Cytotoxic and Genotoxic Effects of Cyanobacterial and Algal Extracts—Microcystin and Retinoic Acid Content



Abstract
:
1. Introduction
2. Results
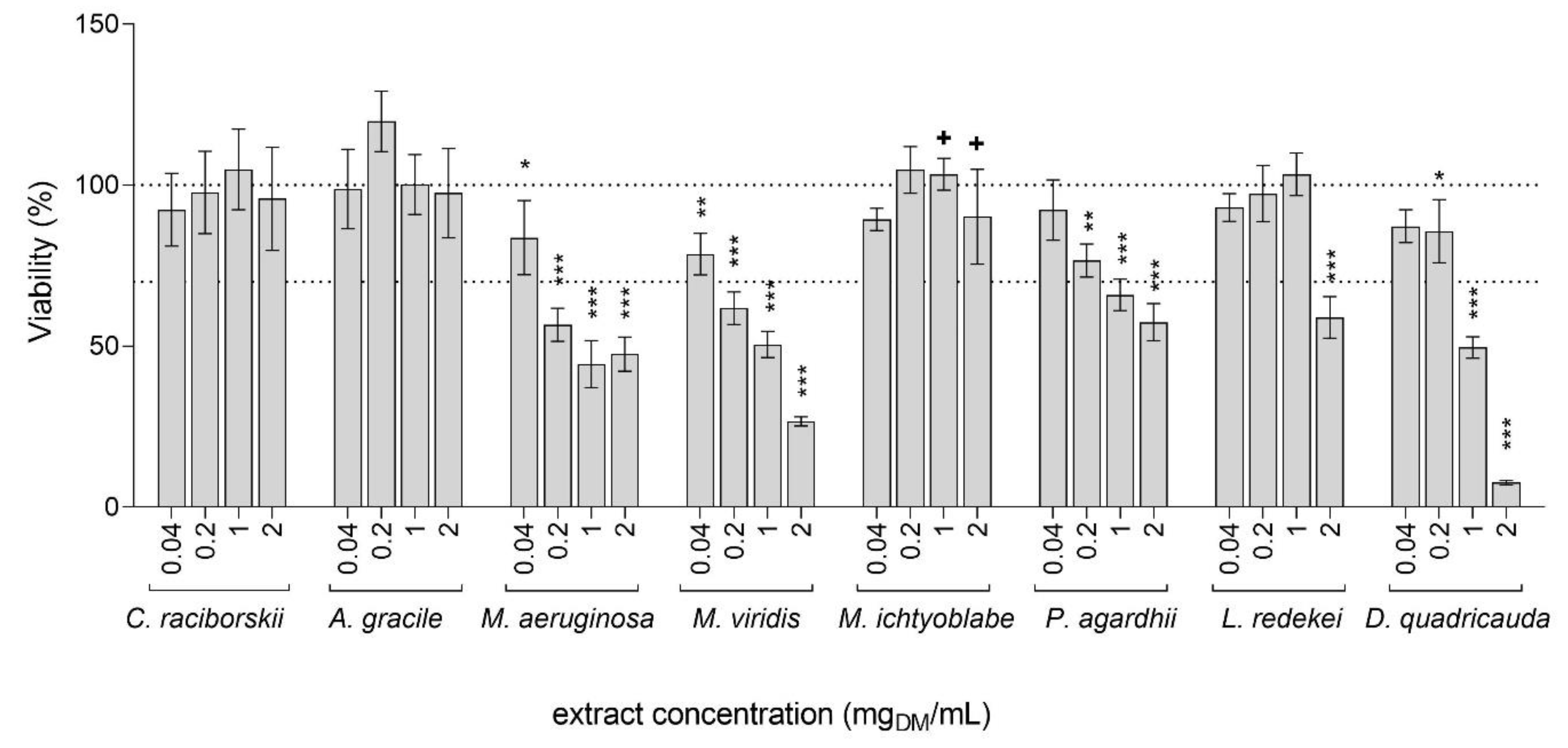
2.1. Cytotoxicity of the Cyanobacterial and Algal Extracts
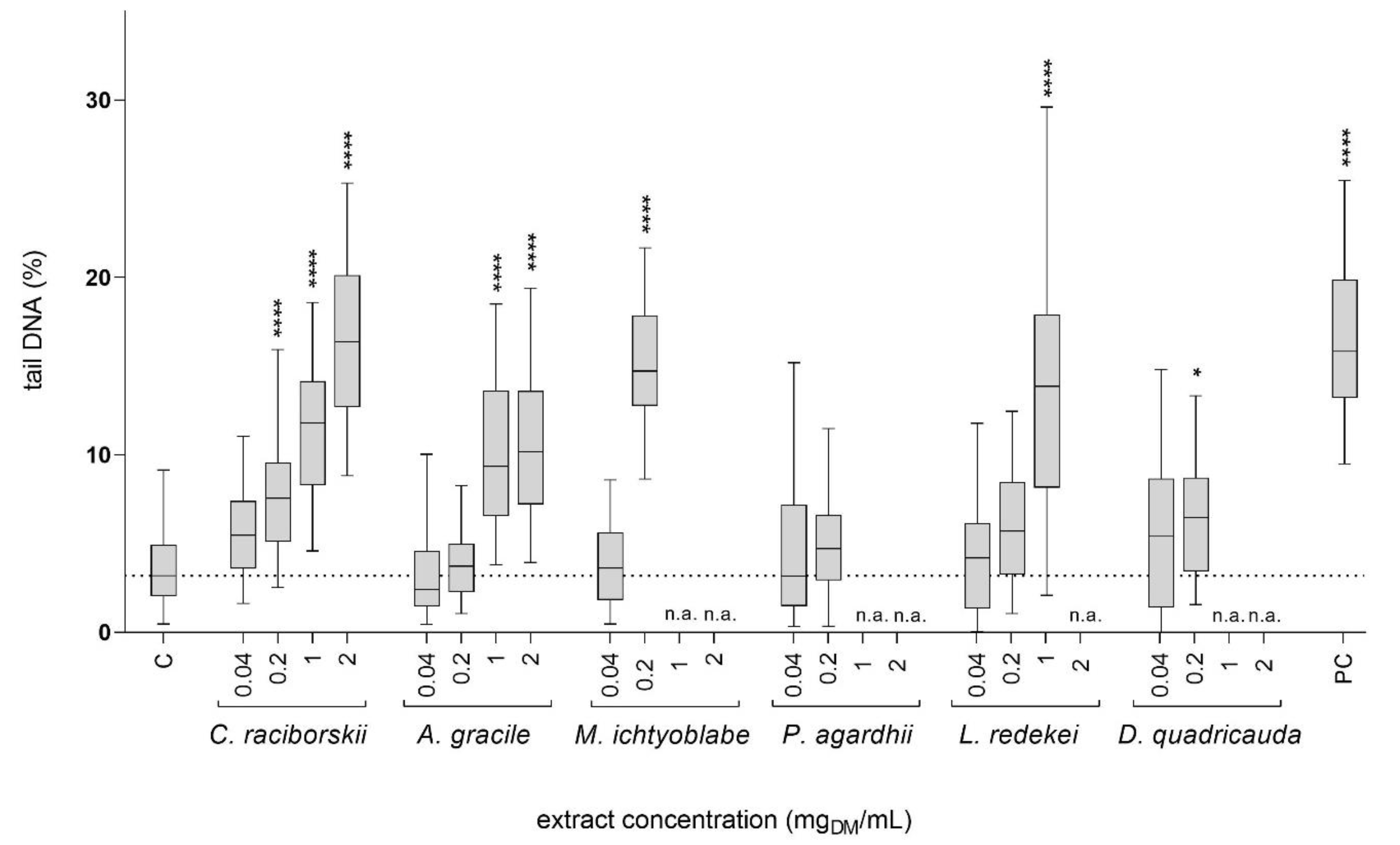
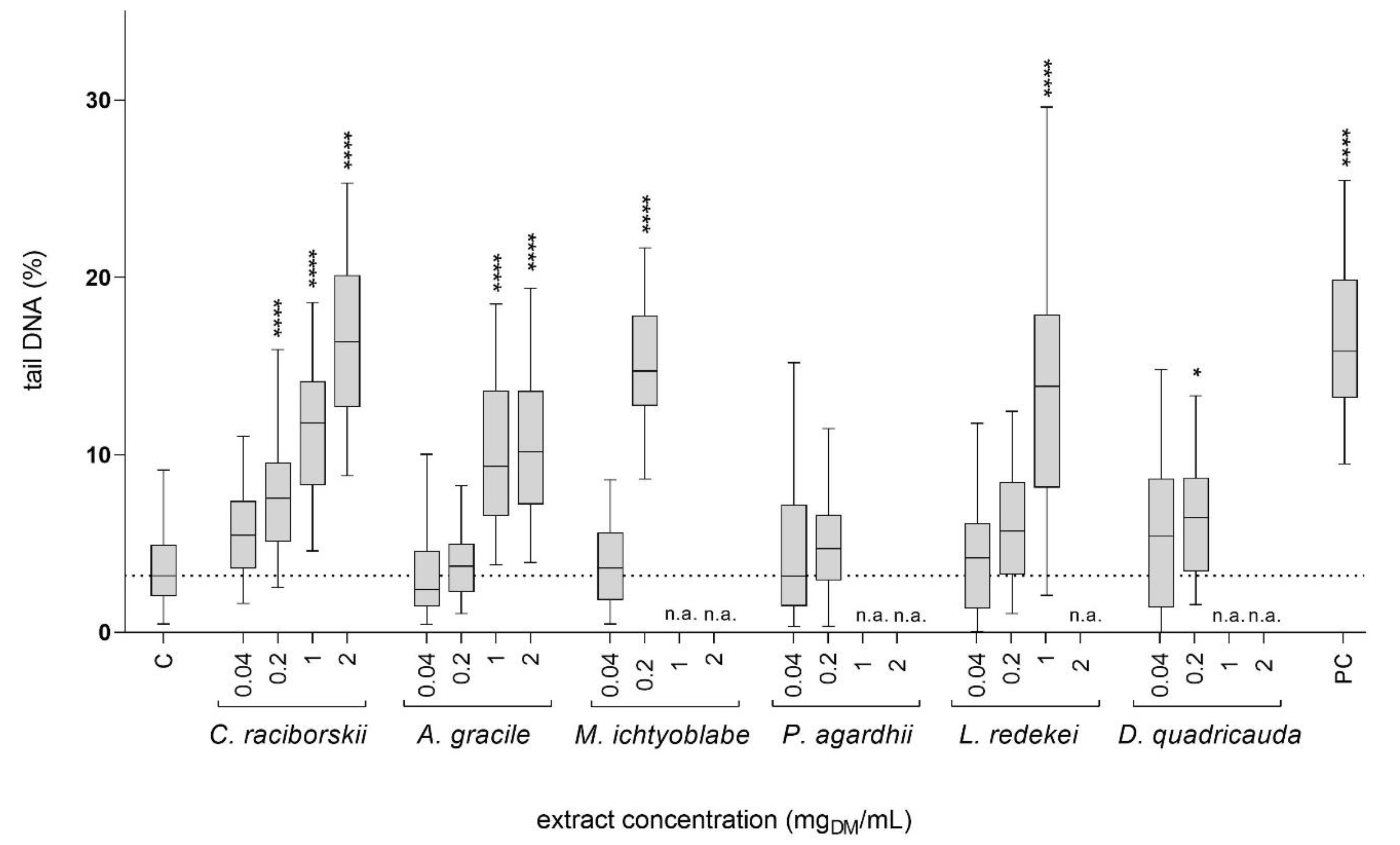
2.2. Genotoxicity of the Cyanobacterial and Algal Extracts—Comet Assay
2.3. Assessment of Genomic Instability Induced by the Cyanobacterial and Algal Extracts—CBMN Assay
2.4. Chemical Analyses
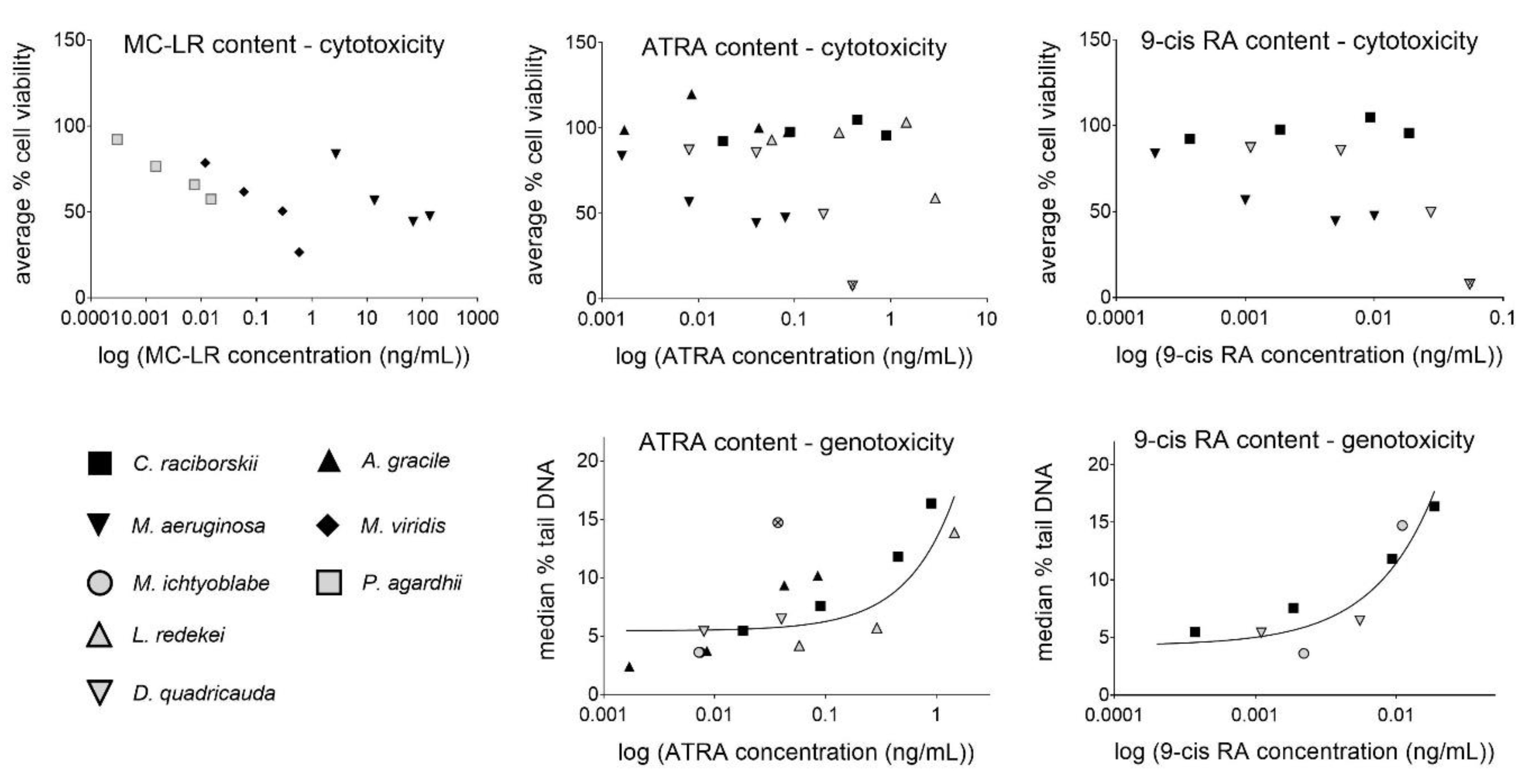
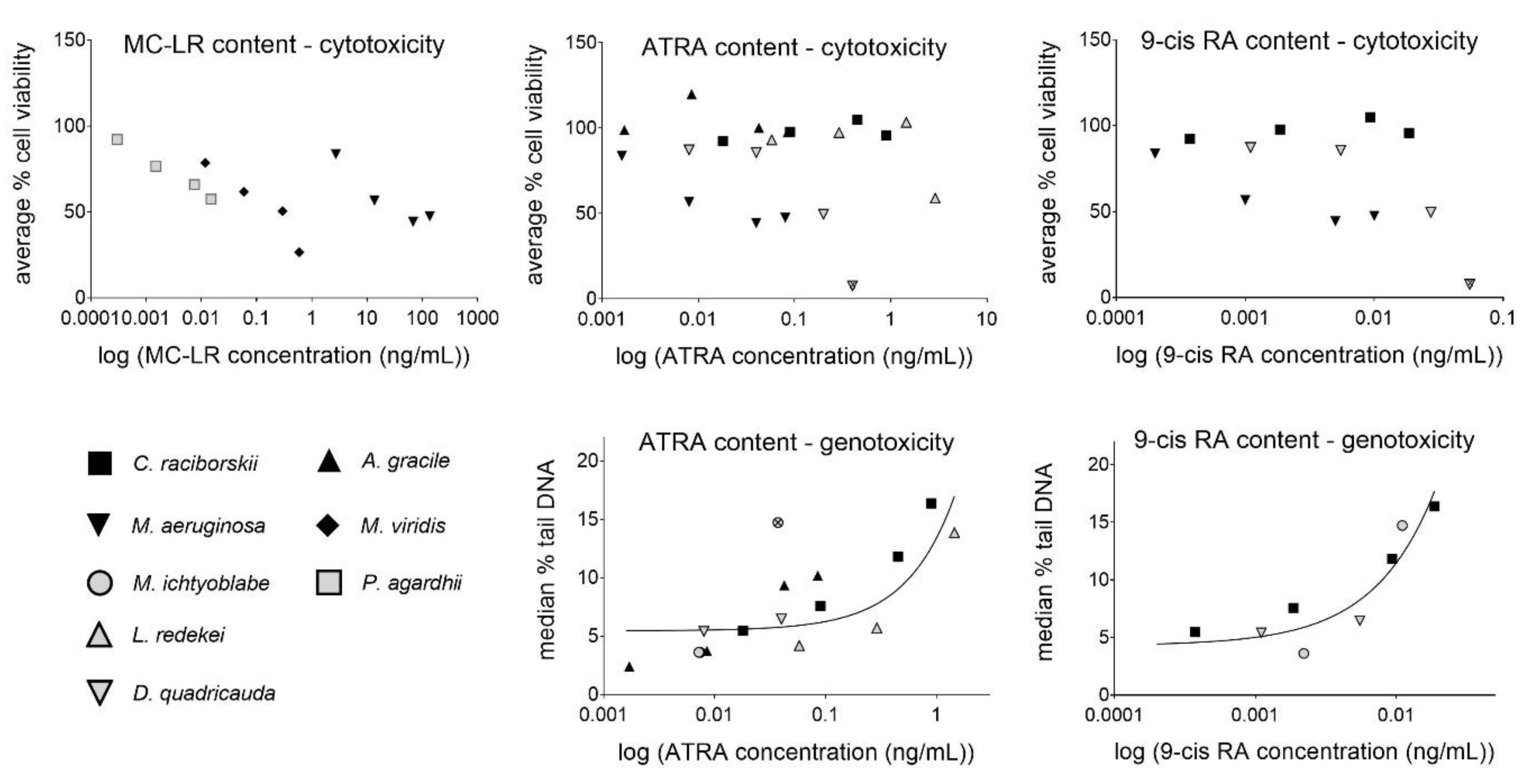
2.5. Assessment of Potential Relationships between MC/RA Content and the Observed Cytotoxic/Genotoxic Effects
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cyanobacterial Cultures and Extraction
5.2. Cell Culture and Treatment
5.3. Cytotoxicity Assay
5.4. Comet Assay
5.5. Cytokinesis-Block Micronucleus (CBMN) Cytome Assay
5.6. Chemical Analyses of Toxins and RAs in the Extracts
5.6.1. Analyses of Microcystins
5.6.2. Analyses of Cylindrospermopsin
5.6.3. Analyses of Retinoic Acids
5.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Scholz, S.N.; Esterhuizen-Londt, M.; Pflugmacher, S. Rise of toxic cyanobacterial blooms in temperate freshwater lakes: Causes, correlations and possible countermeasures. Toxicol. Environ. Chem. 2017, 99, 543–577. [Google Scholar] [CrossRef]
- Codd, G.A.; Meriluoto, J.; Metcalf, J.S. Introduction: Cyanobacteria, Cyanotoxins, Their Human Impact, and Risk Management. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 3–8. ISBN 978-1-119-06868-6. [Google Scholar]
- Lee, J.; Rai, P.K.; Jeon, Y.J.; Kim, K.-H.; Kwon, E.E. The role of algae and cyanobacteria in the production and release of odorants in water. Environ. Pollut. 2017, 227, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Svirčev, Z.; Drobac, D.; Tokodi, N.; Mijović, B.; Codd, G.A.; Meriluoto, J. Toxicology of microcystins with reference to cases of human intoxications and epidemiological investigations of exposures to cyanobacteria and cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef]
