Comprehensive Analysis and Biological Characterization of Venom Components from Solitary Scoliid Wasp Campsomeriella annulata annulata

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Comprehensive Analysis of Venom Extract from Campsomeriella annulata annulata
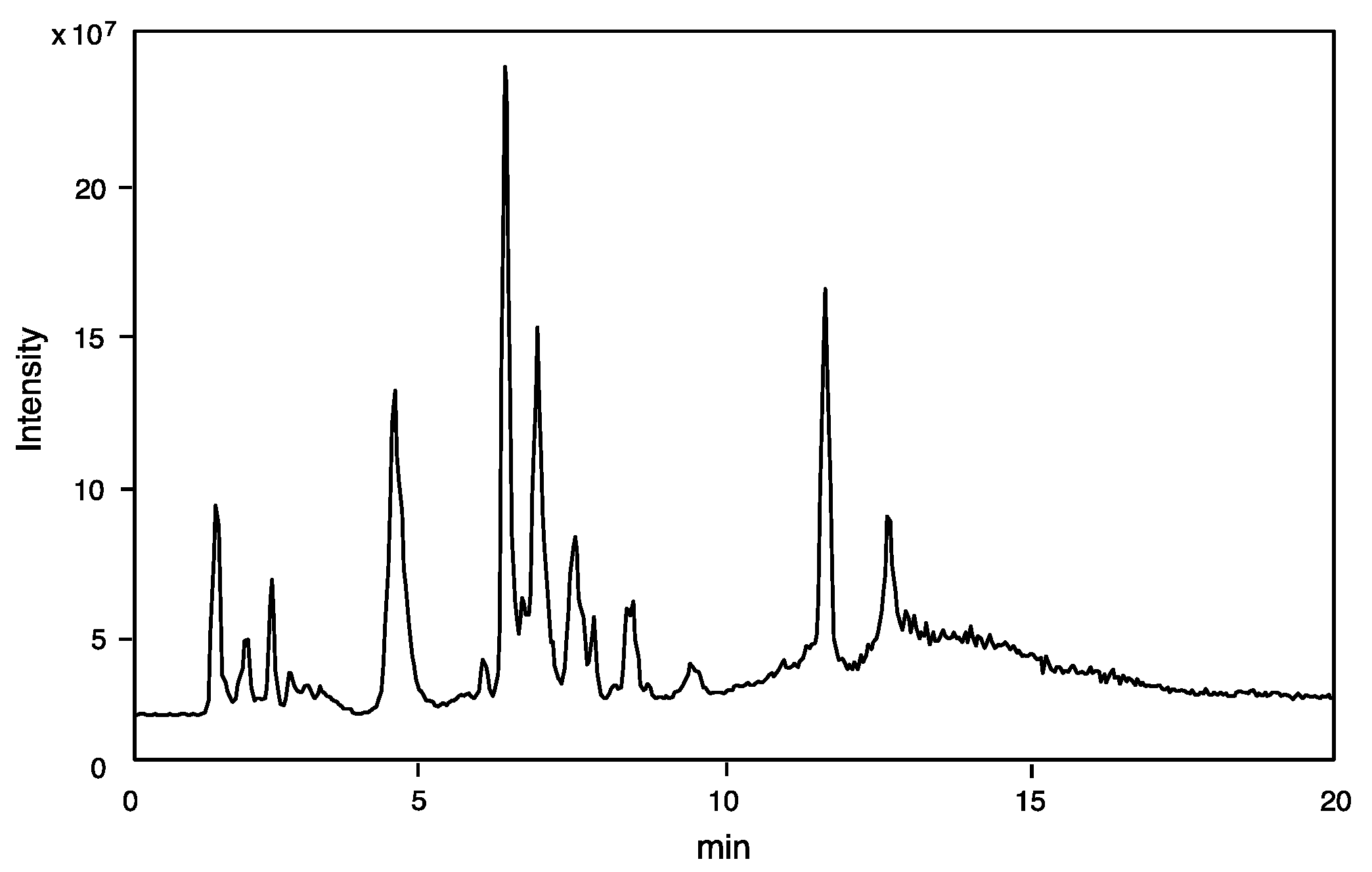
2.1.1. On-Line Mass Fingerprinting by LC-MS
2.1.2. Identification of Small Molecules (Amino Acids, Biogenic Amines and Nucleic Acids)
2.1.3. Peptide Sequencing by MS/MS Analysis
2.2. Biological Characterization of α-Campsomerin and β-Campsomerin
2.2.1. Interaction with NEP and ACE
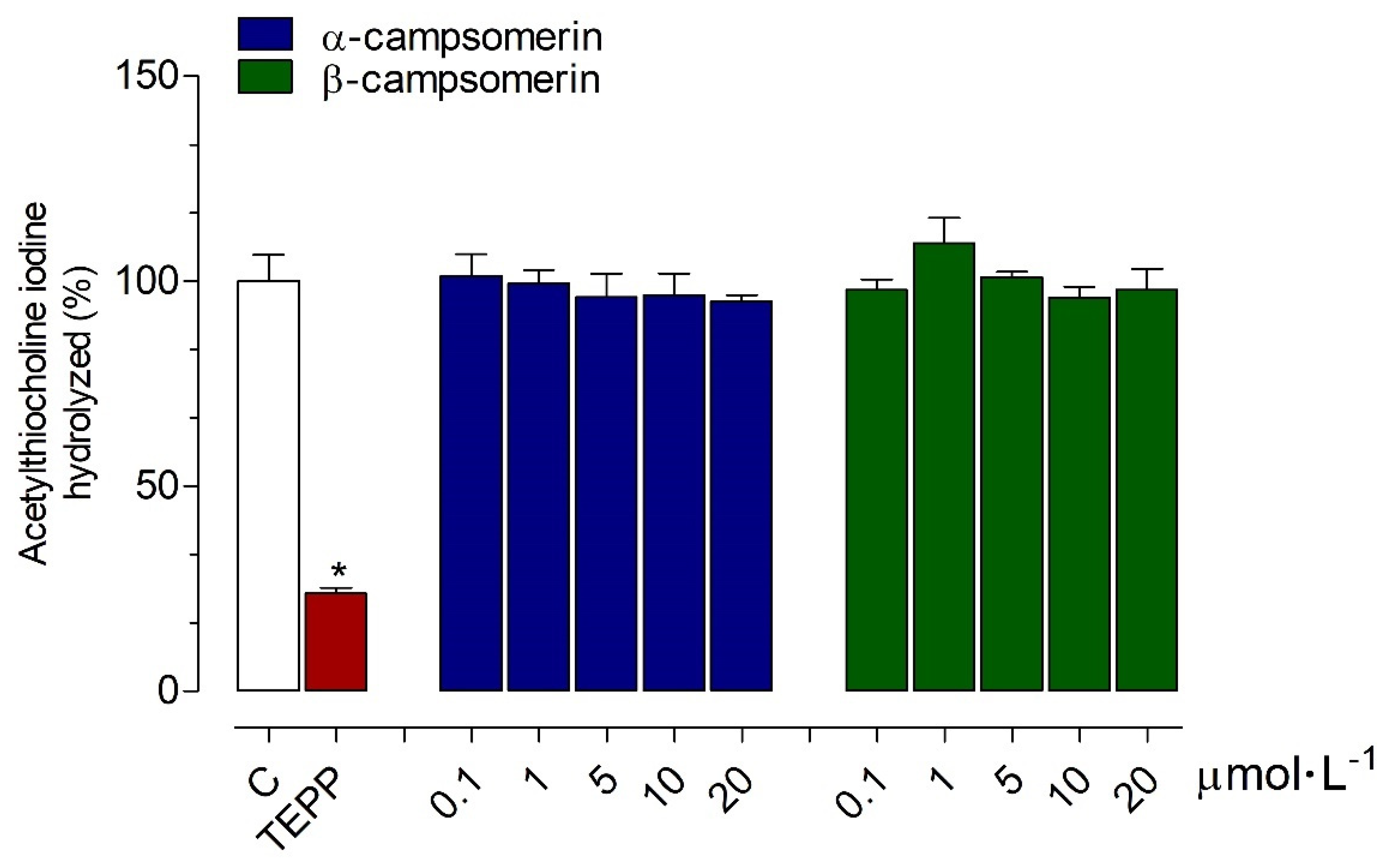
2.2.2. AChE Activity
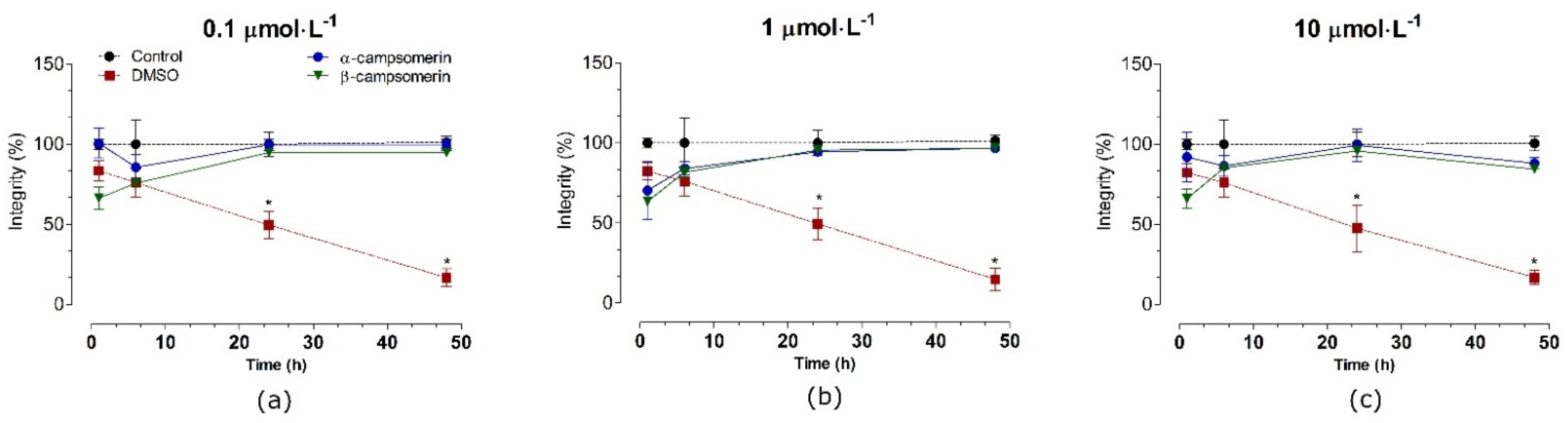
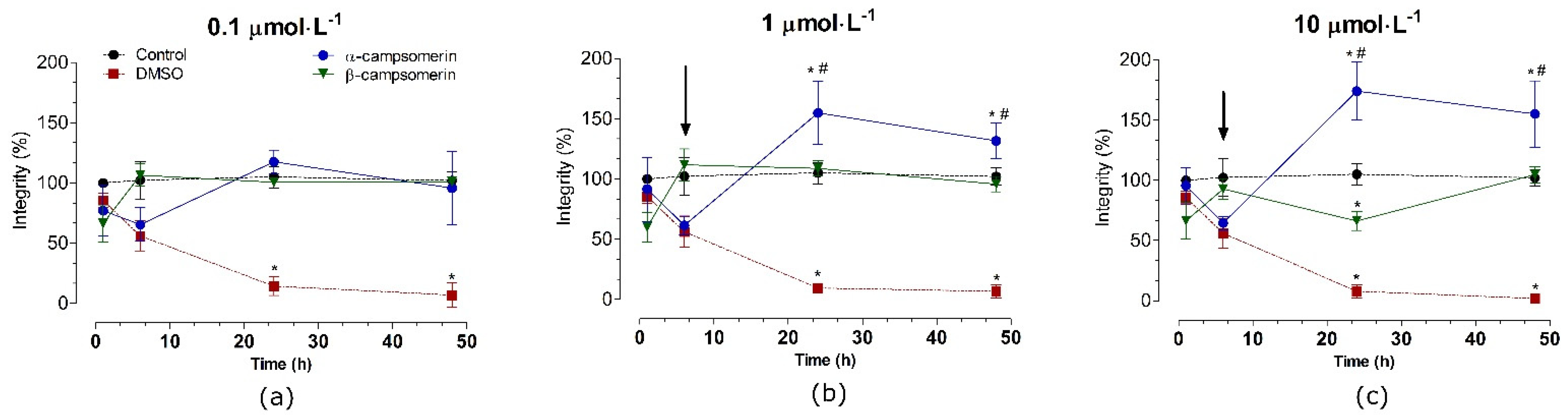
2.2.3. Cytotoxic Effects
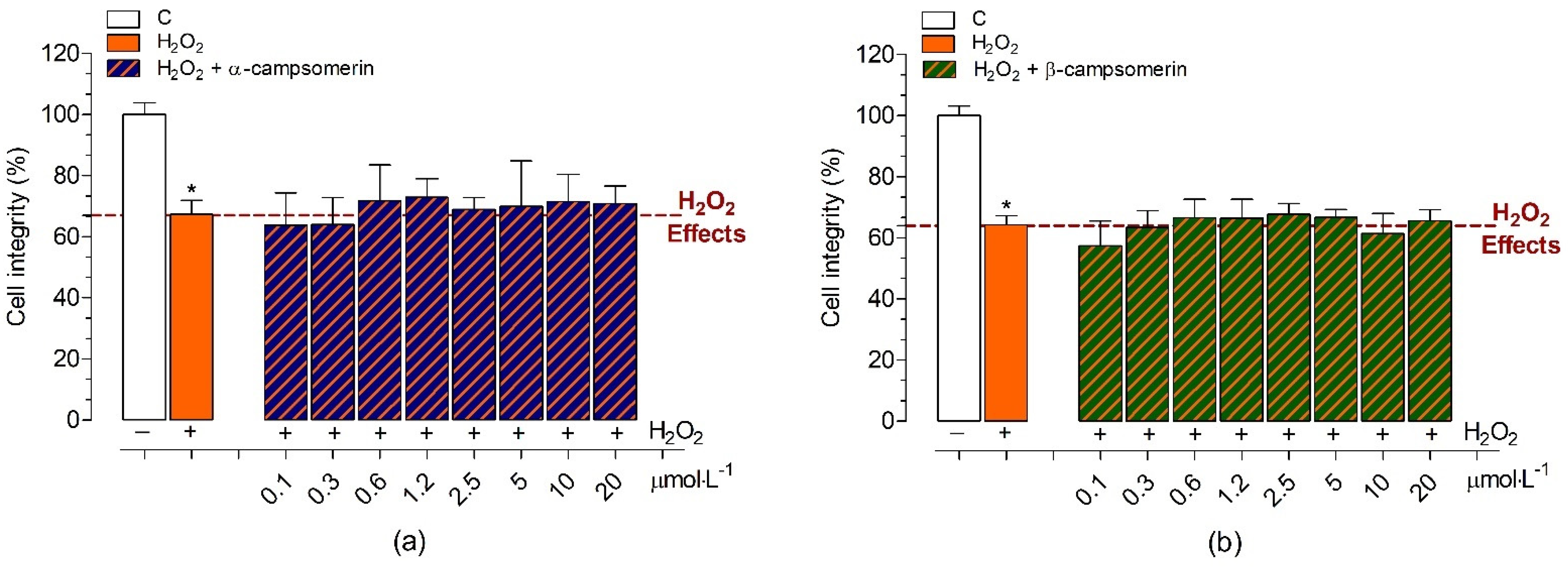
2.2.4. Neuroprotective Effects
2.3. Antimicrobial, Hemolytic, and Histamine-Releasing Activities of Annulatin and Related Peptides
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Wasp Collection
5.3. Cell Lines
5.4. LC-ESI-MS
5.5. MALDI-TOF MS
5.6. Peptide Synthesis
5.7. ACE and NEP Activities
5.8. Stability Tests of Peptides
5.9. Measurement of AChE Activity
5.10. Toxicity Studies on the Integrity Cell
5.11. Neuroprotective Assay
5.12. Antimicrobial, Hemolytic, and Histamine Releasing Activities
5.13. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rádis-Baptista, G.; Konno, K. Arthropod Venom Components and Their Potential Usage; MDPI: Basel, Switzerland, 2020; ISBN 978-3-03928-540-2 (PBK)/978-03928-541-9 (PDF). Available online: https://0-www-mdpi-com.brum.beds.ac.uk/2072-6651/12/2/82/htm (accessed on 8 November 2021).
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Saez, N.J.; Herzig, V. Versatile spider venom peptides and their medical and agricultural applications. Toxicon 2019, 158, 109–126. [Google Scholar] [CrossRef]
- King, G. Tying pest insects in knots: The deployment of spider-venom-derived knottins as bioinsecticides. Pest Manag. Sci. 2019, 75, 2437–2445. [Google Scholar] [CrossRef]
- Cid-Uribe, J.I.; Veytia-Busheli, J.I.; Romero-Gutierres, T.; Ortiz, E.; Possani, L.D. Scorpion venomics: A 2019 overview. Exp. Rev. Proteom. 2020, 17, 67–83. [Google Scholar] [CrossRef]
- Silva, J.; Monge-Fuentes, V.; Gomes, F.; Lopes, K.; dos Anjos, L.; Campos, G.; Arena, C.; Biolochi, A.; Gonçalves, J.; Galante, P.; et al. Pharmacological alternatives for the treatment of neurodegenerative disorders: Wasp and bee venoms and their components as new neuroactive tools. Toxins 2015, 7, 3179–3209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Baek, J.H.; Yoon, K.A. Differential properties of venom peptides and proteins in solitary vs. social hunting wasps. Toxins 2016, 8, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, K.M. Solitary Wasps: Behavior and Natural History; Cornell University Press: Ithaca, NY, USA, 2001; ISBN 0-8014-3721-0. [Google Scholar]
- White, S.R.; Kadavakollu, S. Bradykinin in Hemipepsis ustulata: A novel method for safely milking wasps. Toxicon 2016, 117, 49–52. [Google Scholar] [CrossRef] [Green Version]
