Genotypic and Phenotypic Characterization of Staphylococcus aureus Isolates from the Respiratory Tract in Mechanically-Ventilated Patients

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Patients and Strains
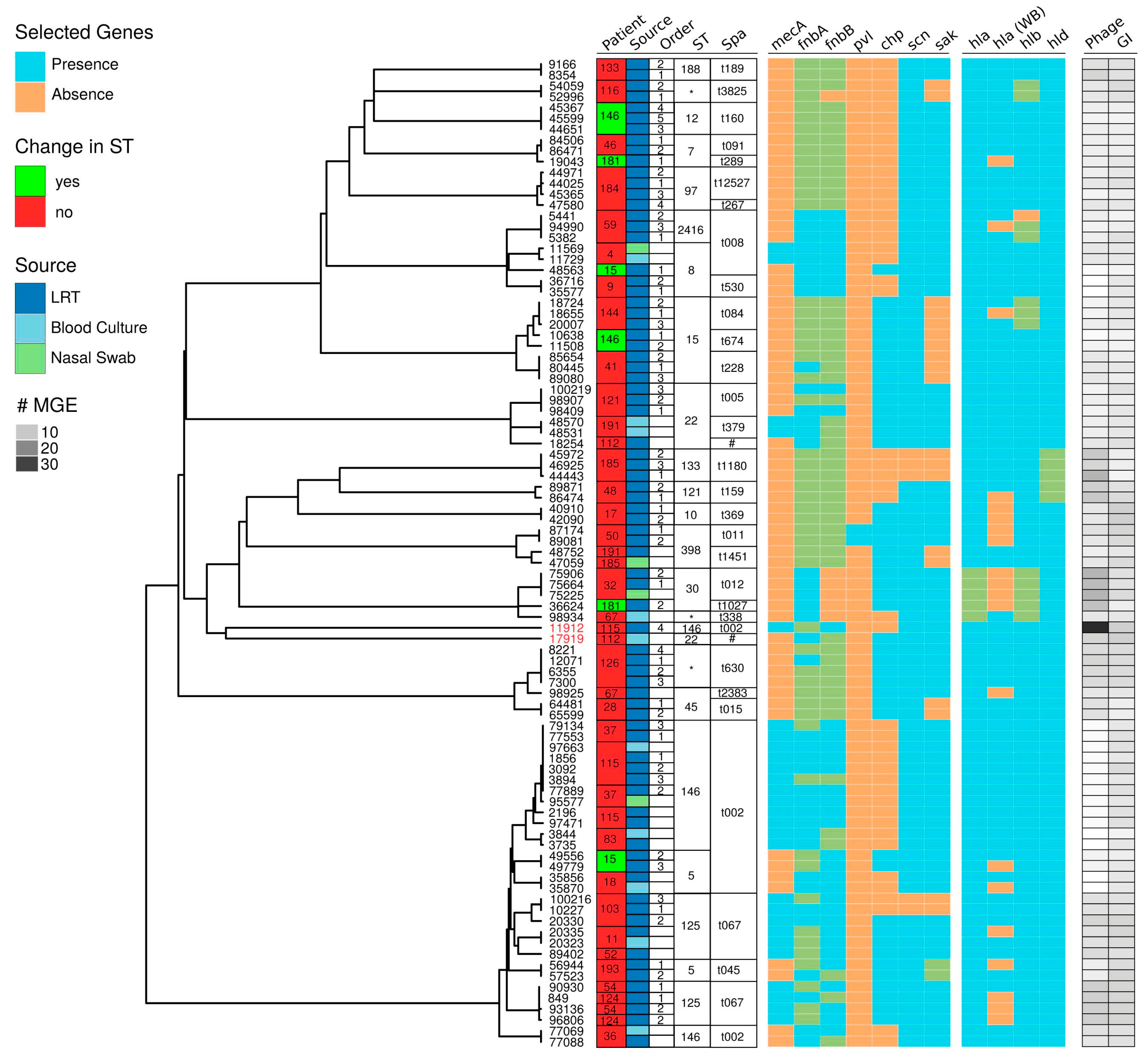
2.2. Genomic Characterization
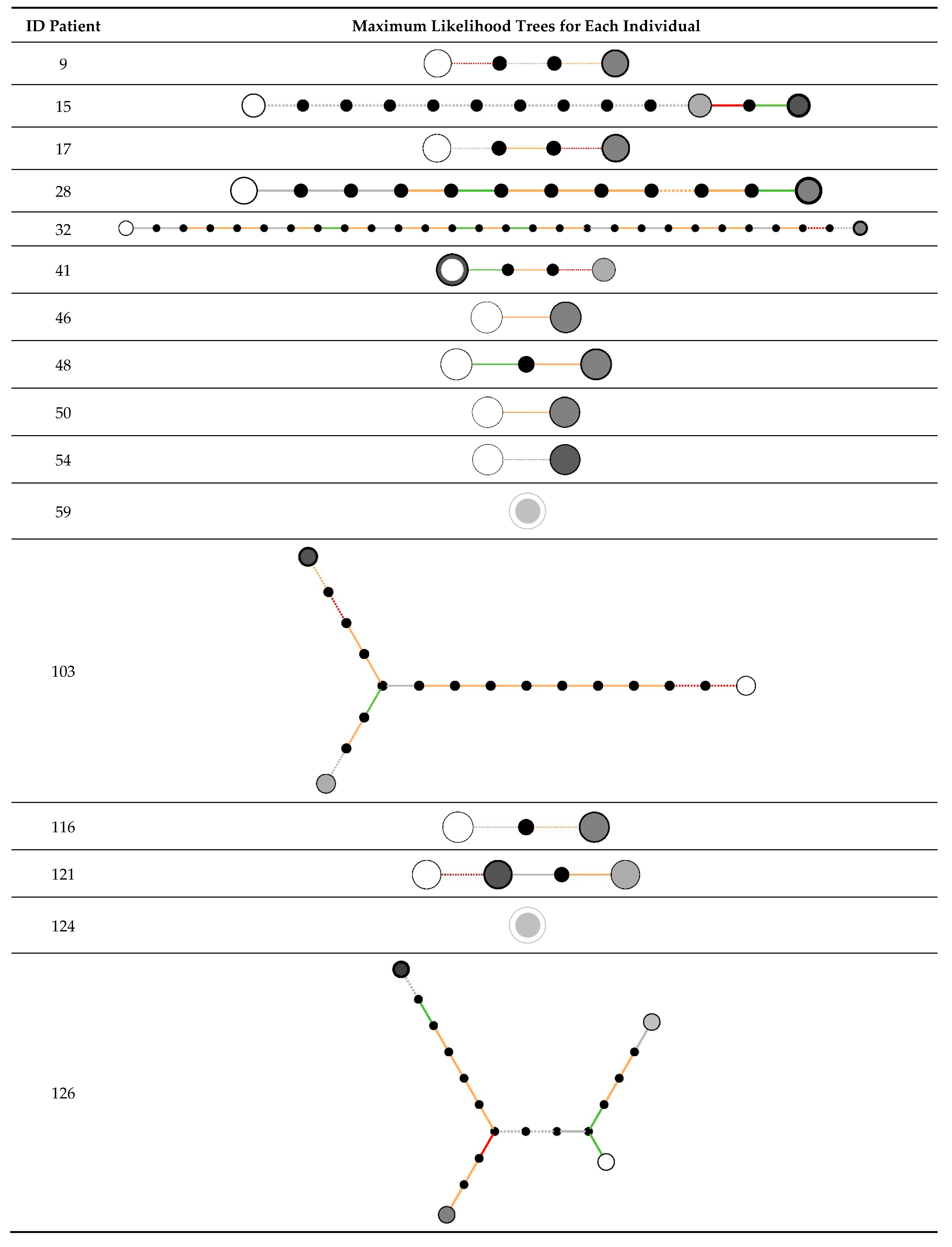
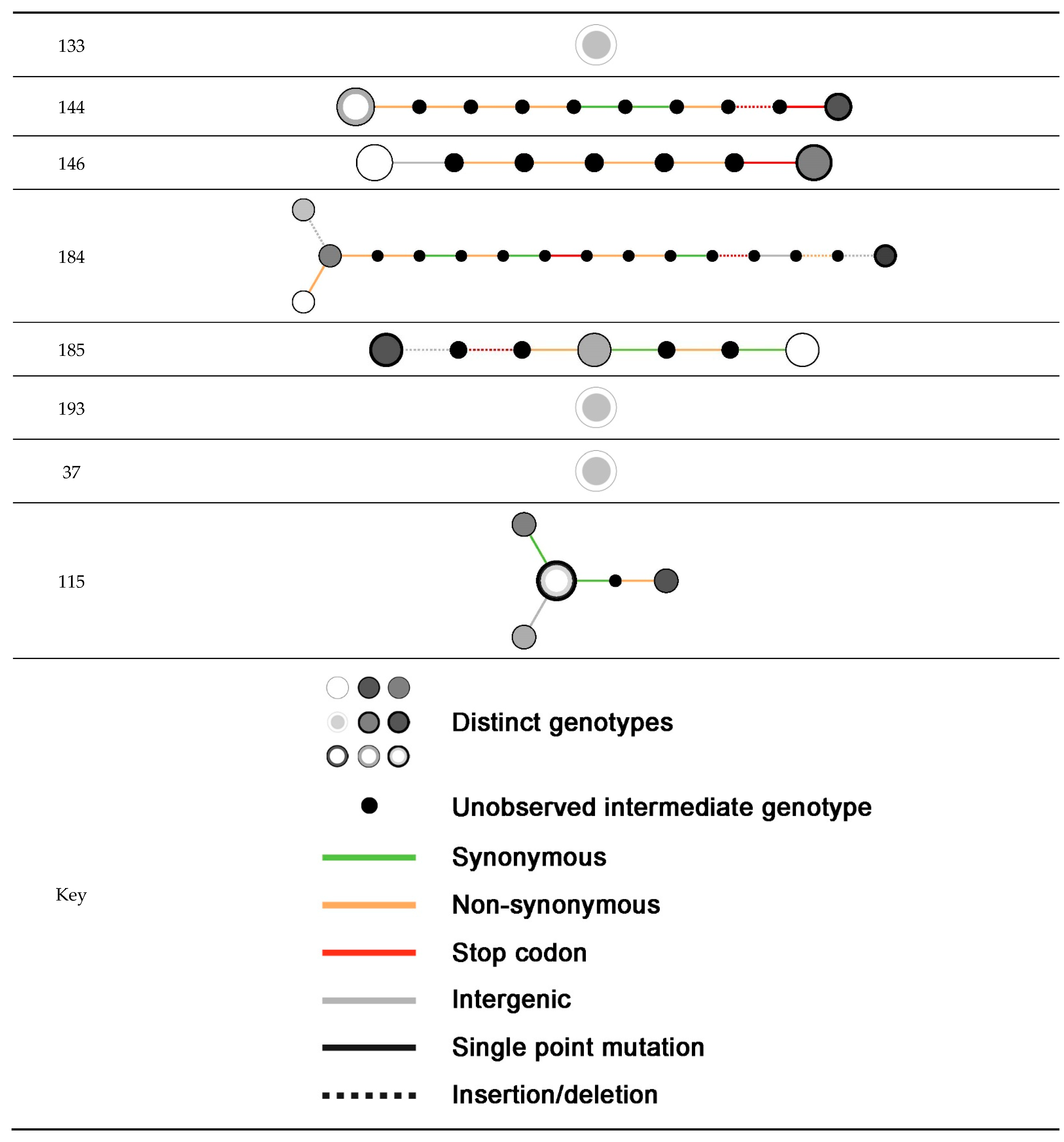
2.3. Within-Host Variation
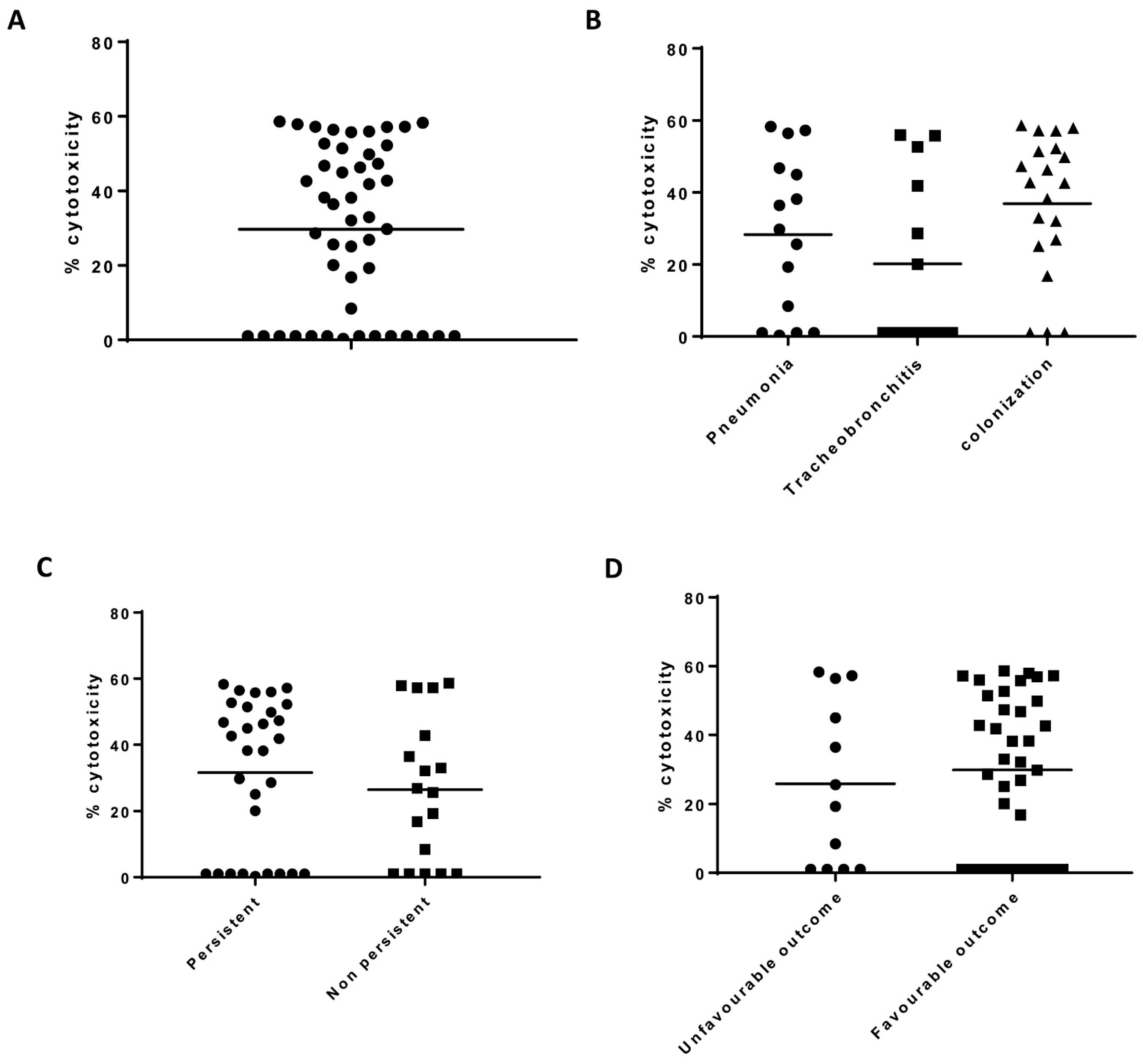
2.4. THP-1 Cytotoxicity
2.5. Fn Binding Capacity
2.6. Alpha Toxin Detection by Western Blot
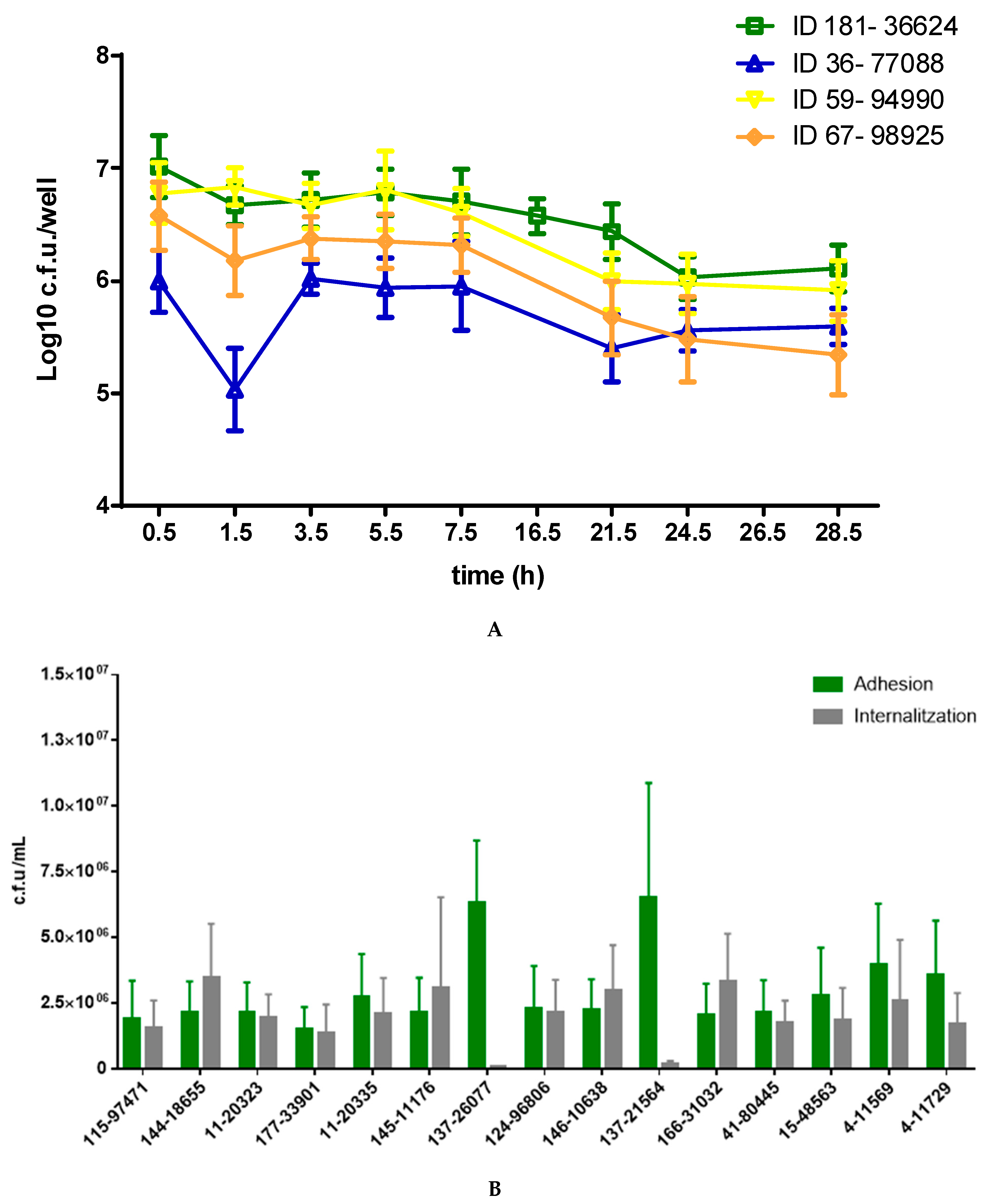
2.7. Invasion and Persistence Assay
3. Discussion
4. Material and Methods
4.1. Patients and Strains
4.2. Whole Genome Sequencing
4.3. Genomic Characterization
4.4. Within-Host Variation Analysis
4.5. Phenotypic Characterization
4.6. THP-1 Cytotoxicity Assay
4.7. Fibronectin Binding Assay
4.8. Detection of Secreted Hla
4.9. Invasion Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, D.; Harper, L.; Shopsin, B.; Torres, V.J. Staphylococcus aureus pathogenesis in diverse host environments. Pathog. Dis. 2017, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dastgheyb, S.S.; Otto, M. Staphylococcal adaptation to diverse physiologic niches: An overview of transcriptomic and phenotypic changes in different biological environments. Future Microbiol. 2015, 10, 1981–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalil, A.C.; Metersky, M.L.; Klompas, M.; Muscedere, J.; Sweeney, D.A.; Palmer, L.B.; Napolitano, L.M.; O’Grady, N.P.; Bartlett, J.G.; Carratala, J.; et al. Management of Adults with Hospital-acquired and Ventilator-associated Pneumonia: 2016 Clinical Practice Guidelines by the Infectious Diseases Society of America and the American Thoracic Society. Clin. Infect. Dis. 2016, 63, e61–e111. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Pavia, A.T. Editorial Commentary: The Modern Quest for the “Holy Grail” of Pneumonia Etiology. Clin. Infect. Dis. 2016, 62, 826–828. [Google Scholar] [CrossRef] [Green Version]
