, e13856. [Google Scholar] [CrossRef]
- Wenneker, M.; Köhl, J. Postharvest decay of apples and pear in the Netherlands. Acta Hortic. 2014, 1053, 107–112. [Google Scholar] [CrossRef]
- Narayanasamy, P. Postharvest Pathogen and Disease Management; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.; Hong, S.-B.; Klaassen, C.; Perrone, G.; Seifert, K.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smiri, M.; Kheireddine, A.; Hammami, R.; Rouissi, M.; Espeso, E.A.; Sadfi-Zouaoui, N. An Assessment of the air quality in apple warehouses: New record of Aspergillus europaeus, Aspergillus pulverulentus, Penicillium allii and Penicillium sumatraense as decay agents. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Vidal, A.; Ouhibi, S.; Ghali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. The mycotoxin patulin: An updated short review on occurrence, toxicity and analytical challenges. Food Chem. Toxicol. 2019, 129, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Morales, H.; Sanchis, V.; Rovira, A.; Ramos, A.J.; Marin, S. Patulin accumulation in apples during postharvest: Effect of controlled atmospheres and fungicide treatments. Food Control 2007, 18, 1443–1448. [Google Scholar] [CrossRef]
- Frisvad, J. A critical review of producers of small lactone mycotoxins: Patulin, penicillic acid and moniliformin. World Mycotoxin J. 2018, 11, 73–100. [Google Scholar] [CrossRef]
- Sholberg, P.L.; Bedford, K.; Stokes, S. Sensitivity of Penicillium spp. and Botrytis cinerea to pyrimethanil and its control of blue and gray mold of stored apples. Crop. Prot. 2005, 24, 127–134. [Google Scholar] [CrossRef]
- Wu, G.; Ii, W.M.J.; Lichtner, F.; Peng, H.; Yin, G.; Gaskins, V.L.; Yin, Y.; Hua, S.-S.; Peter, K.A.; Bennett, J.W. Whole-genome comparisons of Penicillium spp. reveals secondary metabolic gene clusters and candidate genes associated with fungal aggressiveness during apple fruit decay. PeerJ 2019, 7, e6170. [Google Scholar] [CrossRef] [Green Version]
- Marín, S.; Morales, H.; Hasan, H.A.H.; Ramos, A.J.; Sanchis, V. Patulin distribution in Fuji and Golden apples contaminated with Penicillium expansum. Food Addit. Contam. 2006, 23, 1316–1322. [Google Scholar] [CrossRef] [Green Version]
- Morales, H.; Barros, G.; Marin, S.; Chulze, S.; Ramos, A.J.; Sanchis, V. Effects of apple and pear varieties and pH on patulin accumulation by Penicillium expansum. J. Sci. Food Agric. 2008, 88, 2738–2743. [Google Scholar] [CrossRef]
- Spadaro, D.; Lorè, A.; Garibaldi, A.; Gullino, M.L. A new strain of Metschnikowia fructicola for postharvest control of Penicillium expansum and patulin accumulation on four cultivars of apple. Postharvest Biol. Technol. 2013, 75, 1–8. [Google Scholar] [CrossRef]
- Reddy, K.R.N.; Spadaro, D.; Lore, A.; Gullino, M.L.; Garibaldi, A. Potential of patulin production by Penicillium expansum strains on various fruits. Mycotoxin Res. 2010, 26, 257–265. [Google Scholar] [CrossRef]
- Amiri, A.; Bompeix, G. Diversity and population dynamics of Penicillium spp. on apples in pre- and postharvest environments: Consequences for decay development. Plant Pathol. 2005, 54, 74–81. [Google Scholar] [CrossRef]
- Morales, H.; Marín, S.; Ramos, A.J.; Sanchis, V. Influence of post-harvest technologies applied during cold storage of apples in Penicillium expansum growth and patulin accumulation: A review. Food Control 2010, 21, 953–962. [Google Scholar] [CrossRef]
