Ecophysiology of Fusarium chaquense a Novel Type A Trichothecene Producer Species Isolated from Natural Grasses

 ,
,  and
and
Abstract
:1. Introduction
2. Results
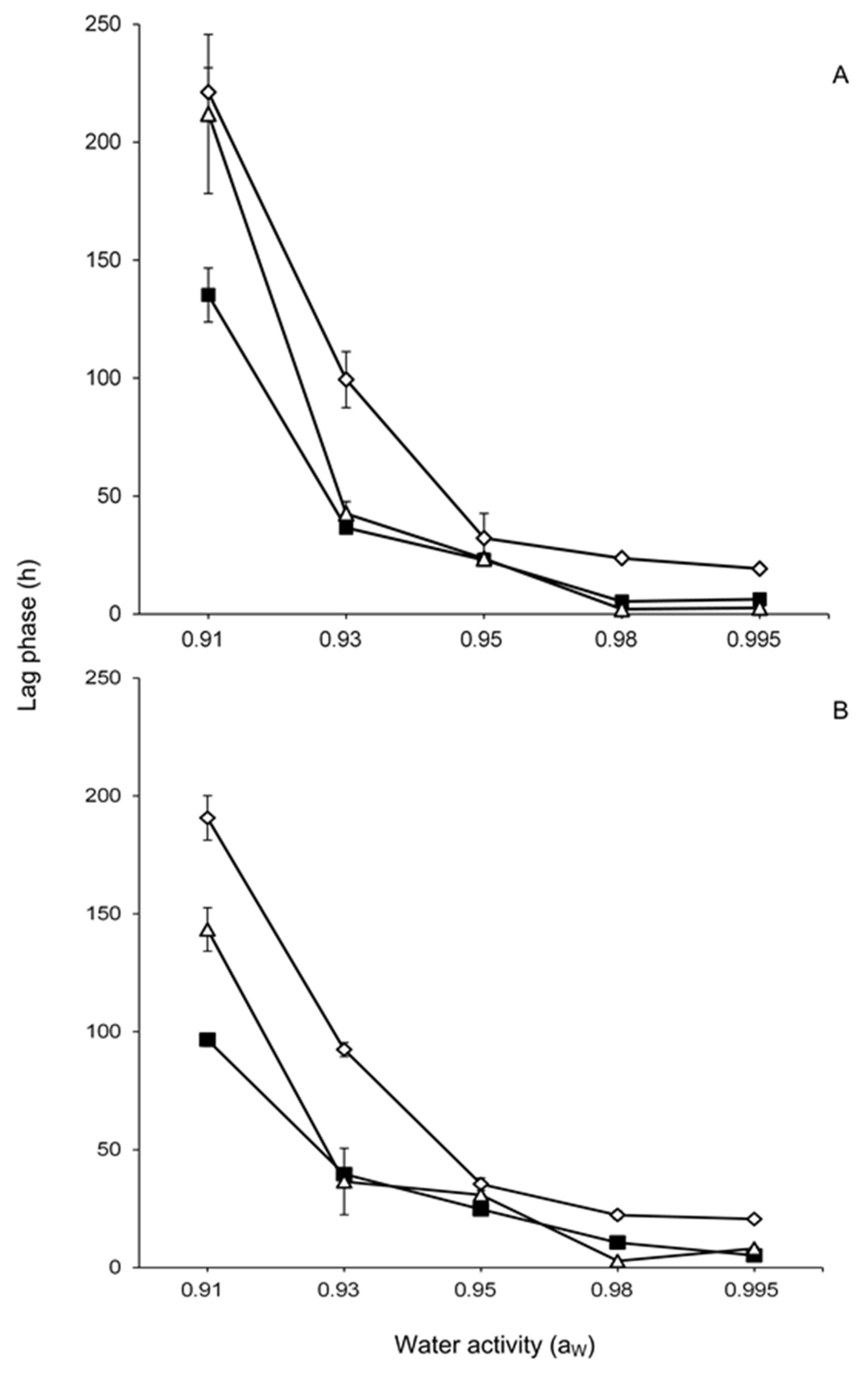
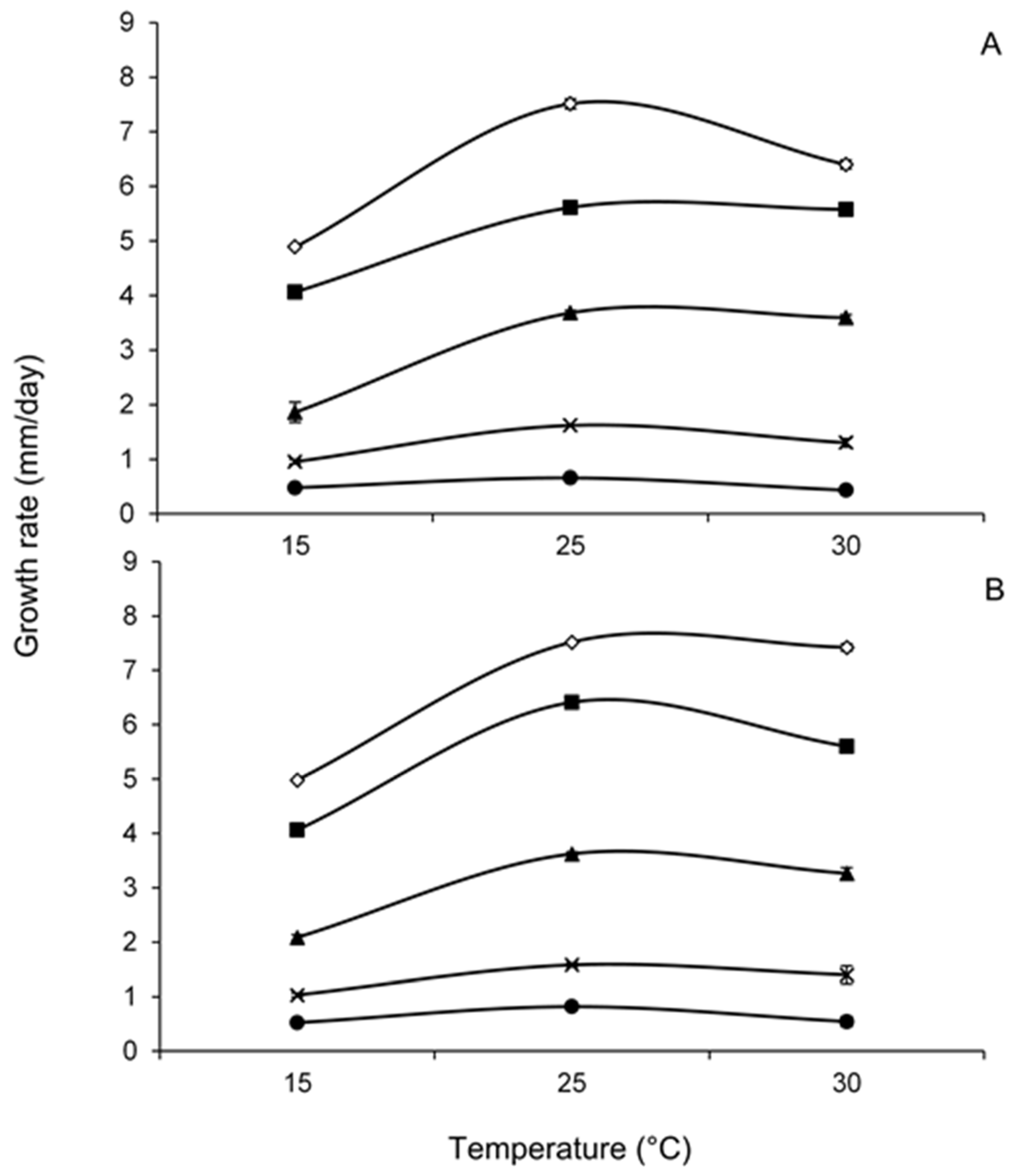
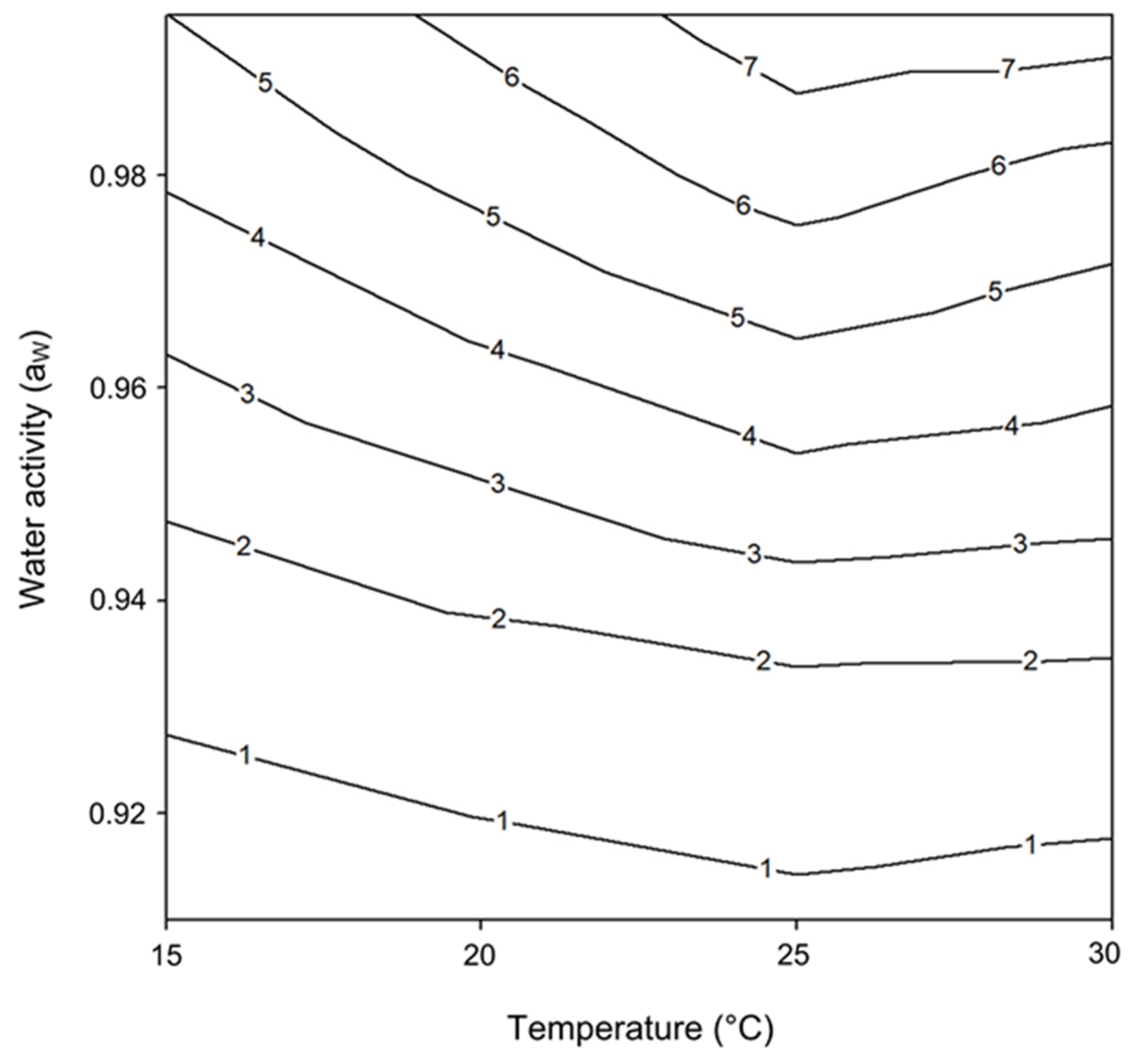
2.1. Effect of Water Activity and Temperature on Lag Phase and Growth
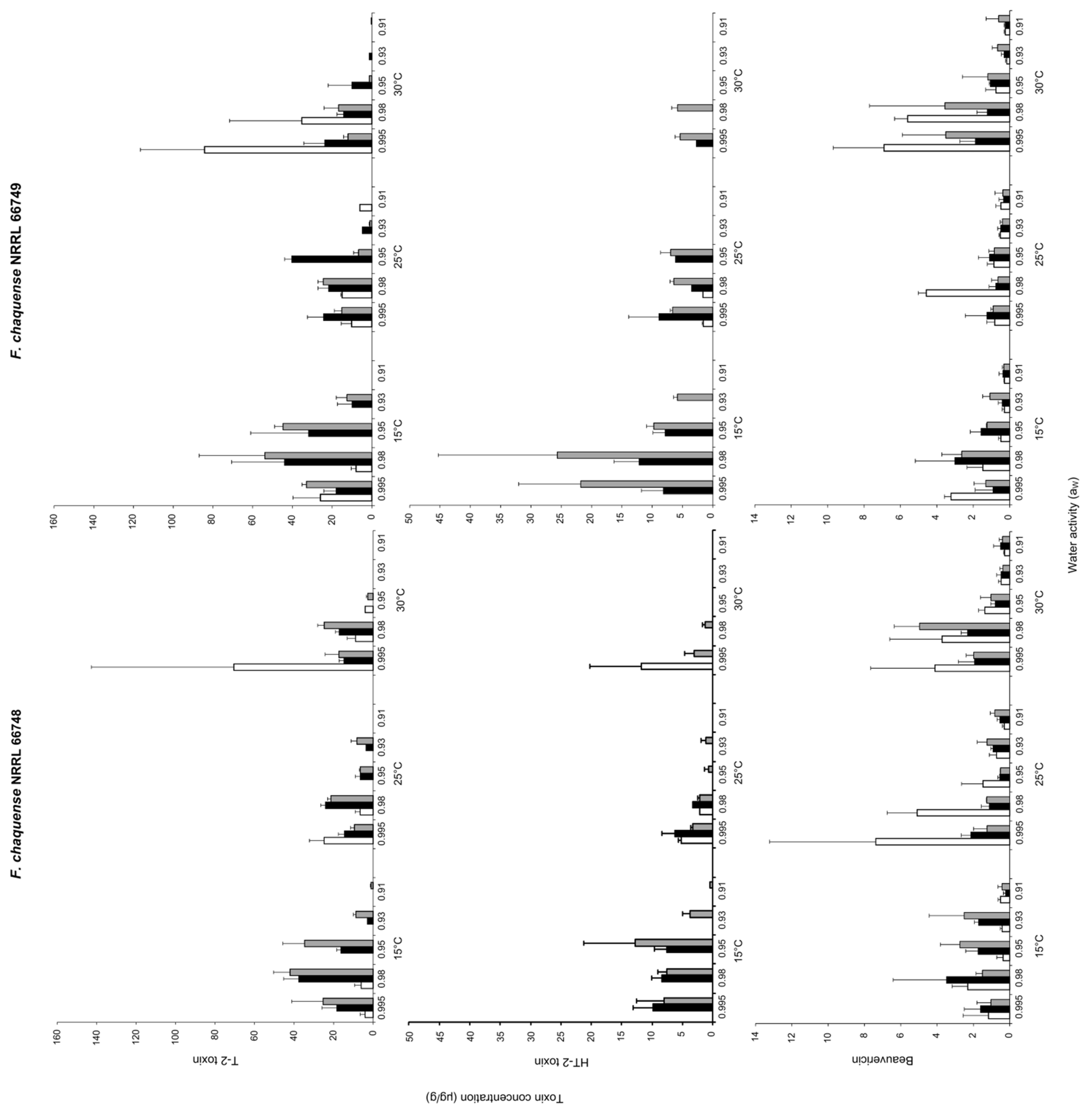
2.2. Effect of aW, Temperature and Incubation Time on Mycotoxin Production
2.3. Effect of aW and Temperature on Mycotoxin Profiles
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Medium Preparation
4.3. Inoculation, Incubation and Growth Assessment
4.4. Mycotoxin Extraction
4.5. Mycotoxin Analysis
4.5.1. Chemicals and Preparation of Standards
4.5.2. Determination of Type A Trichothecenes (T-2 and HT-2)
4.5.3. Determination of Beauvericin
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nichea, M.; Proctor, R.; Probyn, C.; Palacios, S.; Cendoya, E.; Sulyok, M.; Chulze, S.; Torres, A.; Ramirez, M. Fusarium chaquense sp. nov, a novel type A trichothecene-producing species from native grasses in a wetland ecosystem in Argentina. Mycologia 2021, in press. [Google Scholar] [CrossRef]