- Prat, C.; Lacoma, A. Bacteria in the respiratory tract—How to treat? Or do not treat? Int. J. Infect. Dis. 2016, 51, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Paling, F.P.; Hazard, D.; Bonten, M.J.M.; Goossens, H.; Jafri, H.S.; Malhotra-Kumar, S.; Sifakis, F.; Weber, S.; Kluytmans, J.; Team, A.-I.S. Association of Staphylococcus aureus Colonization and Pneumonia in the Intensive Care Unit. JAMA Netw. Open 2020, 3, e2012741. [Google Scholar] [CrossRef]
- Kuehl, R.; Morata, L.; Meylan, S.; Mensa, J.; Soriano, A. When antibiotics fail: A clinical and microbiological perspective on antibiotic tolerance and persistence of Staphylococcus aureus. J. Antimicrob. Chemother. 2020, 75, 1071–1086. [Google Scholar] [CrossRef]
- Lacoma, A.; Gomes-Fernandes, M.; Mesalles, E.; Armestar, F.; Prat, C. Persistent isolation of Staphylococcus aureus in mechanically ventilated patients: Impact of host-pathogen factors on outcome. Arch. Bronconeumol. 2019, 55, 158–160. [Google Scholar] [CrossRef]
- Tan, X.; Coureuil, M.; Ramond, E.; Euphrasie, D.; Dupuis, M.; Tros, F.; Meyer, J.; Nemazanyy, I.; Chhuon, C.; Guerrera, I.C.; et al. Chronic Staphylococcus aureus Lung Infection Correlates With Proteogenomic and Metabolic Adaptations Leading to an Increased Intracellular Persistence. Clin. Infect. Dis. 2019, 69, 1937–1945. [Google Scholar] [CrossRef]
- Cullen, L.; McClean, S. Bacterial Adaptation during Chronic Respiratory Infections. Pathogens 2015, 4, 66–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, J.; Stelzner, K.; Rudel, T.; Fraunholz, M. Inside job: Staphylococcus aureus host-pathogen interactions. Int. J. Med. Microbiol. 2018, 308, 607–624. [Google Scholar] [CrossRef] [PubMed]