- Chorus, I. Introduction: Cyanotoxins—Research for Environmental Safety and Human Health. In Cyanotoxins: Occurrence, Causes, Consequences; Chorus, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; ISBN 978-3-642-59514-1. [Google Scholar]
- Bláhová, L.; Babica, P.; Adamovský, O.; Kohoutek, J.; Maršálek, B.; Bláha, L. Analyses of cyanobacterial toxins (microcystins, cylindrospermopsin) in the reservoirs of the Czech Republic and evaluation of health risks. Environ. Chem. Lett. 2008, 6, 223–227. [Google Scholar] [CrossRef]
- Bláhová, L.; Babica, P.; Maršálková, E.; Maršálek, B.; Bláha, L. Concentrations and Seasonal Trends of Extracellular Microcystins in Freshwaters of the Czech Republic—Results of the National Monitoring Program. CLEAN—Soil Air Water 2007, 35, 348–354. [Google Scholar] [CrossRef]
- Žegura, B.; Štraser, A.; Filipič, M. Genotoxicity and potential carcinogenicity of cyanobacterial toxins—A review. Mutat. Res./Rev. Mutat. Res. 2011, 727, 16–41. [Google Scholar] [CrossRef]
- Svirčev, Z.; Lalić, D.; Bojadžija Savić, G.; Tokodi, N.; Drobac Backović, D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global Geographical and Historical Overview of Cyanotoxin Distribution and Cyanobacterial Poisonings; Springer: Berlin/Heidelberg, Germany, 2019; Volume 93, ISBN 0123456789. [Google Scholar]
- Zegura, B. An Overview of the Mechanisms of Microcystin-LR Genotoxicity and Potential Carcinogenicity. Mini Rev. Med. Chem. 2016, 16, 1042–1062. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. toxins Structural Diversity, Characterization and Toxicology of Microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef] [Green Version]
- IARC. Ingested Nitrate and Nitrite, and Cyanobacterial Peptide Toxins; IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, No. 94; International Agency for Research on Cancer: Lyon, France, 2010. [Google Scholar]