- Sommer, N.; Fortlage, R.J.; Edwards, D.C. Postharvest diseases of selected commodities. In Postharvest Technology of Horticultural Crops, 3rd ed.; Kader, A., Ed.; University of California, Agricultural and Natural Resources: Davis, CA, USA, 2002. [Google Scholar]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarch, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From prevention in the field to detoxification by adsorption or transformation. Food Addit. Contam. Part A 2011, 28, 1590–1609. [Google Scholar] [CrossRef]
- Ammar, H.; Awny, N.M.; Fahmy, H.M. Influence of environmental conditions of atoxigenic Aspergillus flavus HFB1 on biocontrol of patulin produced by a novel apple contaminant isolate, A. terreus HAP1, in vivo and in vitro. Biocatal. Agric. Biotechnol. 2017, 12, 36–44. [Google Scholar] [CrossRef]
- Tang, H.; Peng, X.; Li, X.; Meng, X.; Liu, B. Biodegradation of mycotoxin patulin in apple juice by calcium carbonate immobilized porcine pancreatic lipase. Food Control 2018, 88, 69–74. [Google Scholar] [CrossRef]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; APS Press: Minneapolis, MI, USA, 1998. [Google Scholar]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium: A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–174. [Google Scholar]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi. (CBS Laboratory Manual Series); CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2010. [Google Scholar]
- Pitt, J.I. The Genus Penicillium and Its Teleomorphic States Eupenicillium and Talaromyces; Academic Press Inc.: London, UK, 1979. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Houbraken, J.; Wang, L.; Lee, H.B.; Frisvad, J. New sections in Penicillium containing novel species producing patulin, pyripyropens or other bioactive compounds. Persoonia 2016, 36, 299–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Facility | Location | ‘Red Delicious’ | ‘Golden Delicious’ | ||||
|---|---|---|---|---|---|---|---|
| Decayed Fruits (No.) | Blue Mould (%) | Gray Mould (%) | Decayed Fruits (No.) | Blue Mould (%) | Gray Mould (%) | ||
| 1 | Mayrouba | 38 | 89.5 | 10.5 | 24 | 100 | 0 |
| 2 | Hrajel | 39 | 74.4 | 48.7 | 30 | 86.7 | 26.7 |
| 3 | Hrajel | 37 | 83.8 | 10.8 | 33 | 87.9 | 15.2 |
| 4 | Kfardebian | 40 | 67.5 | 50.0 | 33 | 69.7 | 54.5 |
| 5 | Dbaye | 8 | 37.5 | 75.0 | 24 | 58.3 | 58.3 |
| Total | 162 | 76.5 | 32.7 | 144 | 80.6 | 31.3 | |
| Morphological Characteristics | P. expansum | P. solitum |
|---|---|---|
| Characteristics on CYA (7 days) | ||
| Colony diameter at 25 °C (mm) | 29–47 (x = 37.9) a | 21–29 (x = 26.1) a |
| Colony diameter at 30 °C (mm) | 6–13 (x = 10) | 0–5.5 (x = 2.6) |
| Colony diameter at 37 °C (mm) | 0 | 0 |
| Color obverse | Dull green to green | Blue green to dark green with white margin |
| Color reverse | Orange brown to brown | Light beige |
| Exudate presence | None | None |
| Characteristics on MEA (7 days at 25 °C) | ||
| Colony diameter (mm) | 27–39 (x = 34.4) | 22–25 (x = 23.3) |
| Colony texture | Fasciculate | Velutinous, floccose in center |
| Color colony obverse | Dull green to green | Blue green to dark green |
| Conidia shape | Ellipsoid | Globose to sub-globose |
| Conidia Length × width (µm) | 4.2–4.4 × 3.2–3.4 (x = 4.3 × 3.3) | 4.6–5.0 × 4.3–4.8 (x = 5.0 × 4.7) |
| Conidia ornamentation (wall) | Smooth | Smooth |
| Conidia color | Green | Green |
| Stipe ornamentation | Smooth | Rough |
| Conidiophore pattern | Terverticillate | Terverticillate |
| Characteristics on PDA (30 days at 25 °C) | ||
| Synnemata presence | Often present | None |
| Penicillium Species | Strain Number | Origin | Accession Number | Patulin (mg L−1) a | |
|---|---|---|---|---|---|
| Town | Apple Cultivar | ||||