- Lacoma, A.; Cano, V.; Moranta, D.; Regueiro, V.; Dominguez-Villanueva, D.; Laabei, M.; Gonzalez-Nicolau, M.; Ausina, V.; Prat, C.; Bengoechea, J.A. Investigating intracellular persistence of Staphylococcus aureus within a murine alveolar macrophage cell line. Virulence 2017, 8, 1761–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priest, N.K.; Rudkin, J.K.; Feil, E.J.; van den Elsen, J.M.; Cheung, A.; Peacock, S.J.; Laabei, M.; Lucks, D.A.; Recker, M.; Massey, R.C. From genotype to phenotype: Can systems biology be used to predict Staphylococcus aureus virulence? Nat. Rev. Microbiol. 2012, 10, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus aureus toxins. Curr. Opin. Microbiol. 2014, 17, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Hook, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Peacock, S.J.; Day, N.P.; Thomas, M.G.; Berendt, A.R.; Foster, T.J. Clinical isolates of Staphylococcus aureus exhibit diversity in fnb genes and adhesion to human fibronectin. J. Infect. 2000, 41, 23–31. [Google Scholar] [CrossRef]
- Edwards, A.M.; Potts, J.R.; Josefsson, E.; Massey, R.C. Staphylococcus aureus host cell invasion and virulence in sepsis is facilitated by the multiple repeats within FnBPA. PLoS Pathog. 2010, 6, e1000964. [Google Scholar] [CrossRef] [Green Version]
- Seilie, E.S.; Wardenburg, J.B. Staphylococcus aureus pore-forming toxins: The interface of pathogen and host complexity. Semin. Cell Dev. Biol. 2017, 72, 101–116. [Google Scholar] [CrossRef]
- Berube, B.J.; Wardenburg, J.B. Staphylococcus aureus alpha-toxin: Nearly a century of intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef] [Green Version]
- Sause, W.E.; Buckley, P.T.; Strohl, W.R.; Lynch, A.S.; Torres, V.J. Antibody-Based Biologics and Their Promise to Combat Staphylococcus aureus Infections. Trends Pharmacol. Sci. 2016, 37, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Vlaeminck, J.; Raafat, D.; Surmann, K.; Timbermont, L.; Normann, N.; Sellman, B.; van Wamel, W.J.B.; Malhotra-Kumar, S. Exploring Virulence Factors and Alternative Therapies against Staphylococcus aureus Pneumonia. Toxins 2020, 12, 721. [Google Scholar] [CrossRef]