- Runnegar, M.T.; Xie, C.; Snider, B.B.; Wallace, G.A.; Weinreb, S.M.; Kuhlenkamp, J. In Vitro Hepatotoxicity of the Cyanobacterial Alkaloid Cylindrospermopsin and Related Synthetic Analogues. Toxicol. Sci. 2002, 67, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichardo, S.; Cameán, A.M.; Jos, A. In vitro toxicological assessment of cylindrospermopsin: A review. Toxins 2017, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrouzek, P.; Kapuścik, A.; Vacek, J.; Voráčová, K.; Paichlová, J.; Kosina, P.; Voloshko, L.; Ventura, S.; Kopecký, J. Cytotoxicity evaluation of large cyanobacterial strain set using selected human and murine in vitro cell models. Ecotoxicol. Environ. Saf. 2016, 124, 177–185. [Google Scholar] [CrossRef]
- Smutná, M.; Babica, P.; Jarque, S.; Hilscherová, K.; Maršálek, B.; Haeba, M.; Bláha, L. Acute, chronic and reproductive toxicity of complex cyanobacterial blooms in Daphnia magna and the role of microcystins. Toxicon 2014, 79, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Sieroslawska, A. Assessment of the mutagenic potential of cyanobacterial extracts and pure cyanotoxins. Toxicon 2013, 74, 76–82. [Google Scholar] [CrossRef]
- Ding, W.-X.; Shen, H.-M.; Zhu, H.-G.; Lee, B.-L.; Ong, C.-N. Genotoxicity of microcystic cyanobacteria extract of a water source in China. Mutat. Res. Toxicol. Environ. Mutagen. 1999, 442, 69–77. [Google Scholar] [CrossRef]
- Bláha, L.; Babica, P.; Hilscherová, K.; Upham, B.L. Inhibition of gap-junctional intercellular communication and activation of mitogen-activated protein kinases by cyanobacterial extracts—Indications of novel tumor-promoting cyanotoxins? Toxicon 2010, 55, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Funari, E.; Testai, E. Human Health Risk Assessment Related to Cyanotoxins Exposure. Crit. Rev. Toxicol. 2008, 38, 97–125. [Google Scholar] [CrossRef]
- Jonas, A.; Scholz, S.; Fetter, E.; Sychrova, E.; Novakova, K.; Ortmann, J.; Benisek, M.; Adamovsky, O.; Giesy, J.P.; Hilscherova, K. Endocrine, teratogenic and neurotoxic effects of cyanobacteria detected by cellular in vitro and zebrafish embryos assays. Chemosphere 2015, 120, 321–327. [Google Scholar] [CrossRef]
