rDromaserpin: A Novel Anti-Hemostatic Serpin, from the Salivary Glands of the Hard Tick Hyalomma dromedarii

 , and
, and 
Abstract
:1. Introduction
2. Results
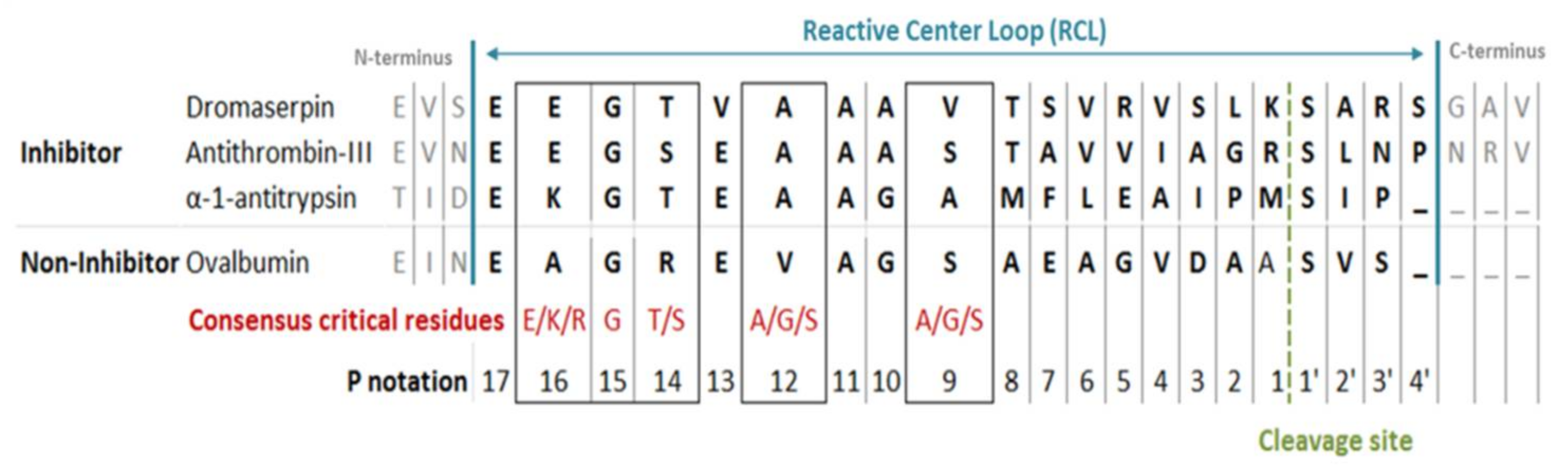
2.1. Bioinformatic and Phylogenetic Analysis on Dromaserpin Sequence
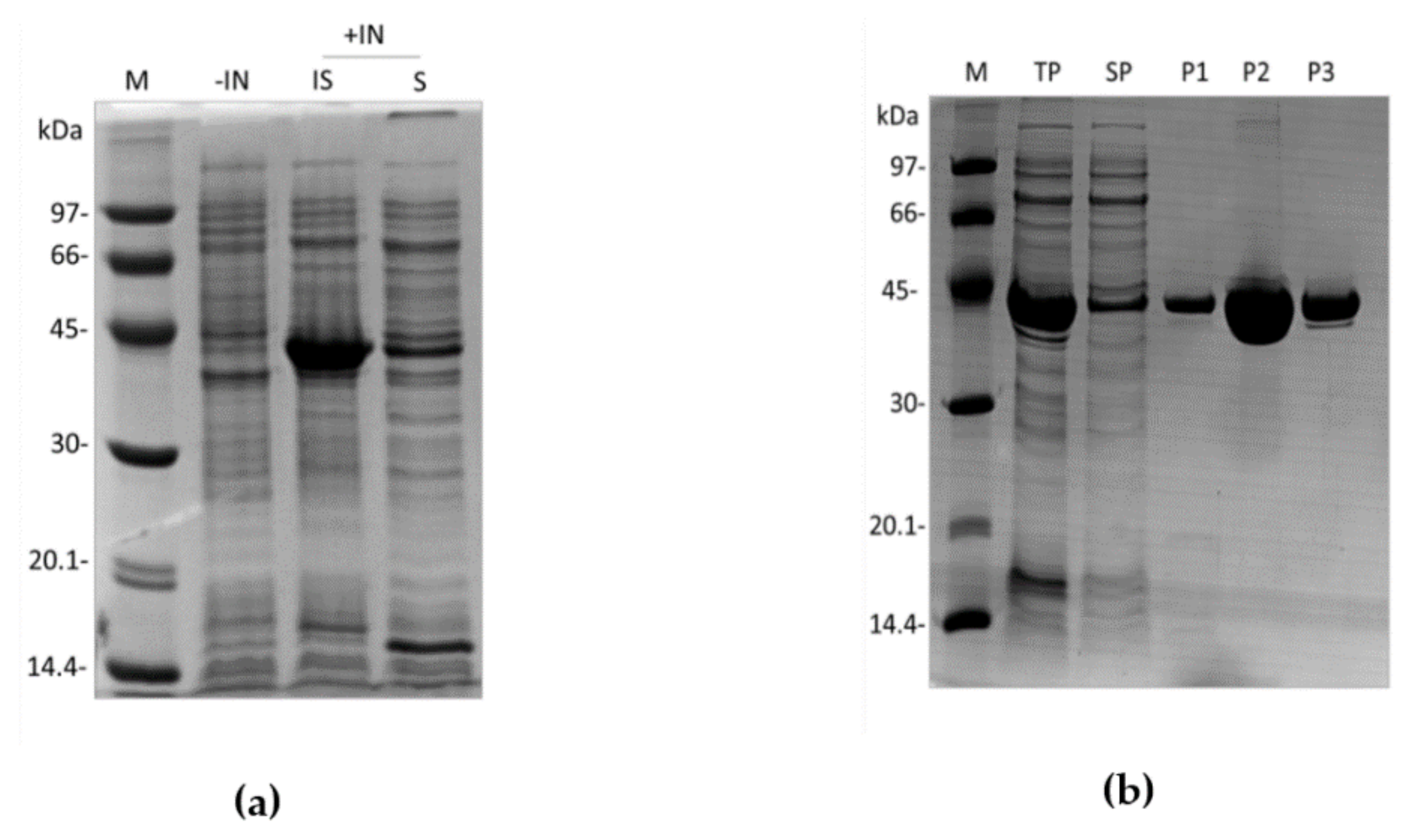
2.2. Expression and Purification of the Recombinant Dromaserpin (rDromaserpin)
2.3. Structural Characterization of the rDromaserpin
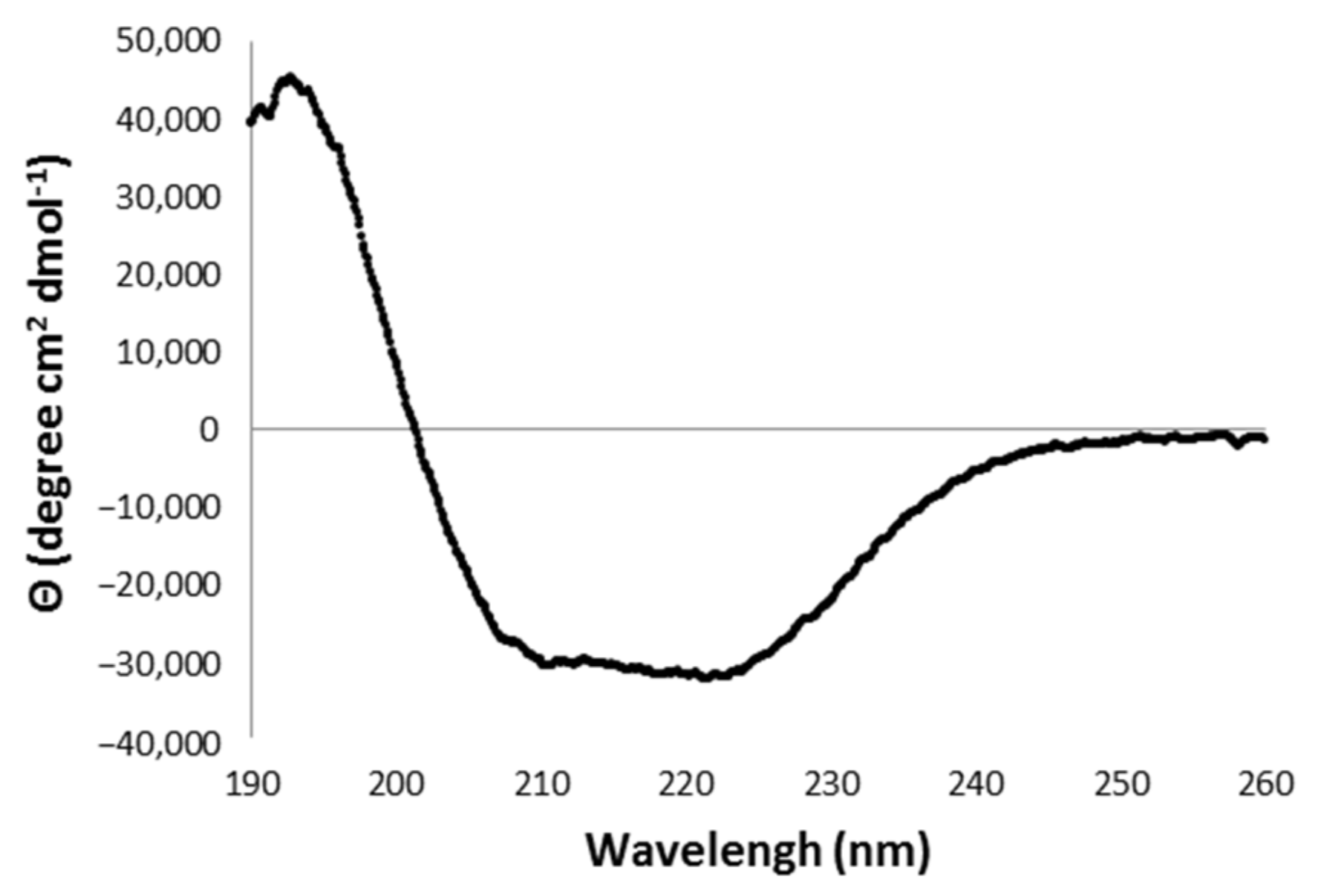
2.3.1. Analysis of the Secondary Structure of rDromaserpin
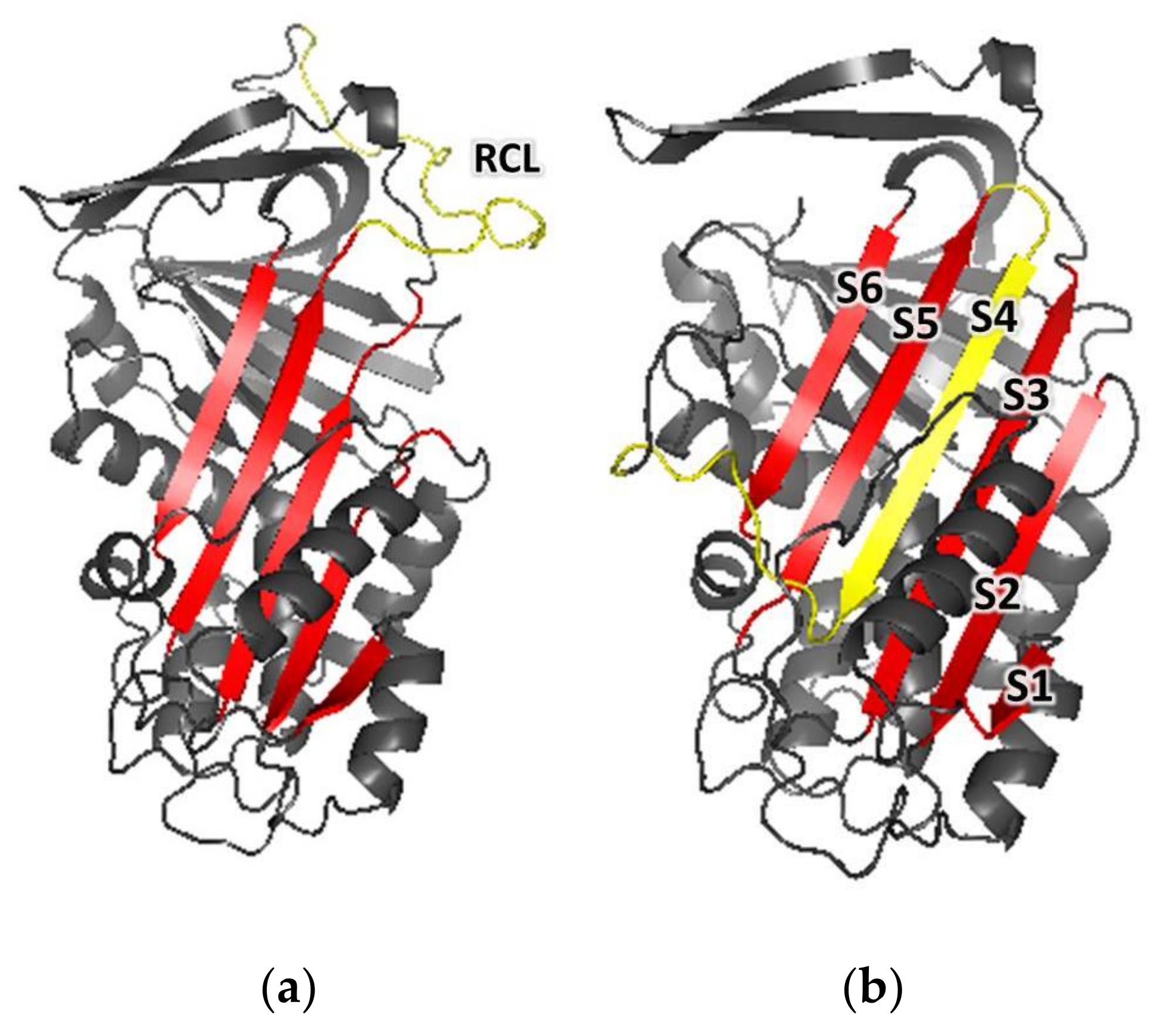
2.3.2. Comparative Modeling
2.4. Functional Characterization of the rDromaserpin
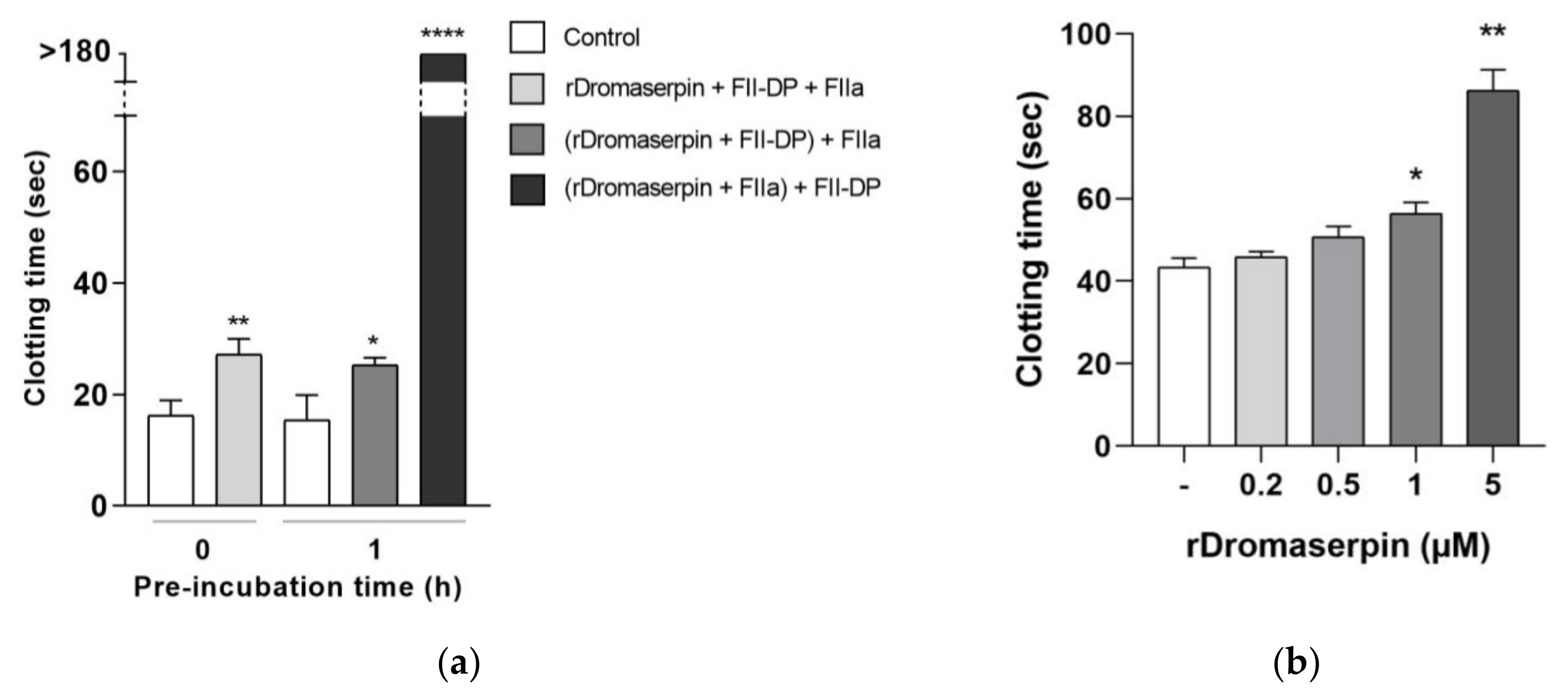
2.4.1. Global Blood Coagulation Assays
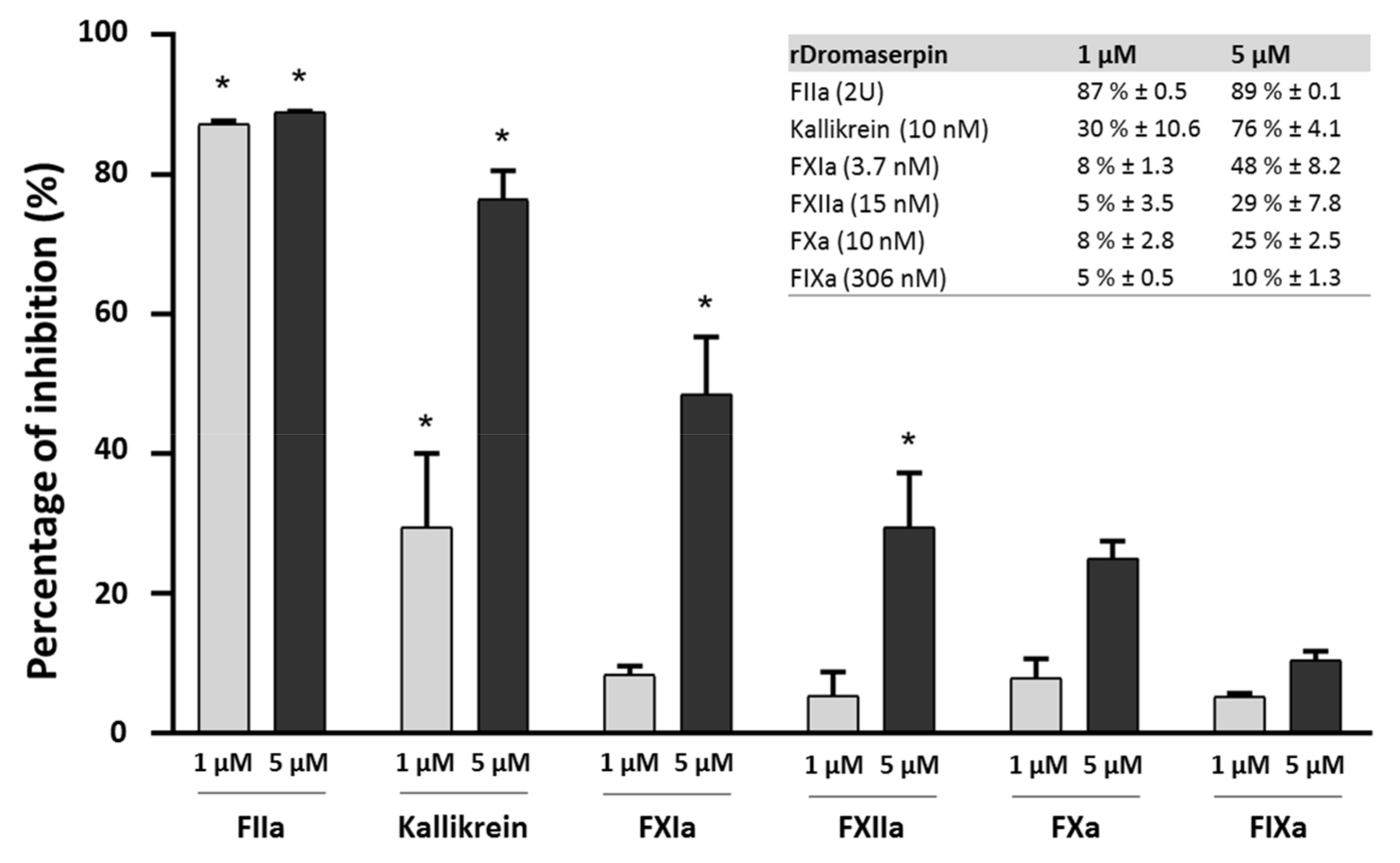
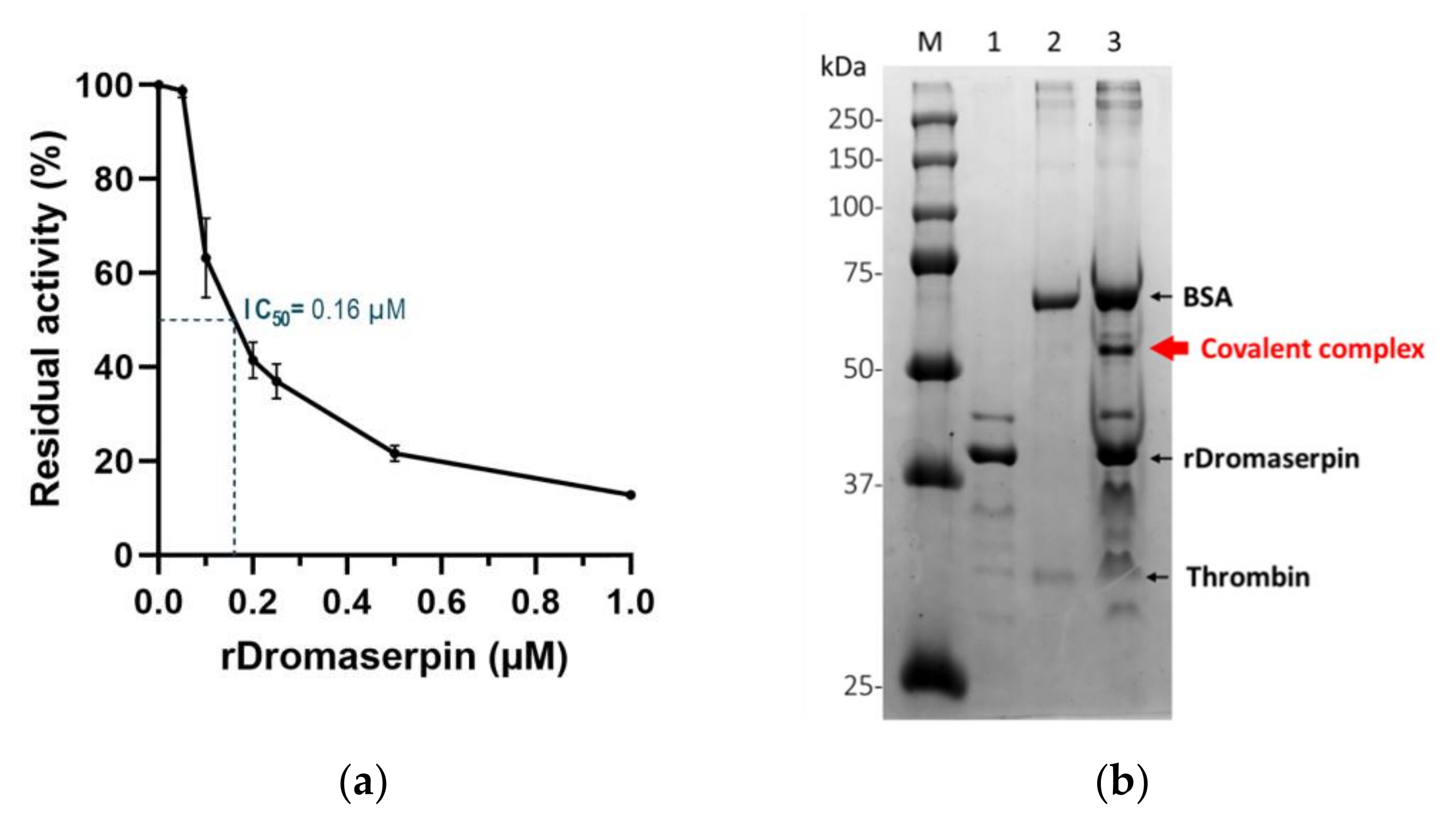
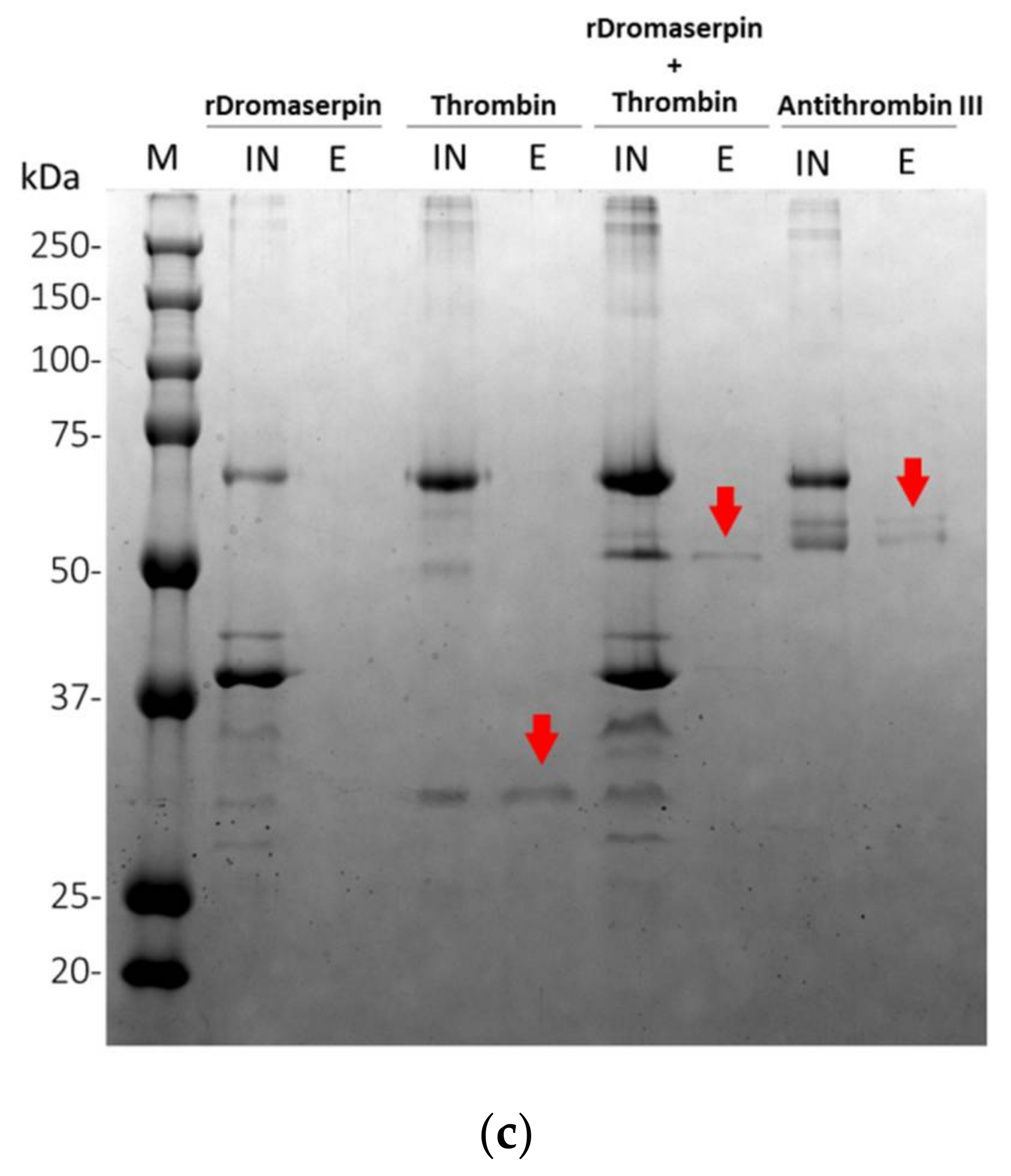
2.4.2. Protease Inhibition Profiling of rDromaserpin
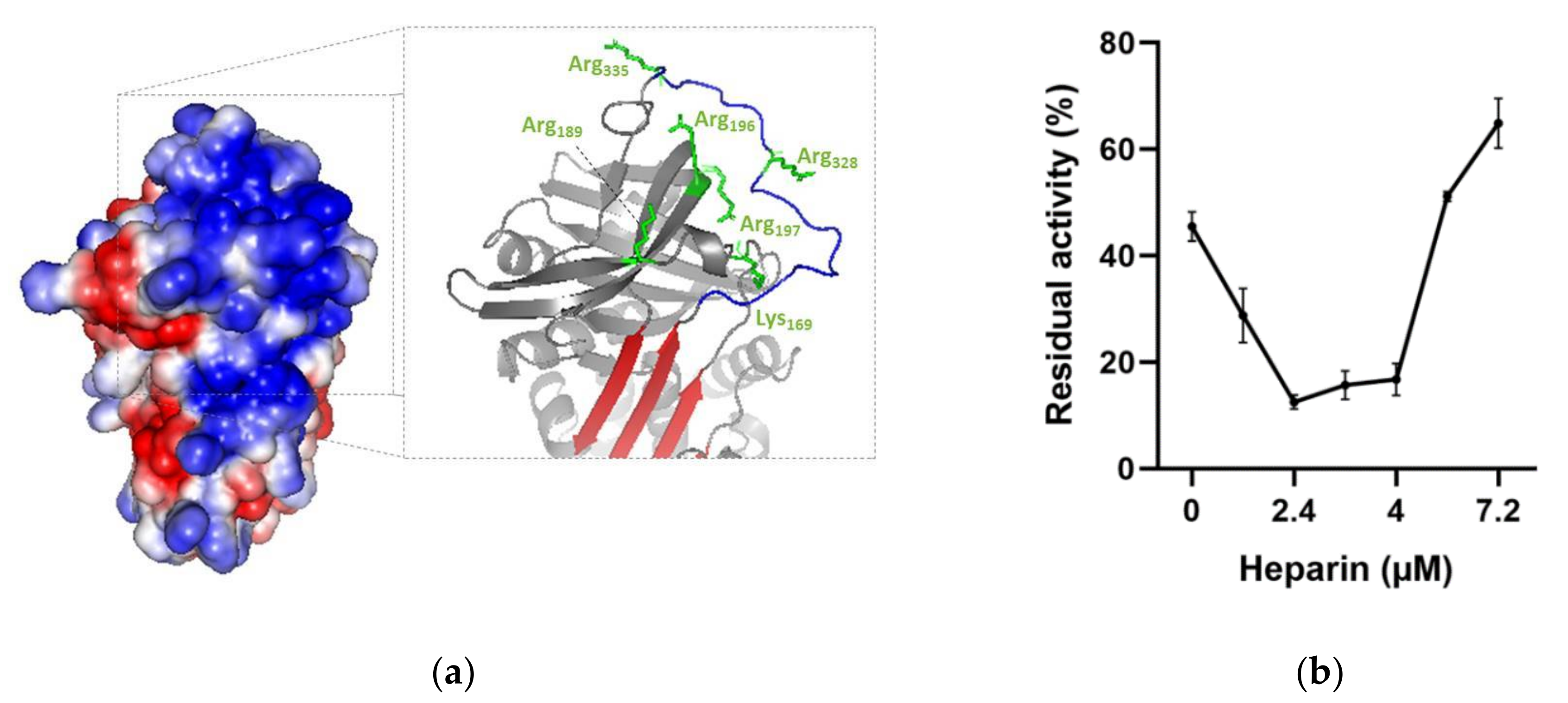
2.4.3. Effect of Heparin on the Rate of Thrombin Inhibition by rDromaserpin
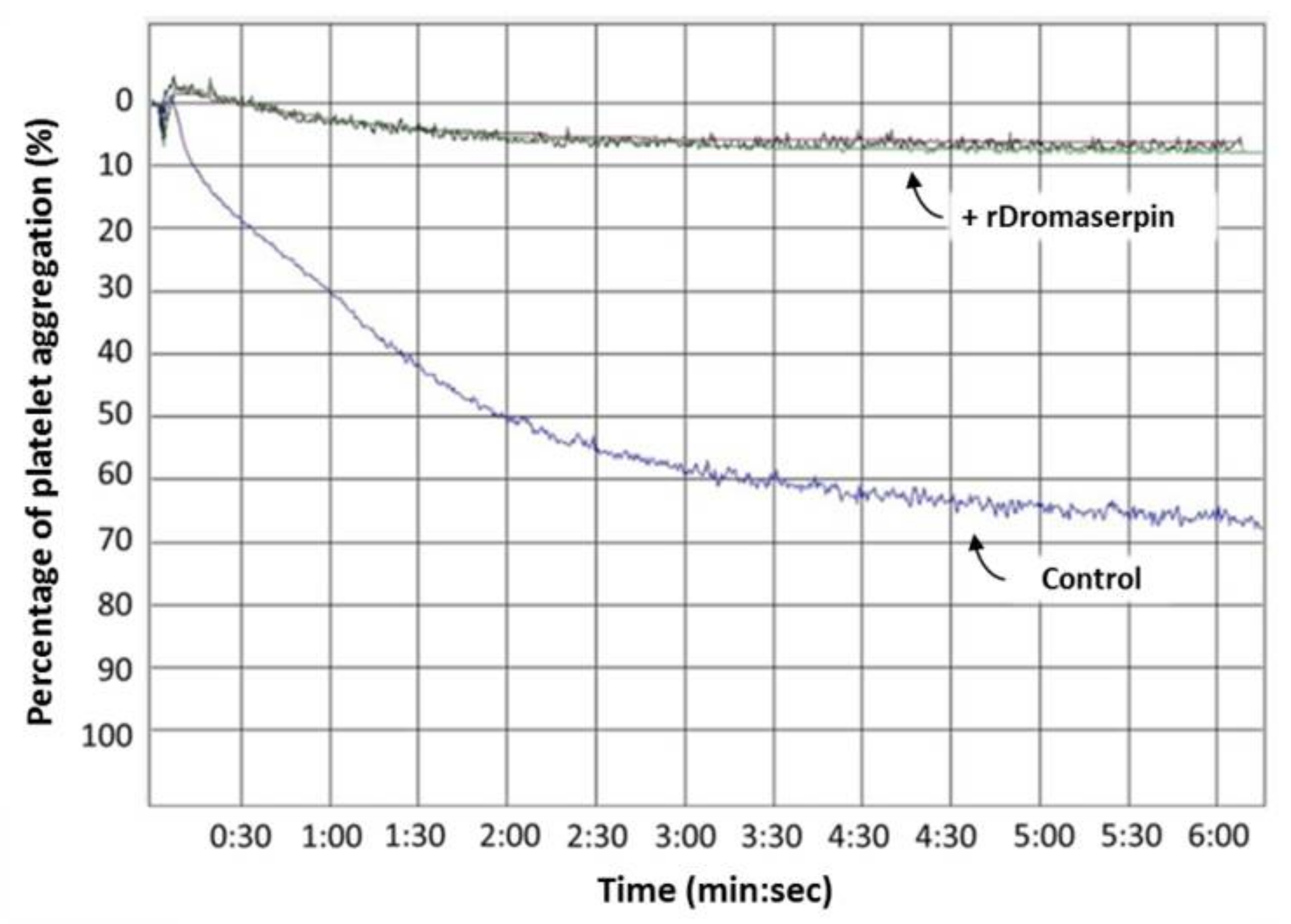
2.4.4. Dromaserpin Inhibits the Function of Thrombin in Activating Platelet Aggregation
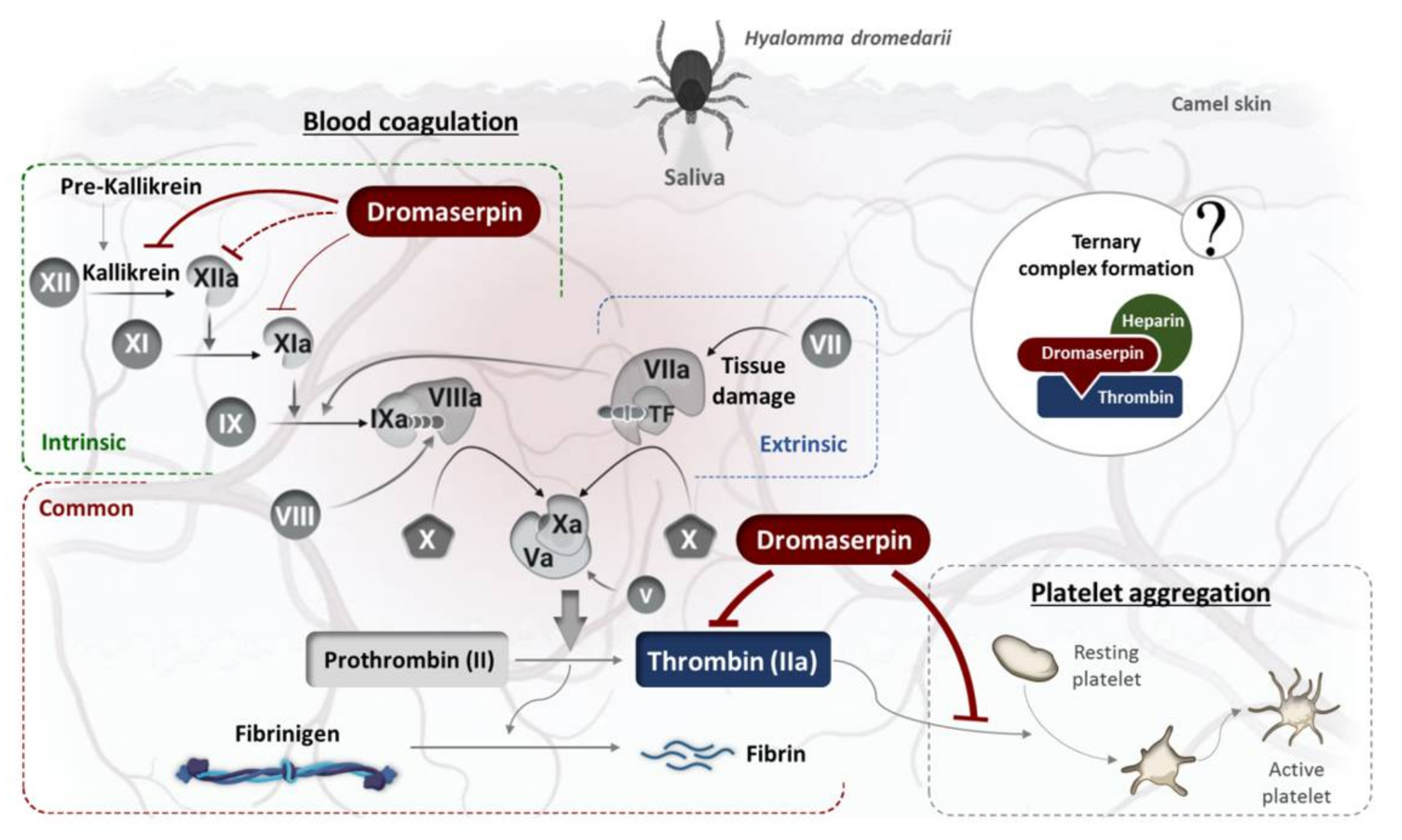
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents and Chemicals
5.2. Bioinformatics Analysis on Pedicted Dromaserpin Sequence
5.3. Plasmid Construction-Expression and Purification of the Recombinant Dromaserpin
5.4. Structural Characterization of the rDromaserpin
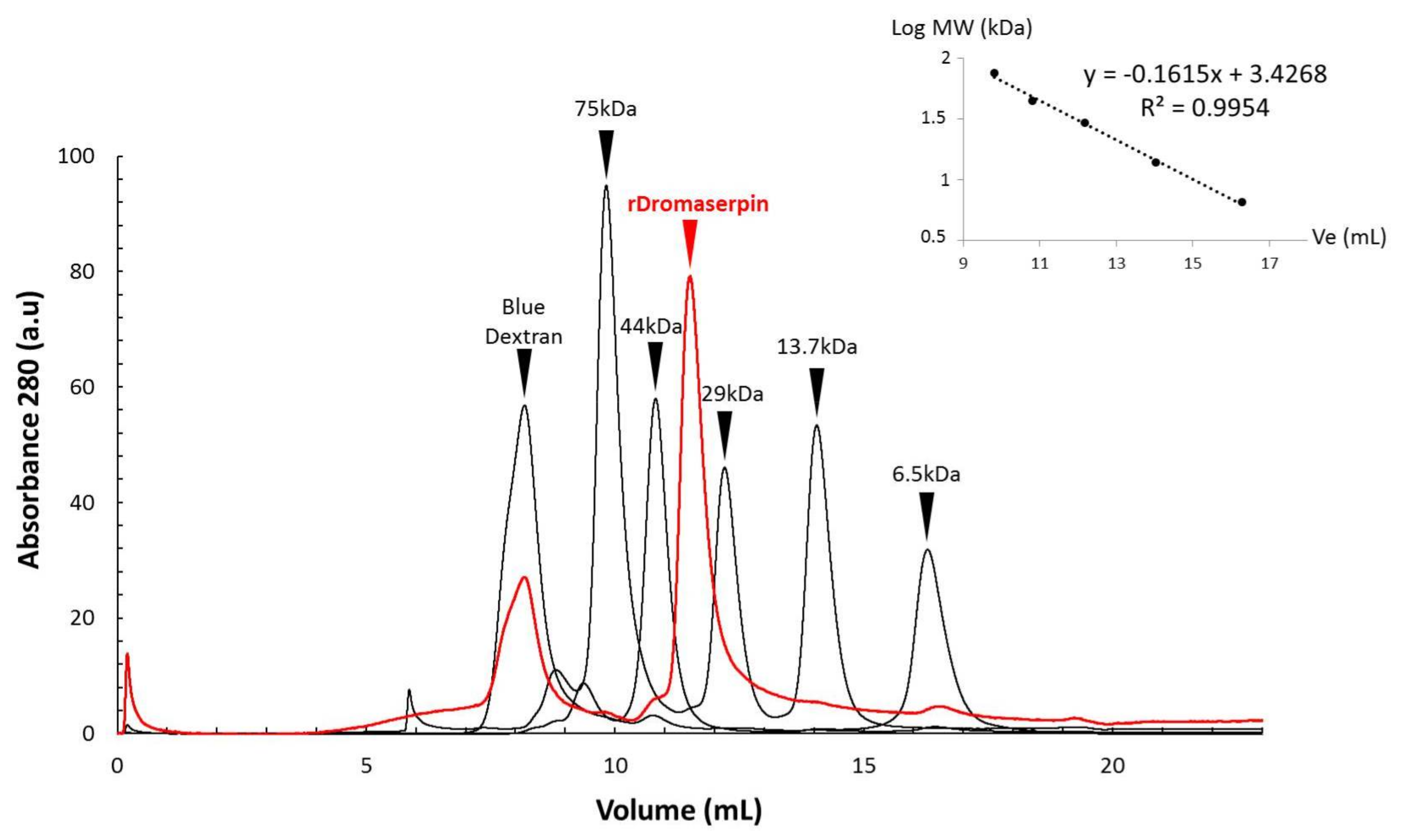
5.4.1. Determination of Molecular Size by Gel Filtration
5.4.2. Circular Dichroism (CD) Spectroscopy
5.4.3. Comparative Modeling and Structural Analysis of Dromaserpin
5.5. Functional Characterization of the rDromaserpin
5.5.1. Global blood Coagulation Assays
5.5.2. Protease Inhibition Assays
5.5.3. Thrombin Inhibition by rDromaserpin in the Presence of Heparin
5.5.4. Evaluation of Binding to Heparin-Sepharose Resin
5.5.5. Effect of rDromaserpin on Platelet Aggregation
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- The World Health Organization Web Site. Available online: https://www.who.int/health-topics/cardiovascular-diseases/#tab=tab_1 (accessed on 17 September 2020).
- Stoney, C.M.; Kaufmann, P.G.; Czajkowski, S.M. Cardiovascular disease: Psychological, social, and behavioral influences: Introduction to the special issue. Am. Psychol. 2018, 73, 949–954. [Google Scholar] [CrossRef]