- Pozzi, C.; Olaniyi, R.; Liljeroos, L.; Galgani, I.; Rappuoli, R.; Bagnoli, F. Vaccines for Staphylococcus aureus and Target Populations. Curr. Top. Microbiol. Immunol. 2017, 409, 491–528. [Google Scholar] [CrossRef]
- Murai, M.; Moriyama, H.; Hata, E.; Takeuchi, F.; Amemura-Maekawa, J. Variation and association of fibronectin-binding protein genes fnbA and fnbB in Staphylococcus aureus Japanese isolates. Microbiol. Immunol. 2016, 60, 312–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchiat, C.; Moreau, K.; Devillard, S.; Rasigade, J.P.; Mosnier, A.; Geissmann, T.; Bes, M.; Tristan, A.; Lina, G.; Laurent, F.; et al. Staphylococcus aureus infective endocarditis versus bacteremia strains: Subtle genetic differences at stake. Infect. Genet. Evol. 2015, 36, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Richards, R.L.; Haigh, R.D.; Pascoe, B.; Sheppard, S.K.; Price, F.; Jenkins, D.; Rajakumar, K.; Morrissey, J.A. Persistent Staphylococcus aureus isolates from two independent cases of bacteremia display increased bacterial fitness and novel immune evasion phenotypes. Infect. Immun. 2015, 83, 3311–3324. [Google Scholar] [CrossRef] [Green Version]
- Lilje, B.; Rasmussen, R.V.; Dahl, A.; Stegger, M.; Skov, R.L.; Fowler, V.G.; Ng, K.L.; Kiil, K.; Larsen, A.R.; Petersen, A.; et al. Whole-genome sequencing of bloodstream Staphylococcus aureus isolates does not distinguish bacteraemia from endocarditis. Microb. Genom. 2017, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, D.; Prince, A. Immunopathogenesis of Staphylococcus aureus pulmonary infection. Semin. Immunopathol. 2012, 34, 281–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welte, T.; Pletz, M.W. Antimicrobial treatment of nosocomial meticillin-resistant Staphylococcus aureus (MRSA) pneumonia: Current and future options. Int. J. Antimicrob. Agents 2010, 36, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Rose, H.R.; Holzman, R.S.; Altman, D.R.; Smyth, D.S.; Wasserman, G.A.; Kafer, J.M.; Wible, M.; Mendes, R.E.; Torres, V.J.; Shopsin, B. Cytotoxic Virulence Predicts Mortality in Nosocomial Pneumonia Due to Methicillin-Resistant Staphylococcus aureus. J. Infect. Dis. 2015, 211, 1862–1874. [Google Scholar] [CrossRef] [Green Version]