- Nolasco, M.; Biondi, I.; Pimenta, D.C.; Branco, A. Extraction and preliminary chemical characterization of the venom of the spider wasp Pepsis decorata (Hymenoptera: Pompilidae). Toxicon 2018, 150, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.L.; Arvidson, R.; Banks, C.; Urenda, J.L.; Duong, E.; Mohammed, H.; Adams, M.E. Ampulexins: A new family of peptides in venom of the emerald jewel wasp, Ampulex compressa. Biochemistry 2018, 57, 1907–1916. [Google Scholar] [CrossRef]
- Kotea, S.; Faktorb, J.; Dapica, I.; Mayordomoa, M.Y.; Kocikowskia, M.; Kagansky, A.; Goodletta, D.; Vojtesek, B.; Huppa, T.; Wilcockson, D.; et al. Analysis of venom sac constituents from the solitary, aculeate wasp Cerceris rybyensis. Toxicon 2019, 169, 1–4. [Google Scholar] [CrossRef]
- Huicab-Uribe, M.A.; Verdel-Aranda, K.; Martínez-Hernández, A.; Zamudio, F.Z.; Jiménez-Vargas, J.M.; Lara-Reyna, J. Molecular composition of the paralyzing venom of three solitary wasps (Hymenoptera: Pompilidae) collected in southeast Mexico. Toxicon 2019, 168, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.; Walker, A.A.; Nguyen, S.H.; Jin, A.-H.; Deuis, J.R.; Vetter, I.; King, G.F.; Schmidt, J.O.; Robinson, S.D. Venom chemistry underlying the painful stings of velvet ants (Hymenoptera: Mutillidae). Cell. Mol. Life Sci. 2021, 78, 5163–5177. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Itagaki, Y.; Naoki, H.; Kawai, N.; Miwa, A.; Yasuhara, T.; Takayama, H. Isolation and structure of pompilidotoxins (PMTXs), novel neurotoxins in solitary wasp venoms. Biochem. Biophys. Res. Commun. 1998, 250, 612–616. [Google Scholar] [CrossRef]
- Kawai, N.; Konno, K. Molecular determinants of two neurotoxins that regulate sodium current inactivation in rat hippocampus. Neurosci. Lett. 2004, 361, 44–46. [Google Scholar] [CrossRef]
- Schiavon, E.; Stevens, M.; Zaharenko, A.J.; Konno, K.; Tytgat, J.; Wanke, E. Voltage-gated sodium channels isoform-specific effects of pompilidotoxins. FEBS J. 2010, 277, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Hernández, C.; Konno, K.; Salceda, E.; Vega, R.; Zaharenko, A.J.; Soto, E. Sa12b peptide from solitary wasp inhibits ASIC currents in rat dorsal root ganglion neurons. Toxins 2019, 10, 585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konno, K.; Kazuma, K.; Rangel, M.; Stolarz-de-Oliveira, J.; Fontana, R.; Kawano, M.; Fuchino, H.; Hide, I.; Yasuhara, T.; Nakata, Y. New mastoparan peptides in the venom of the solitary eumenine wasp Eumenes micado. Toxins 2019, 11, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, M.P.D.S.; Rangel, M.; Ruggiero Neto, J.; Konno, K. Chemical and biological characteristics of antimicrobial α-helical peptides found in solitary wasp venoms and their interaction with model membranes. Toxins 2019, 11, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picolo, G.; Hisada, M.; Sciani, J.M.; Conceição, I.M.; Machado, M.; Oliveira, V.; de Melo, R.L.; Cury, Y.; Konno, K.; Hayashi, M.A.F. Bradykinin-related peptides from the venom of the solitary wasp Cyphononyx fulvognathus. Biochem. Pharmacol. 