- Chang, J.C. Hemostasis based on a bovel “two-path unifying theory” and classification of hemostatic disorders. Blood Coagul. Fibrinolysis 2018, 29, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Ogedegbe, H.O. An overview of hemostasis. Lab. Med. 2002, 33, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Versteeg, H.H.; Heemskerk, J.W.M.; Levi, M.; Reitsma, P.H. New fundamentals in hemostasis. Physiol. Rev. 2013, 93, 327–358. [Google Scholar] [CrossRef] [Green Version]
- Mussbacher, M.; Kral-Pointner, J.B.; Salzmann, M.; Schrottmaier, W.C.; Assinger, A. Mechanisms of hemostasis: Contributions of platelets, coagulation factors, and the vessel wall. In Fundamentals of Vascular Biology; Geiger, M., Ed.; Learning Materials in Biosciences; Springer International Publishing: Cham, Switzerland, 2019; pp. 145–169. ISBN 978-3-030-12270-6. [Google Scholar]
- Page, M.J.; Macgillivray, R.T.A.; Di Cera, E. Determinants of specificity in coagulation proteases. J. Thromb. Haemost. 2005, 3, 2401–2408. [Google Scholar] [CrossRef]
- Gettins, P.G.W. Serpin structure, mechanism, and function. Chem. Rev. 2002, 102, 4751–4804. [Google Scholar] [CrossRef]
- Crawley, J.T.B.; Zanardelli, S.; Chion, C.K.N.K.; Lane, D.A. The central role of thrombin in hemostasis. J. Thromb. Haemost. 2007, 5 (Suppl. S1), 95–101. [Google Scholar] [CrossRef] [PubMed]
- Siller-Matula, J.M.; Schwameis, M.; Blann, A.; Mannhalter, C.; Jilma, B. Thrombin as a multi-functional enzyme. Focus on in vitro and in vivo effects. Thromb. Haemost. 2011, 106, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- Mega, J.L.; Simon, T. Pharmacology of antithrombotic drugs: An assessment of oral antiplatelet and anticoagulant treatments. Lancet 2015, 386, 281–291. [Google Scholar] [CrossRef]
- Tang, N.; Bai, H.; Chen, X.; Gong, J.; Li, D.; Sun, Z. Anticoagulant treatment is associated with decreased mortality in severe coronavirus disease 2019 patients with coagulopathy. J. Thromb. Haemost. 2020, 18, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Carvalhal, F.; Cristelo, R.R.; Resende, D.I.S.P.; Pinto, M.M.M.; Sousa, E.; Correia-da-Silva, M. Antithrombotics from the sea: Polysaccharides and beyond. Mar. Drugs 2019, 17, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, A.; Yaron, J.R.; Zhang, L.; Ambadapadi, S. Overview of serpins and their roles in biological systems. Methods Mol. Biol. 2018, 1826, 1–7. [Google Scholar] [CrossRef]
- Lucas, A.; Yaron, J.R.; Zhang, L.; Macaulay, C.; McFadden, G. Serpins: Development for therapeutic applications. Methods Mol. Biol. 2018, 1826, 255–265. [Google Scholar] [CrossRef]