- Yli-mattila, T.; Ward, T.J.; Donnell, K.O.; Proctor, R.H.; Burkin, A.A.; Kononenko, G.P.; Gavrilova, O.P.; Aoki, T.; McCormick, S.P.; Yu, T. Fusarium sibiricum sp. nov., a novel type A trichothecene-producing Fusarium from northern Asia closely related to F. sporotrichioides and F. langsethiae. Int. J. Food Microbiol. 2011, 147, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.O.; Laurence, M.H.; Proctor, R.H.; Mccormick, S.P.; Summerell, B.A.; Liew, E.C.Y. Variation in type A trichothecene production and trichothecene biosynthetic genes in Fusarium goolgardi from natural ecosystems of Australia. Toxins 2015, 7, 4577–4594. [Google Scholar] [CrossRef] [PubMed]
- Nichea, M.; Palacios, S.; Chiacchiera, S.; Sulyok, M.; Krska, R.; Chulze, S.; Torres, A.; Ramirez, M. Presence of multiple mycotoxins and other fungal metabolites in native grasses from a wetland ecosystem in Argentina intended for grazing cattle. Toxins 2015, 7, 3309–3329. [Google Scholar] [CrossRef] [Green Version]
- Laraba, I.; McCormick, S.P.; Vaughan, M.M.; Geiser, D.M.; O’Donnell, K. Phylogenetic diversity, trichothecene potential, and pathogenicity within Fusarium sambucinum species complex. PLoS ONE 2021, 16, e0245037. [Google Scholar] [CrossRef] [PubMed]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Munkvold, G.P. Fusarium species and their associated mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols; Moretti, A., Susca, A., Eds.; Humana Press: New York, NY, USA, 2017; pp. 51–106. [Google Scholar]