2010, 79, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Nihei, K.; Peigneur, S.; Tytgat, J.; Lange, A.B.; Konno, K. Isolation and characterization of FMRFamide-like peptides in the venoms of solitary sphecid wasps. Peptides 2021, 142, 170575. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Kazuma, K.; Nihei, K. Peptide toxins in solitary wasp venoms. Toxins 2016, 8, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuhara, T.; Mantel, P.; Nakajima, T.; Piek, T. Two kinins isolated from an extract of the venom reservoirs of the solitary wasp Megascolia flavifrons. Toxicon 1987, 25, 527–535. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B.; Mantel, P.; Nakajima, T.; Pelhate, M.; Yasuhara, T. Threonine6-bradykinin in the venom of the wasp Colpa interrupta (F.) presynaptically blocks nicotinic synaptic transmission in the insect CNS. Comp. Biochem. Physiol. 1990, 96, 157–162. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B.; Mony, L.; Nakajima, T.; Pelhate, M.; Yasuhara, T. Block of synaptic transmission in insect CNS by toxins from the venom of the WASP Megascolia flavifrons (FAB.). Comp. Biochem. Physiol. 1987, 87, 287–295. [Google Scholar] [CrossRef]
- Konno, K.; Palma, M.S.; Hitara, I.Y.; Juliano, M.A.; Juliano, L.; Yasuhara, T. Identification of bradykinins in solitary wasp venoms. Toxicon 2002, 40, 309–312. [Google Scholar] [CrossRef]
- Alberto-Silva, C.; Portaro, F.; Kodama, R.; Pantaleão, H.; Rangel, M.; Nihei, K.; Konno, K. Novel neuroprotective peptides in the venom of the solitary scoliid wasp Scolia decorata ventralis. J. Venom. Anim. Toxins Incl. Trop. Dis. 2021, 27, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Fontana, R.; Lorenzi, C.C.B.; Naoki, H.; Itagaki, Y.; Miwa, A.; Kawai, N.; Nakata, Y.; Yasuhara, T.; et al. Anoplin, a novel antimicrobial peptide from the venom of the solitary wasp Anoplius samariensis. Biochim. Biophys. Acta 2001, 1550, 70–80. [Google Scholar] [CrossRef]
- Konno, K.; Rangel, M.; Oliveira, J.S.; Cabrera, M.P.S.; Fontana, R.; Hirata, I.Y.; Nakata, Y.; Mori, K.; Kawano, M.; Fuchino, H.; et al. Decoralin, a novel linear cationic α-helical peptide from the venom of the solitary eumenine wasp Oreumenes decoratus. Peptides 2007, 28, 2320–2327. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Fontana, R.; Rangel, M.; Oliveira, J.S.; Cabrera, M.P.; Ruggiero Neto, J.; Hide, I.; et al. Eumenitin, a novel antimicrobial peptide from the venom of the solitary eumenine wasp Eumenes rubronotatus. Peptides 2006, 27, 2624–2631. [Google Scholar] [CrossRef] [PubMed]
- Rangel, M.; Cabrera, M.P.S.; Kazuma, K.; Ando, K.; Wang, X.; Kato, M.; Nihei, K.; Hirata, I.Y.; Cross, T.; Garcia, A.N.; et al. Chemical and biological characterization of four new antimicrobial and α-helical peptides from the venoms of two solitary eumenine wasps. Toxicon 2011, 57, 1081–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigeri, Y.; Horie, M.; Yoshida, T.; Hagihara, Y.; Imura, T.; Inagaki, H.; Haramoto, Y.; Ito, Y.; Asashima, M. Physicochemical and biological characterizations of Pxt peptides from amphibian (Xenopus tropicalis) skin. J. Biochem. 