- Van Gent, D.; Sharp, P.; Morgan, K.; Kalsheker, N. Serpins: Structure, Function and Molecular Evolution. Int. J. Biochem. Cell Biol. 2003, 35, 1536–1547. [Google Scholar] [CrossRef]
- Stein, P.E.; Leslie, A.G.; Finch, J.T.; Carrell, R.W. Crystal structure of uncleaved ovalbumin at 1.95 A Resolution. J. Mol. Biol. 1991, 221, 941–959. [Google Scholar] [CrossRef]
- Engh, R.; Löbermann, H.; Schneider, M.; Wiegand, G.; Huber, R.; Laurell, C.B. The S variant of human alpha 1-antitrypsin, structure and implications for function and metabolism. Protein Eng. 1989, 2, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Skinner, R.; Abrahams, J.P.; Whisstock, J.C.; Lesk, A.M.; Carrell, R.W.; Wardell, M.R. The 2.6 A structure of antithrombin indicates a conformational change at the heparin binding site. J. Mol. Biol. 1997, 266, 601–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, H.M.; Mottonen, J.; Goldsmith, E.J.; Gerard, R.D. Engineering of plasminogen activator inhibitor-1 to reduce the rate of latency tansition. Nat. Struct. Biol. 1995, 2, 442–445. [Google Scholar] [CrossRef]
- Chen, H.; Davids, J.A.; Zheng, D.; Bryant, M.; Bot, I.; Van Berckel, T.J.C.; Biessen, E.; Pepine, C.; Ryman, K.; Progulski-Fox, A.; et al. The serpin solution; targeting thrombotic and thrombolytic serine proteases in inflammation. Cardiovasc. Hematol. Disord. Drug Targets 2013, 13, 99–110. [Google Scholar] [CrossRef]
- Irving, J.A.; Ekeowa, U.I.; Belorgey, D.; Haq, I.; Gooptu, B.; Miranda, E.; Pérez, J.; Roussel, B.D.; Ordóñez, A.; Dalton, L.E.; et al. The serpinopathies studying serpin polymerization in vivo. Meth. Enzymol. 2011, 501, 421–466. [Google Scholar] [CrossRef]
- Rau, J.C.; Beaulieu, L.M.; Huntington, J.A.; Church, F.C. Serpins in thrombosis, hemostasis and fibrinolysis. J. Thromb. Haemost. 2007, 5 (Suppl. S1), 102–115. [Google Scholar] [CrossRef] [PubMed]
- Bang, N.U. Leeches, snakes, ticks, and vampire bats in today’s cardiovascular drug development. Circulation 1991, 84, 436–438. [Google Scholar] [CrossRef] [Green Version]
- Jmel, M.A.; Aounallah, H.; Bensaoud, C.; Mekki, I.; Chmelař, J.; Faria, F.; M’ghirbi, Y.; Kotsyfakis, M. Insights into the role of tick salivary protease inhibitors during ectoparasite-host crosstalk. Int. J. Mol. Sci. 2021, 22, 892. [Google Scholar] [CrossRef] [PubMed]
- Chmelař, J.; Kotál, J.; Kovaříková, A.; Kotsyfakis, M. The use of tick salivary proteins as novel therapeutics. Front. Physiol. 2019, 10, 812. [Google Scholar] [CrossRef] [PubMed]