- Proctor, R.H.; McCormick, S.P.; Gutiérrez, S. Genetic bases for variation in structure and biological activity of trichothecene toxins produced by diverse fungi. Appl. Microbiol. Biotechnol. 2020, 104, 5185–5199. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Charmley, L.L.; Rosenberg, A.; Trenholm, H.L. Factors responsible for economic losses due to Fusarium mycotoxin contamination of grains, foods and feedstuffs. In Mycotoxins in Grains: Compounds Other than Aflatoxin; Miller, J.D., Trenholm, H.L., Eds.; Eagan Press: St. Paul, MI, USA, 1994; pp. 471–486. [Google Scholar]
- Marin, S.; Magan, N.; Ramos, A.J.; Sanchis, V. Fumonisin-producing strains of Fusarium: A review of their ecophysiology. J. Food Prot. 2004, 67, 1792–1805. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Zeller, K.A.; Logrieco, A.; Mule, G.; Moretti, A.; Ritieni, A. Species diversity of and toxin production by Gibberella fujikuroi species complex strains isolated from native prairie grasses in Kansas. Appl. Environ. Microbiol. 2004, 70, 2254–2262. [Google Scholar] [CrossRef] [Green Version]
- Phan, H.T.; Burgess, L.W.; Summerell, B.A.; Bullock, S.; Liew, E.C.Y.; Clarkson, J.R. Gibberella gaditjirrii (Fusarium gaditjirrii) sp. nov., a new species from tropical grasses in Australia. Stud. Mycol. 2004, 50, 261–272. [Google Scholar]
- Sanchez Marquez, S.; Bills, G.F.; García Criado, B.; Zabalgogeazcoa, I. Diversity and structure of the fungal endophytic assemblages from two sympatric coastal grasses. Fungal. Divers. 2008, 33, 87–100. [Google Scholar]
- Medina, A.; Magan, N. Comparisons of water activity and temperature impacts on growth of Fusarium langsethiae strains from northern Europe on oat-based media. Int. J. Food Microbiol. 2010, 142, 365–369. [Google Scholar] [CrossRef] [Green Version]
- Mateo, J.J.; Mateo, R.; Jime, M. Accumulation of type A trichothecenes in maize, wheat and rice by Fusarium sporotrichioides isolates under diverse culture conditions. Int. J. Food Microbiol. 2002, 72, 115–123. [Google Scholar] [CrossRef]
- Kokkonen, M.; Ojala, L.; Parikka, P.; Jestoi, M. Mycotoxin production of selected Fusarium species at different culture conditions. Int. J. Food Microbiol. 2010, 143, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Kokkonen, M.; Jestoi, M.; Laitila, A. Mycotoxin production of Fusarium langsethiae and Fusarium sporotrichioides on cereal-based substrates. Micotoxin Res. 2012, 28, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Magan, N. Temperature and water activity effects on production of T-2 and HT-2 by Fusarium langsethiae strains from north European countries. Food Microbiol. 2011, 28, 392–398. [Google Scholar] [CrossRef] [Green Version]
- Alberto, J.A. El Chaco oriental y sus fisonomías vegetales. Geográfica Digit. 2006, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Langseth, W. Mycotoxin production and cytotoxicity of Fusarium strains isolated from Norwegian cereals. Mycopathologia 1998, 144, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Thrane, U.; Adler, A.; Clasen, P.E.; Galvano, F.; Langseth, W.; Lew, H.; Logrieco, A.; Nielsen, K.F.; Ritieni, A. Diversity in metabolite production by Fusarium langsethiae, Fusarium poae, and Fusarium sporotrichioides. Int. J. Food Microbiol. 2004, 95, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Jestoi, M.N.; Paavanen-Huhtala, S.; Parikka, P.; Yli-Mattila, T. In vitro and in vivo mycotoxin production of Fusarium species isolated from Finnish grains. Arch Phytopathol. Pflanzenschutz 2008, 41, 545–558. [Google Scholar] [CrossRef]
- Jestoi, M. Emerging Fusarium-mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin-A review. Crit. Rev. Food Sci. Nut. 2008, 48, 21–49. [Google Scholar] [CrossRef] [PubMed]