- Priebojová, J.; Hilscherová, K.; Procházková, T.; Sychrová, E.; Smutná, M. Intracellular and extracellular retinoid-like activity of widespread cyanobacterial species. Ecotoxicol. Environ. Saf. 2018, 150, 312–319. [Google Scholar] [CrossRef]
- Javůrek, J.; Sychrová, E.; Smutná, M.; Bittner, M.; Kohoutek, J.; Adamovský, O.; Nováková, K.; Smetanová, S.; Hilscherová, K. Retinoid compounds associated with water blooms dominated by Microcystis species. Harmful Algae 2015, 47, 116–125. [Google Scholar] [CrossRef]
- Pipal, M.; Priebojova, J.; Koci, T.; Blahova, L.; Smutna, M.; Hilscherova, K. Field cyanobacterial blooms producing retinoid compounds cause teratogenicity in zebra fi sh embryos. Chemosphere 2020, 241, 125061. [Google Scholar] [CrossRef] [PubMed]
- Sehnal, L.; Procházková, T.; Smutná, M.; Kohoutek, J.; Lepšová-Skácelová, O.; Hilscherová, K. Widespread occurrence of retinoids in water bodies associated with cyanobacterial blooms dominated by diverse species. Water Res. 2019, 156, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Kaya, K.; Sano, T. Cyanobacterial Retinoids. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 173–178. [Google Scholar]
- Alakhras, R.S.; Stephanou, G.; Demopoulos, N.A.; Nikolaropoulos, S.S. Genotoxicity of all-trans retinoic acid (ATRA) and its steroidal analogue EA-4 in human lymphocytes and mouse cells in vitro. Cancer Lett. 2011, 306, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Balaban, F.; Ates-Alagoz, Z.; Buyukbingol, E.; Iscan, M. Genotoxicity studies on benzimidazole retinoids. Pharmazie 2005, 60, 861–868. [Google Scholar] [PubMed]
- de Almeida Vasconcelos Fonseca, E.M. All-trans and 9-cis retinoic acids, retinol and -carotene chemopreventive activities during the initial phases of hepatocarcinogenesis involve distinct actions on glutathione S-transferase positive preneoplastic lesions remodeling and DNA damage. Carcinogenesis 2005, 26, 1940–1946. [Google Scholar] [CrossRef] [PubMed]