- Chmelař, J.; Kotál, J.; Langhansová, H.; Kotsyfakis, M. Protease inhibitors in tick saliva: The role of serpins and cystatins in tick-host-pathogen interaction. Front. Cell Infect. Microbiol. 2017, 7, 216. [Google Scholar] [CrossRef] [PubMed]
- Irving, J.A.; Pike, R.N.; Lesk, A.M.; Whisstock, J.C. Phylogeny of the serpin superfamily: Implications of patterns of amino acid conservation for structure and function. Genome Res. 2000, 10, 1845–1864. [Google Scholar] [CrossRef]
- Schechter, I.; Berger, A. On the size of the active site in proteases. I. Papain. 1967. Biochem. Biophys. Res. Commun. 2012, 425, 497–502. [Google Scholar] [CrossRef]
- Nguyen, K.D.; Pan, Y. A knowledge-based multiple-sequence alignment algorithm. IEEE/ACM Trans. Comput. Biol. Bioinform. 2013, 10, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Cao, J.; Zhou, Y.; Zhang, H.; Zhou, J. Isolation and characterization of two novel serpins from the tick Rhipicephalus haemaphysaloides. Ticks Tick Borne Dis. 2013, 4, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirloni, L.; Seixas, A.; Mulenga, A.; Vaz, I.d.S.; Termignoni, C. A Family of serine protease inhibitors (serpins) in the cattle tick Rhipicephalus (boophilus) microplus. Exp. Parasitol. 2014, 137, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.R.; Schilstra, M.J. Circular dichroism and its application to the study of biomolecules. Methods Cell Biol. 2008, 84, 263–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanrattana, W.; Maas, C.; De Maat, S. SERPINs—From trap to treatment. Front. Med. 2019, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, V.L. Prothrombin time and partial thromboplastin time assay considerations. Clin. Lab. Med. 2009, 29, 253–263. [Google Scholar] [CrossRef]
- Higgins, W.J.; Fox, D.M.; Kowalski, P.S.; Nielsen, J.E.; Worrall, D.M. Heparin enhances serpin inhibition of the cysteine protease cathepsin L. J. Biol. Chem. 2010, 285, 3722–3729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmelar, J.; Calvo, E.; Pedra, J.H.F.; Francischetti, I.M.B.; Kotsyfakis, M. Tick salivary secretion as a source of antihemostatics. J. Proteom. 2012, 75, 3842–3854. [Google Scholar] [CrossRef] [Green Version]