2016, 159, 619–629. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, T.; Yasuhara, T.; Yoshida, N.; Takemoto, Y.; Shinonaga, S.; Kano, R.; Yoshida, H. The pattern analysis of biologically active amines in some Hymenopteran venoms by high performance liquid chromatography. Med. Entomol. Zool. 1983, 34, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Arendse, L.; Danser, A.; Poglitsch, M.; Touyz, R.; Burnett, J.; Llorens-Cortes, C.; Ehlers, M.; Sturrock, E. Novel therapeutic approaches targeting the renin-angiotensin system and associated peptides in hypertension and heart failure. Pharmacol. Rev. 2019, 71, 539–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walczak-Nowicka, Ł.J.; Herbet, M. Acetylcholinesterase inhibitors in the treatment of neurodegenerative diseases and the role of acetylcholinesterase in their pathogenesis. Int. J. Mol. Sci. 2021, 22, 9290. [Google Scholar] [CrossRef]
- Prasasty, V.; Radifar, M.; Istyastono, E. Natural peptides in drug discovery targeting acetylcholinesterase. Molecules 2018, 23, 2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, J.; de Castro, A.; Soares, F.; da Cunha, E.; Ramalho, T. Future therapeutic perspectives into the Alzheimer’s disease targeting the oxidative stress hypothesis. Molecules 2019, 24, 4410. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, D.; Brecht, K.; Koplovitz, I.; Sweeney, R. Acetylcholinesterase inhibition: Does it explain the toxicity of organophosphorus compounds? Arch. Toxicol. 2006, 80, 756–760. [Google Scholar] [CrossRef]
- Aksenova, M.; Aksenov, M.; Mactutus, C.; Booze, R. Cell culture models of oxidative stress and injury in the central nervous system. Curr. Neurovasc. Res. 2005, 2, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Querobino, S.M.; Carrettiero, D.C.; Costa, M.S.; Alberto-Silva, C. Neuroprotective property of low molecular weight fraction from B. Jararaca snake venom in H2O2-induced cytotoxicity in cultured hippocampal cells. Toxicon 2017, 129, 134–143. [Google Scholar] [CrossRef]
- Emerit, J.; Edeas, M.; Bricaire, F. Neurodegenerative diseases and oxidative stress. Biomed. Pharmacother. 2004, 58, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Querobino, S.M.; Costa, M.S.; Alberto-Silva, C. Protective effects of distinct proline-rich oligopeptides from B. Jararaca snake venom against oxidative stress-induced neurotoxicity. Toxicon 2019, 167, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Bácskay, I.; Nemes, D.; Fenyvesi, F.; Váradi, J.; Vasvári, G.; Fehér, P.; Vecsernyés, M.; Ujhelyi, Z. Role of cytotoxicity experiments in pharmaceutical development. In Cytotoxicity; Çelik, T.A., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Yi, X.; Liu, M.; Luo, Q.; Zhuo, H.; Cao, H.; Wang, J.; Han, Y. Toxic effects of dimethyl sulfoxide on red blood cells, platelets, and vascular endothelial cells in vitro. FEBS Open Bio 2017, 7, 485–494. [Google Scholar] [CrossRef]