- Peyrani, P.; Allen, M.; Wiemken, T.L.; Haque, N.Z.; Zervos, M.J.; Ford, K.D.; Scerpella, E.G.; Mangino, J.E.; Kett, D.H.; Ramirez, J.A. Severity of disease and clinical outcomes in patients with hospital-acquired pneumonia due to methicillin-resistant Staphylococcus aureus strains not influenced by the presence of the Panton-Valentine Leukocidin gene. Clin. Infect. Dis. 2011, 53, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Tabor, D.E.; Yu, L.; Mok, H.; Tkaczyk, C.; Sellman, B.R.; Wu, Y.; Oganesyan, V.; Slidel, T.; Jafri, H.; McCarthy, M.; et al. Staphylococcus aureus Alpha-Toxin Is Conserved among Diverse Hospital Respiratory Isolates Collected from a Global Surveillance Study and Is Neutralized by Monoclonal Antibody MEDI4893. Antimicrob. Agents Chemother. 2016, 60, 5312–5321. [Google Scholar] [CrossRef] [Green Version]
- Giulieri, S.G.; Baines, S.L.; Guerillot, R.; Seemann, T.; Goncalves da Silva, A.; Schultz, M.; Massey, R.C.; Holmes, N.E.; Stinear, T.P.; Howden, B.P. Genomic exploration of sequential clinical isolates reveals a distinctive molecular signature of persistent Staphylococcus aureus bacteraemia. Genome Med. 2018, 10, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, B.C.; Golubchik, T.; Batty, E.M.; Fung, R.; Larner-Svensson, H.; Votintseva, A.A.; Miller, R.R.; Godwin, H.; Knox, K.; Everitt, R.G.; et al. Evolutionary dynamics of Staphylococcus aureus during progression from carriage to disease. Proc. Natl. Acad. Sci. USA 2012, 109, 4550–4555. [Google Scholar] [CrossRef] [Green Version]