- Hamid, R.; Rotshteyn, Y.; Rabadi, L.; Parikh, R.; Bullock, P. Comparison of alamar blue and MTT assays for high through-put screening. Toxicol. Vitr. 2004, 18, 703–710. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Hercog, K.; Štampar, M.; Filipič, M.; Cameán, A.M.; Jos, Á.; Žegura, B. Genotoxic Effects of Cylindrospermopsin, Microcystin-LR and Their Binary Mixture in Human Hepatocellular Carcinoma (HepG2) Cell Line. Toxins 2020, 12, 778. [Google Scholar] [CrossRef]
- Hercog, K.; Maisanaba, S.; Filipič, M.; Jos, Á.; Cameán, A.M.; Žegura, B. Genotoxic potential of the binary mixture of cyanotoxins microcystin-LR and cylindrospermopsin. Chemosphere 2017, 189, 319–329. [Google Scholar] [CrossRef]
- Hercog, K.; Maisanaba, S.; Filipič, M.; Sollner-Dolenc, M.; Kač, L.; Žegura, B. Genotoxic activity of bisphenol A and its analogues bisphenol S, bisphenol F and bisphenol AF and their mixtures in human hepatocellular carcinoma (HepG2) cells. Sci. Total Environ. 2019, 687, 267–276. [Google Scholar] [CrossRef]
- Westerink, W.M.A.; Schoonen, W.G.E.J. Cytochrome P450 enzyme levels in HepG2 cells and cryopreserved primary human hepatocytes and their induction in HepG2 cells. Toxicol. Vitr. 2007, 21, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Pi, F.; Wang, J.; Ji, J.; Sun, X. New insights into cytotoxicity induced by microcystin-LR, estradiol, and ractopamine with mathematical models: Individual and combined effects. Chemosphere 2017, 168, 223–233. [Google Scholar] [CrossRef]
- Lundqvist, J.; Pekar, H.; Oskarsson, A. Microcystins activate nuclear factor erythroid 2-related factor 2 (Nrf2) in human liver cells in vitro—Implications for an oxidative stress induction by microcystins. Toxicon 2017, 126, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Al-Sheddi, E.S.; Al-Oqail, M.M.; Saquib, Q.; Siddiqui, M.A.; Musarrat, J.; Al-Khedhairy, A.A.; Farshori, N.N. Novel All Trans-Retinoic Acid Derivatives: Cytotoxicity, Inhibition of Cell Cycle Progression and Induction of Apoptosis in Human Cancer Cell Lines. Molecules 2015, 20, 8181–8197. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Hu, C.; Xu, L.; Cui, J.; Tao, L.; Gong, M.; Wang, Y.; He, Y.; He, T.; Bi, Y. All-trans-retinoic acid inhibits the malignant behaviors of hepatocarcinoma cells by regulating autophagy. Am. J. Transl. Res. 2020, 12, 6793. [Google Scholar] [PubMed]
- Um, S.-J.; Sin, H.-S.; Han, H.-S.; Kwon, Y.-J.; Kim, E.-J.; Park, S.-H.; Kim, S.-Y.; Bae, T.-S.; Park, J.-S.; Rho, Y.-S. Potent cytotoxic effects of novel retinamide derivatives in ovarian cancer cells. Biol. Pharm. Bull. 2003, 26, 1412–1417. [Google Scholar] [CrossRef] [Green Version]
- Abramsson-Zetterberg, L.; Sundh, U.B.; Mattsson, R. Cyanobacterial extracts and microcystin-LR are inactive in the micronucleus assay in vivo and in vitro. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2010, 699, 5–10. [Google Scholar] [CrossRef]
- Scarlett, K.R.; Kim, S.; Lovin, L.M.; Chatterjee, S.; Scott, J.T.; Brooks, B.W. Global scanning of cylindrospermopsin: Critical review and analysis of aquatic occurrence, bioaccumulation, toxicity and health hazards. Sci. Total Environ. 2020, 738, 139807. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Occurrence and toxicity of microcystin congeners other than MC-LR and MC-RR: A review. Food Chem. Toxicol. 2019, 125, 106–132. [Google Scholar] [CrossRef]
- Žegura, B.; Sedmak, B.; Filipič, M. Microcystin-LR induces oxidative DNA damage in human hepatoma cell line HepG2. Toxicon 2003, 41, 41–48. [Google Scholar] [CrossRef]
- Žegura, B.; Filipič, M. Application of In Vitro Comet Assay for Genotoxicity Testing. In Optimization in Drug Discovery; Humana Press: Totowa, NJ, USA, 2004; pp. 301–313. [Google Scholar]
- Palus, J.; Dziubałtowska, E.; Stańczyk, M.; Lewińska, D.; Mankiewicz-Boczek, J.; Izydorczyk, K.; Bonisławska, A.; Jurczak, T.; Zalewski, M.; Wąsowicz, W. Biomonitoring Of Cyanobacterial Blooms in Polish Water Reservoir And The Cytotoxicity and Genotoxicity Of Selected Cyanobacterial Extracts. Int. J. Occup. Med. Environ. Health 2007, 20, 48–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. In Proceedings of the Environmental and Molecular Mutagenesis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2000; Volume 35, pp. 206–221. [Google Scholar]