- Bensaoud, C.; Aounallah, H.; Sciani, J.M.; Faria, F.; Chudzinski-Tavassi, A.M.; Bouattour, A.; M’ghirbi, Y. Proteomic informed by transcriptomic for salivary glands components of the camel tick Hyalomma dromedarii. BMC Genom. 2019, 20, 675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensaoud, C.; Nishiyama, M.Y.; Ben Hamda, C.; Lichtenstein, F.; Castro de Oliveira, U.; Faria, F.; Loiola Meirelles Junqueira-de-Azevedo, I.; Ghedira, K.; Bouattour, A.; M’Ghirbi, Y.; et al. De novo assembly and annotation of Hyalomma dromedarii Tick (Acari: Ixodidae) sialotranscriptome with regard to gender differences in gene expression. Parasit. Vectors 2018, 11, 314. [Google Scholar] [CrossRef]
- Schussler, G.C. The thyroxine-binding proteins. Thyroid 2000, 10, 141–149. [Google Scholar] [CrossRef]
- Simard, M.; Underhill, C.; Hammond, G.L. Functional implications of corticosteroid-binding globulin N-glycosylation. J. Mol. Endocrinol. 2018, 60, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Polderdijk, S.G.I.; Huntington, J.A. Identification of serpins specific for activated protein C using a lysate-based screening assay. Sci. Rep. 2018, 8, 8793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, T.H.; Hejgaard, J.; Saunders, N.F.W.; Cavicchioli, R.; Curmi, P.M.G. Serpins in unicellular eukarya, archaea, and bacteria: Sequence analysis and evolution. J. Mol. Evol. 2004, 59, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Radulovic, Z.; Mulenga, A. Target validation of highly conserved Amblyomma americanum tick saliva serine protease inhibitor 19. Ticks Tick Borne Dis. 2016, 7, 405–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Song, Z.; Chen, J.; Wu, Q.; Zhou, X.; Ni, X.; Dai, J. The immunosuppressive functions of two novel tick serpins, HlSerpin-a and HlSerpin-b, from Haemaphysalis longicornis. Immunology 2019, 159, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Mahon, B.P.; Ambadapadi, S.; Yaron, J.R.; Lomelino, C.L.; Pinard, M.A.; Keinan, S.; Kurnikov, I.; Macaulay, C.; Zhang, L.; Reeves, W.; et al. Crystal structure of cleaved Serp-1, a Myxomavirus-derived immune modulating serpin: Structural design of serpin reactive center loop peptides with improved therapeutic function. Biochemistry 2018, 57, 1096–1107. [Google Scholar] [CrossRef]
- Pongprayoon, P.; Niramitranon, J.; Kaewhom, P.; Kaewmongkol, S.; Suwan, E.; Stich, R.W.; Jittapalapong, S. Dynamic and structural insights into tick serpin from Ixodes ricinus. J. Biomol. Struct. Dyn. 2019, 38, 2296–2303. [Google Scholar] [CrossRef]