- den Brouw, B.O.; Ghezellou, P.; Casewell, N.; Ali, S.; Fathinia, B.; Fry, B.; Bos, N.; Ikonomopoulou, M. Pharmacological characterisation of Pseudocerastes and Eristicophis viper venoms reveal anticancer (Melanoma) properties and a potentially novel mode of fibrinogenolysis. Int. J. Mol. Sci. 2021, 22, 6896. [Google Scholar] [CrossRef] [PubMed]
- Rodeiro, I.; Hernández, I.; Herrera, J.; Riera, M.; Donato, M.; Tolosa, L.; González, K.; Ansoar, Y.; Gómez-Lechón, M.; Berghe, W.V.; et al. Assessment of the cytotoxic potential of an aqueous-ethanolic extract from Thalassia testudinum angiosperm marine grown in the Caribbean sea. J. Pharm. Pharmacol. 2018, 70, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.M.; Reis, P.V.; Pimenta, A.M.C. Antimicrobial peptides in spider venoms. In Spider Venoms; Gopalakrishnakone, P., Corzo, G.A., de Lima, M.E., Diego-García, E., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 361–377. ISBN 978-94-007-6389-0. [Google Scholar]
- Fratini, F.; Cilia, G.; Turchi, B.; Felicioli, A. Insects, arachnids and centipedes venom: A powerful weapon against bacteria. A literature review. Toxicon 2017, 130, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Harry, J.; Tilson, H.A.; Padilla, S.; Lassiter, T.L.; Hunter, D. Biochemical measurement of cholinesterase activity. In Neurodegeneration Methods and Protocols; Humana Press: Tortowa, NJ, USA, 2003; pp. 237–246. [Google Scholar]
- Feoktistova, M.; Geserick, P.; Leverkus, M. Crystal violet assay for determining viability of cultured cells. Cold Spring Harb. Protoc. 2016, 4, 343–346. [Google Scholar] [CrossRef]
- Inagaki, H.; Akagi, M.; Imai, H.T.; Taylor, R.W.; Kubo, T. Molecular cloning and biological characterization of novel antimicrobial peptides, pilosulin 3 and pilosulin 4, from a species of the Australian ant genus Myrmecia. Arch. Biochem. Biophys. 2004, 428, 170–178. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fr. No. | Retention Time (min) | [M + H]+ m/z |
|---|---|---|
| 1 | 0.8–1.5 | 90.054, 112.086, 116.069, 118.085, 120.064, 146.164, 147.075, 148.059, 154.085, 156.076, 175.118, 203.222, 258.108, 752.482 |
| 2 | 1.5–2.0 | 132.101, 138.090, 150.057, 182.080, 245.075, 253.116, 268.102, 284.097, 324.126, 348.067, 664.111 |
| 3 | 2.0–3.0 | 166.085, 269.086, 732.288, 873.461, 911.428, 916.637, 952.596, 1006.514, 1050.563, 1377.700, 1629.831 |
| 4 | 3.0–4.0 | 272.133, 660.387, 911.497, 953.580, 1164.582, 1435.706, 1487.628 |
| 5 | 4.0–5.0 | 205.095, 573.309, 585.367, 642.388, 1085.694, 1227.626, 2741.410, 2913.678, 2927.693, 2941.668, 2981.607, 3025.066 |
| 6 | 5.0–5.6 | 497.400, 771.408, 939.662, 1148.716, 1423.836, 2218.047 |
| 7 | 5.6–6.1 | 378.163, 487.319, 728.497, 1418.844, 1439.702, 1548.844, 5297.517, 6211.754 |
| 8 | 6.1–6.3 | 403.251, 555.356, 656.346, 1074.574, 1127.483, 1265.684, 1447.679, 1719.983 |
| 9 | 6.3–6.8 | 753.455, 969.590, 1105.569, 1534.865, 1587.776, 2068.808, 2229.234, 3251.858 |
| 10 | 6.8–7.1 | 419.237, 638.392, 1056.622, 1247.703, 1702.955 |
| 11 | 7.1–7.7 | 1060.438, 1378.768, 2208.189, 3015.057, 3251.857 |
| 12 | 7.7–8.3 | 918.474, 1499.748, 3152.777, 4089.223 |
| 13 | 8.3–8.6 | 900.542, 1128.687, 1182.607, 1250.760, 1266.756, 1500.731, 2244.169, 4716.118 |