- Dallyn, H.; Fox, A. Spoilage of materials of reduced water activity by xerophilic fungi. In Microbial Growth and Survival in Extremes of Environment; Gould, G.H., Corry, J.E.L., Eds.; The Society for Applied Bacteriology, Technical Series Number 15; Academic Press: London, UK, 1980; Volume 3, pp. 129–139. [Google Scholar]
- Gerlach, W.; Nirenberg, H. The Genus Fusarium—A Pictorial Atlas; Mitteilungen aus der Biologischen Bundesanstalt fur Land- und Forstwirtschaft Berlin-Dahlem; Kommissionsverlag P. Parey: Berlin, Germany, 1982; Volume 209, pp. 1–406. [Google Scholar]
- Pascale, M.; Panzarini, G.; Visconti, A. Determination of HT-2 and T-2 toxins in oats and wheat by ultra-performance liquid chromatography with photodiode array detection. Talanta 2012, 89, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Meca, G.; Font, G.; Ruiz, M.J. Study of the cytotoxic activity of beauvericin and fusaproliferin and bioavailability in vitro on Caco-2 cells. Food Chem. Toxicol. 2012, 50, 2356–2361. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.; Gonzalez, L.; Cuadroda, M.; Robledo, C. InfoStat Versión 2016; Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2018. [Google Scholar]



 ), 15 (
), 15 (  ) and 25 (
) and 25 (  ) days.