- Young, B.C.; Wu, C.H.; Gordon, N.C.; Cole, K.; Price, J.R.; Liu, E.; Sheppard, A.E.; Perera, S.; Charlesworth, J.; Golubchik, T.; et al. Severe infections emerge from commensal bacteria by adaptive evolution. elife 2017, 6, e30637. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.O.; Lee, S.; Lee, J.E.; Song, K.H.; Kang, C.K.; Wi, Y.M.; San-Juan, R.; Lopez-Cortes, L.E.; Lacoma, A.; Prat, C.; et al. Dysfunctional accessory gene regulator (agr) as a prognostic factor in invasive Staphylococcus aureus infection: A systematic review and meta-analysis. Sci. Rep. 2020, 10, 20697. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Pollath, C.; Siegmund, A.; Deinhardt-Emmer, S.; Hoerr, V.; Svensson, C.M.; Figge, M.T.; Monecke, S.; Loffler, B. Clinical S. aureus Isolates Vary in Their Virulence to Promote Adaptation to the Host. Toxins 2019, 11, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laabei, M.; Uhlemann, A.C.; Lowy, F.D.; Austin, E.D.; Yokoyama, M.; Ouadi, K.; Feil, E.; Thorpe, H.A.; Williams, B.; Perkins, M.; et al. Evolutionary Trade-Offs Underlie the Multi-faceted Virulence of Staphylococcus aureus. PLoS Biol. 2015, 13, e1002229. [Google Scholar] [CrossRef] [Green Version]
- Berube, B.J.; Sampedro, G.R.; Otto, M.; Wardenburg, J.B. The psm-alpha Locus Regulates Production of Staphylococcus aureus Alpha-Toxin during Infection. Infect. Immun. 2014, 82, 3350–3358. [Google Scholar] [CrossRef] [Green Version]
- Tavares, A.; Nielsen, J.B.; Boye, K.; Rohde, S.; Paulo, A.C.; Westh, H.; Schonning, K.; de Lencastre, H.; Miragaia, M. Insights into alpha-hemolysin (Hla) evolution and expression among Staphylococcus aureus clones with hospital and community origin. PLoS ONE 2014, 9, e98634. [Google Scholar] [CrossRef] [Green Version]
- Lacoma, A.; Mateo, L.; Blanco, I.; Mendez, M.J.; Rodrigo, C.; Latorre, I.; Villar-Hernandez, R.; Dominguez, J.; Prat, C. Impact of Host Genetics and Biological Response Modifiers on Respiratory Tract Infections. Front. Immunol. 2019, 10, 1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, B.; Ballal, A.; Manna, A.C. Transcriptional variation of regulatory and virulence genes due to different media in Staphylococcus aureus. Microb. Pathog. 2009, 47, 94–100. [Google Scholar] [CrossRef]