- Chmelar, J.; Oliveira, C.J.; Rezacova, P.; Francischetti, I.M.B.; Kovarova, Z.; Pejler, G.; Kopacek, P.; Ribeiro, J.M.C.; Mares, M.; Kopecky, J.; et al. A Tick salivary protein targets cathepsin G and chymase and inhibits host inflammation and platelet aggregation. Blood 2011, 117, 736–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovářová, Z.; Chmelař, J.; Sanda, M.; Brynda, J.; Mareš, M.; Rezáčová, P. Crystallization and diffraction analysis of the serpin IRS-2 from the hard tick Ixodes ricinus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 1453–1457. [Google Scholar] [CrossRef] [Green Version]
- Meekins, D.A.; Kanost, M.R.; Michel, K. Serpins in arthropod biology. Semin. Cell Dev. Biol. 2017, 62, 105–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.A. The Cell-based model of coagulation. J. Vet. Emerg. Crit. Care 2009, 19, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Masoud, H.M.M. Thrombin inhibitor from the salivary gland of the camel Tick Hyalomma dromedarii. Exp. Appl. Acarol. 2018, 74, 85–97. [Google Scholar] [CrossRef]
- Francischetti, I.M.B.; Sa-Nunes, A.; Mans, B.J.; Santos, I.M.; Ribeiro, J.M.C. The role of saliva in tick feeding. Front. Biosci. 2009, 14, 2051–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumbaut, R.E.; Thiagarajan, P. Platelet Aggregation; Morgan & Claypool Life Sciences, Texas Medical Center: Houston, TX, USA, 2010. [Google Scholar]
- Davì, G.; Patrono, C. Platelet activation and atherothrombosis. N. Engl. J. Med. 2007, 357, 2482–2494. [Google Scholar] [CrossRef] [PubMed]
- Furman, M.I.; Liu, L.; Benoit, S.E.; Becker, R.C.; Barnard, M.R.; Michelson, A.D. The cleaved peptide of the thrombin receptor is a strong platelet agonist. Proc. Natl. Acad. Sci. USA 1998, 95, 3082–3087. [Google Scholar] [CrossRef] [Green Version]
- Al Ghumlas, A.K.; Gader, A.G.M.A. Characterization of the aggregation responses of camel platelets. Vet. Clin. Pathol. 2013, 42, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Abdel Gader, A.G.M.; Al Momen, A.K.M.; Alhaider, A.; Brooks, M.B.; Catalfamo, J.L.; Al Haidary, A.A.; Hussain, M.F. Clotting factor VIII (FVIII) and thrombin generation in camel plasma: A comparative study with humans. Can. J. Vet. Res. 2013, 77, 150–157. [Google Scholar]
- Schechter, N.M.; Plotnick, M.I. Measurement of the kinetic parameters mediating protease-serpin inhibition. Methods 2004, 32, 159–168. [Google Scholar] [CrossRef]
- Gray, E.; Hogwood, J.; Mulloy, B. The anticoagulant and antithrombotic mechanisms of heparin. In Heparin—A Century of Progress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 43–61. [Google Scholar] [CrossRef]
- Varki, A.; Cummings, R.D.; Esko, J.D.; Freeze, H.H.; Stanley, P.; Bertozzi, C.R.; Hart, G.W.; Etzler, M.E. Essentials of Glycobiology, 2nd ed.; Cold Spring Harbor Laboratory Press: Woodbury, NY, USA, 2009; 784p. [Google Scholar]