| 14 | 8.6–9.0 | 813.492, 1072.654, 1258.713, 2913.497 |
| 15 | 9.0–10.0 | 1365.787, 1452.818, 1902.932, 2765.478 |
| 16 | 10.0–11.0 | 918.557, 995.474, 1281.875, 1415.896, 1840.016, 1893.933 |
| 17 | 11.0–11.8 | 1274.625, 1593.932, 1645.945, 1689.987, 3640.506 |
| 18 | 11.8–12.3 | 1761.009, 1991.029, 2043.940, 2091.104, 2291.182 |
| 19 | 12.3–12.7 | 1324.985, 1689.951 |
| 20 | 12.7–13.8 | 1317.740, 1518.891, 1856.998, 1909.904 |
| RT (min) | Intensity × 104 | [M + H]+ m/z | Elemental Composition | Iminium Ion m/z | Elemental Composition | Compound |
|---|---|---|---|---|---|---|
| 1.07 | 50 | 156.076 | C6H10N3O2 | — | Histidine | |
| 140 | 175.118 | C6H15N4O2 | — | Arginine | ||
| 1.28 | 24 | 90.054 | C3H7NO2 | — | Alanine | |
| 18 | 120.064 | C4H10NO3 | — | Threonine | ||
| 180 | 147.075 | C5H11N2O3 | — | Glutamine | ||
| 200 | 148.059 | C5H10NO4 | 102.054 | C4H8NO2 | Glutamic acid | |
| 1.34 | 700 | 116.069 | C5H10NO2 | 70.064 | C4H8N | Proline |
| 70 | 118.085 | C5H12NO2 | 72.080 | C4H10N | Valine | |
| 1.55 | 13 | 150.057 | C5H12NO2S | 104.052 | C4H10NS | Methionine |
| 1.68 | 180 | 132.101 | C6H14NO2 | 86.096 | C5H12N | L/I * |
| 1.72 | 80 | 182.080 | C9H12NO3 | — | Tyrosine | |
| 2.37 | 180 | 166.085 | C9H12NO2 | 120.080 | C8H10N | Phenylalanine |
| 4.17 | 13 | 205.096 | C11H13N2O2 | — | Tryptophan |
| RT (min) | Intensity × 104 | [M + H]+ m/z | Elemental Composition | Deamination m/z | Elemental Composition | Compound |
|---|---|---|---|---|---|---|
| 1.01 | 7 | 146.164 | C7H20N3 | — | Spermidine | |
| 1.07 | 12 | 112.086 | C5H10N3 | 95.060 | C5H7N2 | Histamine |
| 6 | 203.222 | C10H27N4 | — | Spermine | ||
| 1.34 | 12 | 154.086 | C8H12NO2 | 137.059 | C8H9O2 | Dopamine |
| 1.61 | 700 | 138.090 | C8H12NO | 121.064 | C8H9O | Tyramine |
| RT (min) | Intensity × 104 | [M + H]+ m/z | Elemental Composition | Compound |
|---|---|---|---|---|
| 1.21 | 10 | 244.091 | C9H14N3O5 | Cytidine |
| 1.28 | 180 | 258.108 | C10H16N3O5 | Thymidine |
| 1.61 | 100 | 348.068 | C10H15N5O7P | AMP |
| 18 | 664.111 | C21H28N7O14P2 | NAD | |
| 1.68 | 1800 | 268.102 | C10H14N5O4 | Adenosine |
| 1.72 | 8 | 245.075 | C9H13N2O6 | Uridine |
| 1.97 | 90 | 284.097 | C10H14N5O5 | Guanosine |
| Fr | RT | Intens. × 103 | MSMS m/z | Charge | (M + H)+ | Sequence |
|---|---|---|---|---|---|---|
| 1 | 1.23 | 70 | 251.499 | 3+ | 752.482 | SKLHRL-NH2 |
| 2 | 1.95 | 85 | 253.116 | + | 253.116 | SF |
| 3 | 2.09 | 91 | 459.905 | 3+ | 1377.700 | RGPRTYSHGHPL |
| 2.27 | 1500 | 318.204 | 3+ | 952.596 | SLSKLHRL-NH2 | |
| 2.53 | 8 | 304.481 | 3+ | 911.428 | TYSHGHPL | |
| 2.78 | 35 | 366.648 | 2+ | 732.288 | HNAEFD | |
| 4 | 3.06 | 8 | 318.532 | 3+ | 953.580 | SLSKLHRL |
| 3.34 | 72 | 330.697 | 2+ | 660.387 | LSEALK | |
| 3.55 | 330 | 456.253 | 2+ | 911.497 | PRLPRLT | |
| 3.66 | 750 | 479.240 | 3+ | 1435.706 | RDPRTYSHGHPL | |
| 3.80 | 1500 | 388.865 | 3+ | 1164.582 | PRTYSHGHPL | |
| 5 | 4.06 | 42 | 293.187 | 2+ | 585.367 | LSPLR |
| 4.56 | 350 | 287.159 | 2+ | 573.309 | RPPGF | |
| 4.99 | 60 | 321.698 | 2+ | 642.388 | GLSPLR | |
| 6 | 5.18 | 85 | 383.577 | 3+ | 1148.716 | VPSLKSLHRL-NH2 |