- Ferrando, C.; Soro, M.; Canet, J.; Unzueta, M.C.; Suarez, F.; Librero, J.; Peiro, S.; Llombart, A.; Delgado, C.; Leon, I.; et al. Rationale and study design for an individualized perioperative open lung ventilatory strategy (iPROVE): Study protocol for a randomized controlled trial. Trials 2015, 16, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarter, Y.; Sharp, S. Best laboratory practices for respiratory cultures. Clin. Microbiol. Newsl. 2013, 35, 35–43. [Google Scholar] [CrossRef]
- Sanchez-Herrero, J.; Lacoma, A.; Molina-Moya, B.; Giménez, M.; Molinos, S.; Prat, C.; Sumoy, L. BacterialTyper: A Bioinformatics Pipeline for the Integrative Analysis of Bacterial Whole Genome Sequencing. Abstract 1981. In Proceedings of the 30th European Congress of Clinical Microbiology and Infectious Diseases, Paris, France, 18–21 April 2020. [Google Scholar]
- Babraham Bioinformatics. FastQC. A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 November 2020).
- Ewels, P.; Magnusson, M.; Lundin, S.; Kaller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Simao, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Ferres, I.; Iraola, G. MLSTar: Automatic multilocus sequence typing of bacterial genomes in R. PeerJ 2018, 6, e5098. [Google Scholar] [CrossRef]
- Sanchez-Herrero, J.; Sullivan, M.J. SpaTyper: Staphylococcal Protein A (spa) Characterization Pipeline (Version v0.3.1). 2020. Available online: https://zenodo.org/record/4063625#.YBqESOhKiUk (accessed on 1 November 2020).
- Clausen, P.; Aarestrup, F.M.; Lund, O. Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinform. 2018, 19, 307. [Google Scholar] [CrossRef]
- Sanchez-Herrero, J. Islandpath. Multi Contig Implementation (Version v1b-0.1). 2020. Available online: https://zenodo.org/record/4063730#.YBqEeehKiUk (accessed on 1 November 2020).