- Forster, M.; Mulloy, B. Computational approaches to the identification of heparin-binding sites on the surfaces of proteins. Biochem. Soc. Trans. 2006, 34, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Handel, T.M.; Johnson, Z.; Crown, S.E.; Lau, E.K.; Sweeney, M.; Proudfoot, A.E. Regulation of protein function by glycosaminoglycans—As exemplified by chemokines. Annu. Rev. Biochem. 2005, 74, 385–410. [Google Scholar] [CrossRef]
- Fromm, J.R.; Hileman, R.E.; Caldwell, E.E.O.; Weiler, J.M.; Linhardt, R.J. Pattern and spacing of basic amino acids in heparin binding sites. Arch. Biochem. Biophys. 1997, 343, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Huntington, J.A. Crystal structures of protease Nexin-1 in complex with heparin and thrombin suggest a 2-Step recognition mechanism. Blood 2012, 120, 459–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Johnson, D.J.D.; Esmon, C.T.; Huntington, J.A. Structure of the antithrombin-thrombin-heparin ternary complex reveals the antithrombotic mechanism of heparin. Nat. Struct. Mol. Biol. 2004, 11, 857–862. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Batista, I.F.C.; Ramos, O.H.P.; Ventura, J.S.; Junqueira-de-Azevedo, I.L.M.; Ho, P.L.; Chudzinski-Tavassi, A.M. A new Factor Xa inhibitor from Amblyomma cajennense with a Unique Domain Composition. Arch. Biochem. Biophys. 2010, 493, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Bock, S.C.; Wion, K.L.; Vehar, G.A.; Lawn, R.M. Cloning and Expression of the cDNA for Human Antithrombin III. Nucleic Acids Res. 1982, 10, 8113–8125. [Google Scholar] [CrossRef] [PubMed]
- Mulenga, A.; Khumthong, R.; Blandon, M.A. Molecular and Expression Analysis of a Family of the Amblyomma americanum Tick Lospins. J. Exp. Biol. 2007, 210, 3188–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, L.; Radulović, Ž.; Kim, T.; Braz, G.R.C.; Da Silva Vaz, I.; Mulenga, A. Bioinformatic Analyses of Male and Female Amblyomma americanum Tick Expressed Serine Protease Inhibitors (Serpins). Ticks Tick Borne Dis. 2015, 6, 16–30. [Google Scholar] [CrossRef] [Green Version]
- Imamura, S.; Da Silva Vaz Junior, I.; Sugino, M.; Ohashi, K.; Onuma, M. A Serine Protease Inhibitor (Serpin) from Haemaphysalis longicornis as an Anti-Tick Vaccine. Vaccine 2005, 23, 1301–1311. [Google Scholar] [CrossRef]