), 15 ( ) and 25 ( ) days.
) days.
), 15 ( ) and 25 ( ) days.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Df a | Lag Phase (h) | Growth Rate (mm/day) | ||
|---|---|---|---|---|---|
| MS b | F c | MS | F | ||
| S | 1 | 0.01 | 31.85 * | 30.66 | 831.09 * |
| T | 2 | 0.11 | 241.97 * | 0.09 | 23.63 * |
| aW | 4 | 0.14 | 307.61 * | 0.63 | 168.57 * |
| S × T | 2 | 0.01 | 19.79 * | 1.08 | 288.61 * |
| S × aW | 4 | 0.01 | 15.19 * | 13.41 | 3585.19 * |
| T × aW | 8 | 0.04 | 98.41 * | 0.03 | 7.42 * |
| S × T × aW | 8 | 0.01 | 17.38 * | 0.10 | 25.91 * |
| NRRL 66748 | |||||||
| Source of variation | Df a | T-2 | HT-2 | BEA | |||
| MS b | F c | MS | F | MS | F | ||
| T | 2 | 188.37 | 1.32 | 59.04 | 14.62 * | 0.01 | 0.12 |
| aW | 4 | 2938.12 | 20.64 * | 72.81 | 18.03 * | 3.14 | 31.28 * |
| D | 2 | 220.05 | 1.55 | 24.29 | 6.01 * | 0.12 | 1.22 |
| T × aW | 8 | 363.2 | 2.55 | 10.57 | 2.62 * | 0.44 | 4.36 * |
| T × D | 4 | 722.38 | 5.07 * | 41.1 | 10.18 * | 0.66 | 6.52 * |
| aW × D | 8 | 558.43 | 3.92 * | 6.38 | 1.58 | 0.33 | 3.24 * |
| T × aW × D | 16 | 228.5 | 1.61 | 11.12 | 2.75 * | 0.24 | 2.41 * |
| NRRL 66749 | |||||||
| Source of variation | Df a | T-2 | HT-2 | BEA | |||
| MS b | F c | MS | F | MS | F | ||
| T | 2 | 1.02 | 7.26 | 9.13 | 30.46 * | 0.3 | 2.73 |
| aW | 4 | 7.03 | 49.84 * | 7.6 | 25.37 | 3.48 | 31.46 * |
| D | 2 | 1.21 | 8.54 * | 14.96 | 49.91 * | 0.2 | 1.85 |
| T × aW | 8 | 0.3 | 2.11 | 0.86 | 2.87 | 0.4 | 3.60 * |
| T × D | 4 | 0.4 | 2.82 | 2.56 | 8.55 | 0.2 | 1.85 |
| aW × D | 8 | 0.71 | 5.05 * | 1.97 | 6.59 | 0.3 | 2.75 * |
| T × aW × D | 16 | 0.12 | 0.88 | 0.57 | 1.9 | 0.25 | 2.25 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nichea, M.J.; Cendoya, E.; Haidukowski, M.; Torres, A.M.; Ramirez, M.L. Ecophysiology of Fusarium chaquense a Novel Type A Trichothecene Producer Species Isolated from Natural Grasses. Toxins 2021, 13, 895. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120895
Nichea MJ, Cendoya E, Haidukowski M, Torres AM, Ramirez ML. Ecophysiology of Fusarium chaquense a Novel Type A Trichothecene Producer Species Isolated from Natural Grasses. Toxins. 2021; 13(12):895. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120895
Chicago/Turabian StyleNichea, Maria J., Eugenia Cendoya, Miriam Haidukowski, Adriana M. Torres, and María L. Ramirez. 2021. "Ecophysiology of Fusarium chaquense a Novel Type A Trichothecene Producer Species Isolated from Natural Grasses" Toxins 13, no. 12: 895. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120895