- OECD. Test No. 487: In Vitro Mammalian Cell Micronucleus Test. In OECD Guideline for the Testing of Chemicals; OECD: Paris, France, 2016. [Google Scholar]
- Štraser, A.; Filipič, M.; Žegura, B. Genotoxic effects of the cyanobacterial hepatotoxin cylindrospermopsin in the HepG2 cell line. Arch. Toxicol. 2011, 85, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M. The in vitro micronucleus technique. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2000, 455, 81–95. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Order | Source | Place of Origin |
|---|---|---|---|
| Cylindrospermopsis raciborskii | Nostocales | SAG 1.97 | Lake Balaton, Hungary |
| Aphanizomenon gracile | Nostocales | RCX 06 | Lough Neah, Ireland |
| Microcystis aeruginosa | Chroococcales | PCC 7806 | Braakman Reservoir, The Netherlands |
| Microcystis viridis | Chroococcales | PC II21LiP10/IX | Lipno, Czech Republic |
| Microcystis ichtyoblabe | Chroococcales | PC II42LiP10/V | Lipno, Czech Republic |
| Planktothrix agardhii | Oscillatoriales | CCALA 159 | Plon lake, Germany |
| Limnothrix redekei | Synechococcales | SAG 3.89 | Edebergsee at Plön, Germany |
| Desmodesmus quadricauda | Sphaeropleales | CCALA 463 | Greifswald, Germany |
| Sample | MNed Cells/103 BNC | Fold Induction | NDI |
|---|---|---|---|
| C. raciborskii | 32.0 ± 4.0 | 1.07 | 1.8 ± 0.04 |
| A. gracile | 32.7 ± 4.9 | 1.09 | 1.8 ± 0.1 |
| M. aeruginosa | 40.0 ± 10.8 | 1.33 | 1.7 ± 0.1 |
| M. viridis | 33.3 ± 2.1 | 1.11 | 1.8 ± 0.1 |
| M. ichtyoblabe | 30.7 ± 9.5 | 1.02 | 1.7 ± 0.1 |
| P. agardhii | 32.7 ± 10.1 | 1.09 | 1.8 ± 0.01 |
| L. redekei | 35.3 ± 7.2 | 1.18 | 1.8 ± 0.1 |
| D. quadricauda | 31.7 ± 3.5 | 1.06 | 1.8 ± 0.04 |
| Vehicle control | 30.0 ± 2.0 | 1 | 1.8 ± 0.1 |
| Etoposide (l μg/mL) | 199.7 ± 35.5 * | 6.65 | 1.2 ± 0.1 * |