- Chlastáková, A.; Kotál, J.; Beránková, Z.; Kaščáková, B.; Martins, L.A.; Langhansová, H.; Prudnikova, T.; Ederová, M.; Kutá Smatanová, I.; Kotsyfakis, M.; et al. Iripin-3, a New Salivary Protein Isolated From Ixodes ricinus Ticks, Displays Immunomodulatory and Anti-Hemostatic Properties In Vitro. Front. Immunol. 2021, 12, 626200. [Google Scholar] [CrossRef]
- Schwarz, A.; Von Reumont, B.M.; Erhart, J.; Chagas, A.C.; Ribeiro, J.M.C.; Kotsyfakis, M. De Novo Ixodes ricinus Salivary Gland Transcriptome Analysis Using Two Next-Generation Sequencing Methodologies. FASEB J. 2013, 27, 4745–4756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibelli, A.M.G.; Kim, T.K.; Hill, C.C.; Lewis, L.A.; Bakshi, M.; Miller, S.; Porter, L.; Mulenga, A. A Blood Meal-Induced Ixodes scapularis Tick Saliva Serpin Inhibits Trypsin and Thrombin, and Interferes with Platelet Aggregation and Blood Clotting. Int. J. Parasitol. 2014, 44, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulenga, A.; Tsuda, A.; Onuma, M.; Sugimoto, C. Four Serine Proteinase Inhibitors (Serpin) from the Brown Ear Tick, Rhiphicephalus appendiculatus; cDNA Cloning and Preliminary Characterization. Insect Biochem. Mol. Biol. 2003, 33, 267–276. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elements from the Models | Elements from CD Data | ||
|---|---|---|---|
| Exposed RCL | Inserted RCL | ||
| Total aligned residues (100%) | 372 | 372 | 391 |
| Helix (%) | 29.83 | 30.11 | 54.20 |
| Strand (%) | 30.65 | 33.60 | 21.40 |
| Other (Coil, bridge, Turn) (%) | 39.52 | 36.29 | 24.40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aounallah, H.; Fessel, M.R.; Goldfeder, M.B.; Carvalho, E.; Bensaoud, C.; Chudzinski-Tavassi, A.M.; Bouattour, A.; M’ghirbi, Y.; Faria, F. rDromaserpin: A Novel Anti-Hemostatic Serpin, from the Salivary Glands of the Hard Tick Hyalomma dromedarii. Toxins 2021, 13, 913. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120913
Aounallah H, Fessel MR, Goldfeder MB, Carvalho E, Bensaoud C, Chudzinski-Tavassi AM, Bouattour A, M’ghirbi Y, Faria F. rDromaserpin: A Novel Anti-Hemostatic Serpin, from the Salivary Glands of the Hard Tick Hyalomma dromedarii. Toxins. 2021; 13(12):913. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120913
Chicago/Turabian StyleAounallah, Hajer, Melissa Regina Fessel, Mauricio Barbugiani Goldfeder, Eneas Carvalho, Chaima Bensaoud, Ana Marisa Chudzinski-Tavassi, Ali Bouattour, Youmna M’ghirbi, and Fernanda Faria. 2021. "rDromaserpin: A Novel Anti-Hemostatic Serpin, from the Salivary Glands of the Hard Tick Hyalomma dromedarii" Toxins 13, no. 12: 913. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13120913