| 5.37 | 90 | 386.208 | 2+ | 771.408 | RPPGFTP | |
| 5.60 | 29 | 313.893 | 3+ | 939.662 | LVKQKVLL-NH2 | |
| 7 | 5.79 | 80 | 378.163 | + | 378.163 | DFP |
| 6.00 | 90 | 364.751 | 2+ | 728.495 | LKSLLVG-NH2 | |
| 8 | 6.07 | 8 | 403.251 | + | 403.251 | LTGL |
| 6.26 | 600 | 358.863 | 3+ | 1074.574 | RPPGFTPFR (Thr6-BK) | |
| 9 | 6.41 | 11 | 377.231 | 2+ | 753.455 | RLPGLTP |
| 6.45 | 62,000 | 512.294 | 3+ | 1534.865 | PRLRRLTGLSPLR | |
| 6.74 | 860 | 323.869 | 3+ | 969.590 | PATLPAPFR | |
| 10 | 6.84 | 210 | 416.572 | 3+ | 1247.703 | RLVKPVPFYE |
| 6.94 | 1200 | 352.879 | 3+ | 1056.622 | RLPGLTPFR | |
| 7.02 | 12,000 | 568.324 | 3+ | 1702.955 | PRLRRLTGLSPLRAP | |
| 11 | 7.63 | 85 | 530.723 | 2+ | 1060.438 | HNAEFDAAW |
| 7.67 | 390 | 689.887 | 2+ | 1378.768 | PRLRRLTGLSPL | |
| 12 | 7.80 | 52 | 459.741 | 2+ | 918.474 | RPPGFTPF |
| 13 | 8.57 | 2200 | 564.848 | 2+ | 1128.687 | LSEALKSLLVG-NH2 |
| 14 | 8.72 | 470 | 536.829 | 2+ | 1072.650 | LSEALKSLLV |
| 8.73 | 160 | 407.249 | 2+ | 813.490 | PATLPAPF | |
| 8.84 | 460 | 629.861 | 2+ | 1258.713 | LSEALKSLLVGE | |
| 15 | 9.07 | 45 | 455.934 | 3+ | 1365.787 | DALPRLLPAPFR |
| 9.17 | 31 | 484.945 | 3+ | 1452.818 | DALPRLLPGTPFR | |
| 9.70 | 135 | 634.982 | 3+ | 1902.932 | DTFGPLYDKLHQYLGH-NH2 | |
| 16 | 10.69 | 16 | 427.964 | 3+ | 1281.875 | FLLPLLKGLLVG-NH2 |
| 10.81 | 28 | 472.637 | 3+ | 1415.896 | GLLTDLRKFLLK-NH2 | |
| 10.95 | 16 | 459.783 | 2+ | 918.557 | GLVYLLQL | |
| 17 | 11.72 | 9 | 549.320 | 3+ | 1645.945 | DDGLLTDLRKFLLK-NH2 |
| 18 | 11.92 | 30 | 697.706 | 3+ | 2091.104 | HPDDDDDFLLPLLKGLLVG-NH2 |
| 12.01 | 1600 | 664.348 | 3+ | 1991.029 | DDDDDGLLTDLRKFLLK-NH2 | |
| 20 | 13.11 | 1050 | 619.669 | 3+ | 1856.992 | DDDDDFLLPLLKGLLVG-NH2 |
| RT | Intensity × 103 | (M + H)+ | Sequence |
|---|---|---|---|
| Bradykinin-related peptides | |||
| 4.56 | 350 | 573.309 | RPPGF |
| 5.37 | 90 | 771.408 | RPPGFTP |
| 7.80 | 52 | 918.474 | RPPGFTPF |
| 6.26 | 600 | 1074.574 | RPPGFTPFR (Thr6-BK) |
| 6.41 | 11 | 753.455 | RLPGLTP |
| 6.94 | 1200 | 1056.622 | RLPGLTPFR (Ca-1056) |
| 8.73 | 160 | 813.490 | PATLPAPF |
| 6.74 | 860 | 969.590 | PATLPAPFR (Ca-969) |
| 9.07 | 45 | 1365.787 | DALPRLLPAPFR (Ca-1365) |
| 9.17 | 31 | 1452.818 | DALPRLLPGTPFR (Ca-1452) |
| 3.55 | 330 | 911.497 | PRLRRLT |
| 7.67 | 390 | 1378.768 | PRLRRLTGLSPL |
| 6.07 | 8 | 403.251 | LTGL |
| 4.06 | 42 | 585.367 | LSPLR |
| 4.99 | 60 | 642.388 | GLSPLR |
| 6.45 | 62,000 | 1534.865 | PRLRRLTGLSPLR (α-campsomerin) |
| 7.02 | 12,000 | 1702.955 | PRLRRLTGLSPLRAP (β-campsomerin) |
| Linear α-helical peptides | |||
| 3.34 | 72 | 660.387 | LSEALK |
| 8.72 | 470 | 1072.650 | LSEALKSLLV |
| 6.00 | 90 | 728.495 | LKSLLVG-NH2 |
| 8.57 | 2200 | 1128.687 | LSEALKSLLVG-NH2 (annulatin) |
| 8.84 | 460 | 1258.713 | LSEALKSLLVGE |
| 10.69 | 16 | 1281.875 | FLLPLLKGLLVG-NH2 (Ca-1281) |
| 13.11 | 1050 | 1856.992 | DDDDDFLLPLLKGLLVG-NH2 |
| 11.92 | 30 | 2091.104 | HPDDDDDFLLPLLKGLLVG-NH2 |
| 10.81 | 28 | 1415.896 | GLLTDLRKFLLK-NH2 (Ca-1415) |
| 11.72 | 9 | 1645.945 | DDGLLTDLRKFLLK-NH2 |
| 12.01 | 1600 | 1991.029 | DDDDDGLLTDLRKFLLK-NH2 |
| Miscellaneous | |||