| P. expansum | ITEM 18646 | Mayrouba | Red Delicious | FR997369 | 95.2 |
| ITEM 18647 | Mayrouba | Red Delicious | FR997367 | n.d. | |
| ITEM 18648 | Mayrouba | Red Delicious | FR997368 | 46.2 | |
| ITEM 18649 | Mayrouba | Golden Delicious | FR997370 | 125.9 | |
| ITEM 18650 | Mayrouba | Golden Delicious | FR997371 | 40.6 | |
| ITEM 18652 | Hrajel | Red Delicious | FR997374 | <LOQ | |
| ITEM 18653 | Hrajel | Red Delicious | FR997375 | n.d. | |
| ITEM 18654 | Hrajel | Red Delicious | FR997376 | n.d. | |
| ITEM 18655 | Hrajel | Red Delicious | FR997372 | 10.7 | |
| ITEM 18656 | Hrajel | Red Delicious | FR997373 | 101.0 | |
| ITEM 18657 | Hrajel | Golden Delicious | FR997378 | n.d. | |
| ITEM 18658 | Hrajel | Golden Delicious | FR997379 | n.d. | |
| ITEM 18659 | Hrajel | Golden Delicious | FR997377 | n.d. | |
| ITEM 18660 | Hrajel | Red Delicious | FR997381 | n.d. | |
| ITEM 18661 | Hrajel | Red Delicious | FR997383 | n.d. | |
| ITEM 18662 | Hrajel | Red Delicious | FR997384 | 34.7 | |
| ITEM 18663 | Hrajel | Red Delicious | FR997380 | <LOQ | |
| ITEM 18664 | Hrajel | Red Delicious | FR997382 | 24.8 | |
| ITEM 18665 | Hrajel | Golden Delicious | FR997385 | 75.3 | |
| ITEM 18666 | Hrajel | Golden Delicious | FR997386 | n.d. | |
| ITEM 18667 | Hrajel | Golden Delicious | FR997387 | 13.5 | |
| ITEM 18668 | Hrajel | Golden Delicious | FR997388 | 47.9 | |
| ITEM 18669 | Kfardebian | Red Delicious | FR997389 | 83.6 | |
| ITEM 18670 | Kfardebian | Red Delicious | FR997390 | 16.3 | |
| ITEM 18671 | Kfardebian | Red Delicious | FR997391 | n.d. | |
| ITEM 18672 | Kfardebian | Golden Delicious | FR997394 | 91.7 | |
| ITEM 18673 | Kfardebian | Golden Delicious | FR997395 | 26.4 | |
| ITEM 18674 | Kfardebian | Golden Delicious | FR997396 | 19.7 | |
| ITEM 18675 | Kfardebian | Golden Delicious | FR997392 | 23.8 | |
| ITEM 18676 | Kfardebian | Golden Delicious | FR997393 | n.d. | |
| ITEM 18677 | Dbaye | Red Delicious | FR997397 | <LOQ | |
| ITEM 18678 | Dbaye | Red Delicious | FR997398 | 31.3 | |
| ITEM 18679 | Dbaye | Golden Delicious | FR997400 | 13.8 | |
| ITEM 18680 | Dbaye | Golden Delicious | FR997399 | <LOQ | |
| P. solitum | ITEM 18681 | Mayrouba | Red Delicious | FR997401 | n.d. |
| ITEM 18682 | Hrajel | Red Delicious | FR997402 | n.d. | |
| ITEM 18683 | Hrajel | Golden Delicious | FR997403 | n.d. | |
| ITEM 18684 | Hrajel | Golden Delicious | FR997405 | n.d. | |
| ITEM 18685 | Hrajel | Golden Delicious | FR997404 | n.d. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habib, W.; Masiello, M.; Chahine-Tsouvalakis, H.; Al Moussawi, Z.; Saab, C.; Tawk, S.T.; Piemontese, L.; Solfrizzo, M.; Logrieco, A.F.; Moretti, A.; et al. Occurrence and Characterization of Penicillium Species Isolated from Post-Harvest Apples in Lebanon. Toxins 2021, 13, 730. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100730
Habib W, Masiello M, Chahine-Tsouvalakis H, Al Moussawi Z, Saab C, Tawk ST, Piemontese L, Solfrizzo M, Logrieco AF, Moretti A, et al. Occurrence and Characterization of Penicillium Species Isolated from Post-Harvest Apples in Lebanon. Toxins. 2021; 13(10):730. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100730
Chicago/Turabian StyleHabib, Wassim, Mario Masiello, Hala Chahine-Tsouvalakis, Zahraa Al Moussawi, Carine Saab, Salwa Tohmé Tawk, Luca Piemontese, Michele Solfrizzo, Antonio Francesco Logrieco, Antonio Moretti, and et al. 2021. "Occurrence and Characterization of Penicillium Species Isolated from Post-Harvest Apples in Lebanon" Toxins 13, no. 10: 730. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100730