| Sample | Microcystins (ng/gDM) | Cylindrospermopsin (ng/gDM) | Retinoic Acids (ng/gDM) | |||
|---|---|---|---|---|---|---|
| MC-RR | MC-YR | MC-LR | ATRA | 9cis-RA | ||
| C. raciborskii | <0.6 | <2.5 | <10 | <0.25 | 450 | 9.3 |
| A. gracile | <0.6 | <2.5 | <10 | <0.25 | 42.5 | <5 |
| M. aeruginosa | 23.65 | <2.5 | 68,000 | <0.25 | 40 | 5.5 |
| M. viridis | 28.8 | 12.8 | 295.5 | <0.25 | <7.5 | <5 |
| M. ichtyoblabe | 8.7 | <2.5 | 50.5 | <0.25 | 180 | 55 |
| P. agardhii | <0.6 | 10,750 | 7.5 | <0.25 | <7.5 | <5 |
| L. redekei | <0.6 | <2.5 | <10 | <0.25 | 1450 | <5 |
| D. quadricauda | <0.6 | <2.5 | <10 | <0.25 | 200 | 27.5 |
| Linear Regression | Cytotoxicity (Average % Viability) | Genotoxicity (Median Tail DNA%) | ||||
|---|---|---|---|---|---|---|
| R2 | p | n | R2 | p | n | |
| ATRA content | 0.02 | 0.5686 | 20 | 0.63 | 0.0004 | 15 |
| 9cis-RA content | 0.10 | 0.3253 | 12 | 0.88 | 0.0002 | 9 |
| Combined RAs content | 0.02 | 0.5584 | 20 | 0.64 | 0.0004 | 15 |
| MC-LR content | 0.14 | 0.2011 | 13 | n.a. | n.a. | 6 |
| MC-RR content | n.a. | n.a. | 9 | n.a. | n.a. | 2 |
| MC-YR content | n.a | n.a | 9 | n.a. | n.a. | 4 |
| Combined MCs content | 0.18 | 0.1122 | 13 | n.a. | n.a. | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bittner, M.; Štern, A.; Smutná, M.; Hilscherová, K.; Žegura, B. Cytotoxic and Genotoxic Effects of Cyanobacterial and Algal Extracts—Microcystin and Retinoic Acid Content. Toxins 2021, 13, 107. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020107
Bittner M, Štern A, Smutná M, Hilscherová K, Žegura B. Cytotoxic and Genotoxic Effects of Cyanobacterial and Algal Extracts—Microcystin and Retinoic Acid Content. Toxins. 2021; 13(2):107. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020107
Chicago/Turabian StyleBittner, Michal, Alja Štern, Marie Smutná, Klára Hilscherová, and Bojana Žegura. 2021. "Cytotoxic and Genotoxic Effects of Cyanobacterial and Algal Extracts—Microcystin and Retinoic Acid Content" Toxins 13, no. 2: 107. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020107