| 1.23 | 70 | 752.482 | SKLHRL-NH2 |
| 2.27 | 1500 | 952.596 | SLSKLHRL-NH2 |
| 3.06 | 8 | 953.580 | SLSKLHRL |
| 5.18 | 85 | 1148.716 | VPSLKSLHRL-NH2 |
| 2.78 | 35 | 732.288 | HNAEFD |
| 7.63 | 85 | 1060.438 | HNAEFDAAW |
| 2.53 | 8 | 911.428 | TYSHGHPL |
| 3.80 | 1500 | 1164.582 | PRTYSHGHPL |
| 2.09 | 91 | 1377.700 | RGPRTYSHGHPL |
| 3.66 | 750 | 1435.706 | RDPRTYSHGHPL |
| 1.95 | 85 | 253.116 | SF |
| 5.79 | 80 | 378.163 | DFP |
| 10.95 | 16 | 918.557 | GLVYLLQL |
| 5.60 | 29 | 939.662 | LVKQKVLL-NH2 |
| 6.84 | 210 | 1247.703 | RLVKPVPFYE |
| 9.70 | 135 | 1902.931 | DTFGPLYDKLHQYLGH-NH2 |
| Peptide | Sequence | References |
|---|---|---|
| Bradykinin (BK) | RPPGFSPFR | [21,27] |
| Thr6-Bradykinin (Thr6-BK) | RPPGFTPFR | [21,24,25,27] |
| Megascoliakinin | RPPGFTPFRKA | [24] |
| Cyphokinin | DTRPPGFTPFR | [21] |
| Fulvonin | SIVLRGKAPFR | [21] |
| Cd-146 | SETGNTVTVKGFSPLR | [21] |
| α-Scoliidine | DYVTVKGFSPLR | [28] |
| β-Scoliidine | DYVTVKGFSPLRKA | [28] |
| α-Campsomerin | PRLRRLTGLSPLR | This work |
| β-Campsomerin | PRLRRLTGLSPLRAP | This work |
| Peptide | Sequence | References |
|---|---|---|
| Anoplin | GLLKRIKTLL-NH2 | [29] |
| Decoralin | SLLSLIRKLIT | [30] |
| Eumenitin | LNLKGLFKKVKSLLT | [31] |
| Eumenitin-F | LNLKGLFKKVASLLT | [32] |
| Eumenitin-R | LNLKGLIKKVASLLN | [32] |
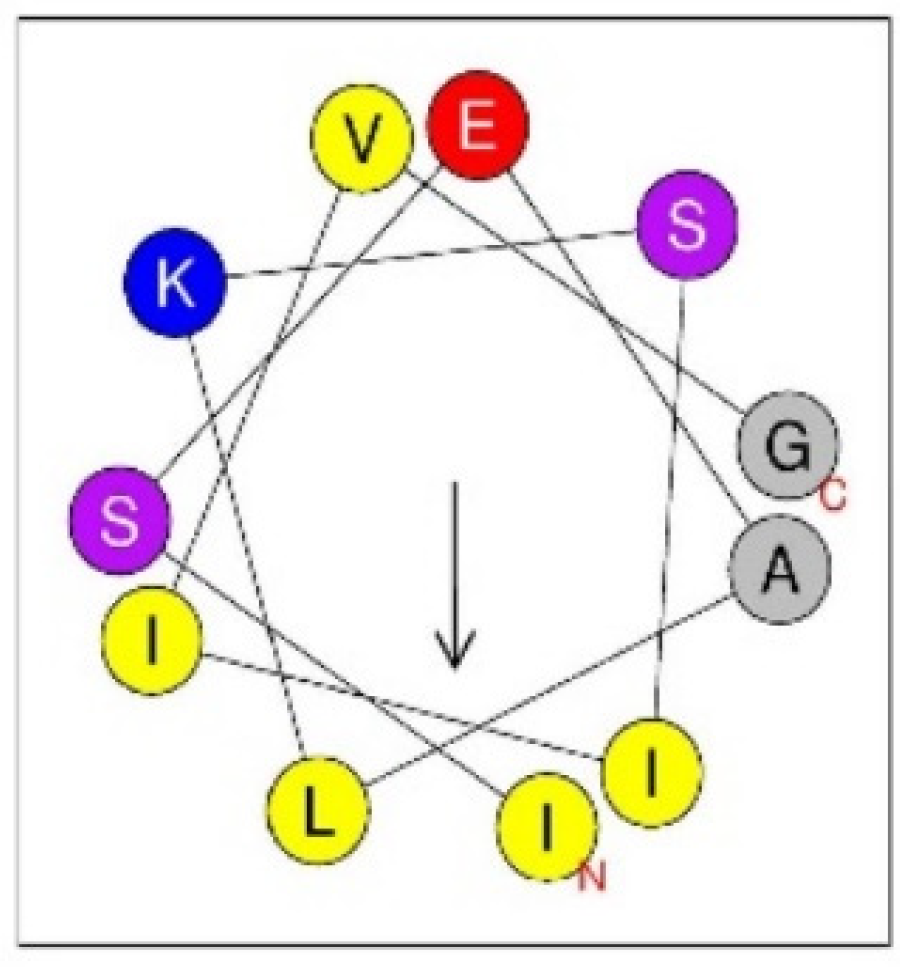
| Annulatin | ISEALKSIIVG-NH2 | This work |
| Inhibition (%) | Cleavage (%) | |||
|---|---|---|---|---|
| Peptide | NEP | ACE | NEP | ACE |
| α-campsomerin | <0.01 | <0.01 | <0.01 | <0.01 |
| β-campsomerin | <0.01 | <0.01 | <0.01 | <0.01 |
| MIC a (μmol·L−1) | Hemolytic Activity (%) | Histamine Releasing Activity at 10 μmol·L−1 (%) | ||||
|---|---|---|---|---|---|---|
| E. coli (NBRC 14237) | S. aureus (NBRC 12732) | S. cerevisiae (NBRC 10217) | ||||
| at 10 μmol·L−1 | at 12.5 μmol·L−1 | |||||
| Annulatin | Negative | Negative | Negative | - | 15.4 | 33.0 |
| Mastoparan | - | - | - | 13.5 b | - | 31.1 b |
| Melittin | - | - | - | 100.0 b | - | 64.3 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alberto-Silva, C.; Vieira Portaro, F.C.; Kodama, R.T.; Pantaleão, H.Q.; Inagaki, H.; Nihei, K.-i.; Konno, K. Comprehensive Analysis and Biological Characterization of Venom Components from Solitary Scoliid Wasp Campsomeriella annulata annulata. Toxins 2021, 13, 885. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120885
Alberto-Silva C, Vieira Portaro FC, Kodama RT, Pantaleão HQ, Inagaki H, Nihei K-i, Konno K. Comprehensive Analysis and Biological Characterization of Venom Components from Solitary Scoliid Wasp Campsomeriella annulata annulata. Toxins. 2021; 13(12):885. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120885
Chicago/Turabian StyleAlberto-Silva, Carlos, Fernanda Calheta Vieira Portaro, Roberto Tadashi Kodama, Halyne Queiroz Pantaleão, Hidetoshi Inagaki, Ken-ichi Nihei, and Katsuhiro Konno. 2021. "Comprehensive Analysis and Biological Characterization of Venom Components from Solitary Scoliid Wasp Campsomeriella annulata annulata" Toxins 13, no. 12: 885. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120885