- Akhter, S.; Aziz, R.K.; Edwards, R.A. PhiSpy: A novel algorithm for finding prophages in bacterial genomes that combines similarity- and composition-based strategies. Nucleic Acids Res. 2012, 40, e126. [Google Scholar] [CrossRef]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast genome and metagenome distance estimation using MinHash. Genome Biol. 2016, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.T.; Irber, L. Sourmash: A library for MinHash sketching of DNA. J. Open Source Softw. 2016, 1. [Google Scholar] [CrossRef] [Green Version]
- Hunt, M.; Mather, A.E.; Sanchez-Buso, L.; Page, A.J.; Parkhill, J.; Keane, J.A.; Harris, S.R. ARIBA: Rapid antimicrobial resistance genotyping directly from sequencing reads. Microb. Genom. 2017, 3, e000131. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Wang, L.G.; Lam, T.T.; Xu, S.; Dai, Z.; Zhou, L.; Feng, T.; Guo, P.; Dunn, C.W.; Jones, B.R.; Bradley, T.; et al. Treeio: An R Package for Phylogenetic Tree Input and Output with Richly Annotated and Associated Data. Mol. Biol. Evol. 2020, 37, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2016, 8, 28–36. [Google Scholar] [CrossRef]
- Holden, M.T.; Feil, E.J.; Lindsay, J.A.; Peacock, S.J.; Day, N.P.; Enright, M.C.; Foster, T.J.; Moore, C.E.; Hurst, L.; Atkin, R.; et al. Complete genomes of two clinical Staphylococcus aureus strains: Evidence for the rapid evolution of virulence and drug resistance. Proc. Natl. Acad. Sci. USA 2004, 101, 9786–9791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunter, G.; Goodson, M. Stampy: A statistical algorithm for sensitive and fast mapping of Illumina sequence reads. Genome Res. 2011, 21, 936–939. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, Z.; Caccamo, M.; Turner, I.; Flicek, P.; McVean, G. De novo assembly and genotyping of variants using colored de Bruijn graphs. Nat. Genet. 2012, 44, 226–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Recker, M.; Laabei, M.; Toleman, M.S.; Reuter, S.; Saunderson, R.B.; Blane, B.; Torok, M.E.; Ouadi, K.; Stevens, E.; Yokoyama, M.; et al. Clonal differences in Staphylococcus aureus bacteraemia-associated mortality. Nat. Microbiol. 2017, 2, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Rudkin, J.K.; Laabei, M.; Edwards, A.M.; Joo, H.S.; Otto, M.; Lennon, K.L.; O’Gara, J.P.; Waterfield, N.R.; Massey, R.C. Oxacillin alters the toxin expression profile of community-associated methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 1100–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient ID | Study Group | Number of Isolates | Clinical Samples a | Persistent Isolation b (Days) | Respiratory Complications and/or Mortality Related |
|---|---|---|---|---|---|
| 4 | Pneumonia | 2 | Nasal swab, blood culture | Yes (54) c | Yes |
| 9 | Bronchial colonization | 2 | Tracheal aspirate | No | No |
| 11 | Tracheobronchitis | 2 | Tracheal aspirate, blood culture | No | No |
| 15 | Tracheobronchitis | 3 | Tracheal aspirate | Yes (11) | No |
| 17 | Bronchial colonization | 2 | Tracheal aspirate | Yes (7) | No |
| 18 | Bronchial colonization | 2 | Tracheal aspirate, blood culture | No | No |
| 28 | Tracheobronchitis | 2 | Tracheal aspirate | Yes (8) | No |
| 32 | Bronchial colonization | 3 | Nasal swab, sputum, tracheal aspirate | No | No |
| 36 | Pneumonia | 2 | Tracheal aspirate, blood culture | No | Yes |
| 37 | Bronchial colonization | 4 | Tracheal aspirate (3), nasal swab | Yes (52) | No |
| 41 | Bronchial colonization | 3 | Tracheal aspirate (2), sputum | Yes (57) | No |
| 46 | Tracheobronchitis | 2 | Tracheal aspirate (2) | Yes (8) | No |
| 48 | Bronchial colonization | 2 | Tracheal aspirate (2) | Yes (15) | No |
| 50 | Tracheobronchitis | 2 | Tracheal aspirate (2) | Yes (9) | No |
| 52 | Bronchial colonization | 1 | Tracheal aspirate | No | No |
| 54 | Tracheobronchitis | 2 | Tracheal aspirate (2) | Yes (14) | No |
| 59 | Pneumonia | 3 | Tracheal aspirate (3) | Yes (62) | No |
| 67 | Pneumonia | 2 | Tracheal aspirate, blood culture | No | Yes |
| 83 | Bronchial colonization | 2 | Tracheal aspirate, blood culture | Yes (73) | No |
| 103 | Bronchial colonization | 6 | Sputum (6) | Yes (524) | No |
| 112 | Tracheobronchitis | 2 | Tracheal aspirate, blood culture | No | No |
| 115 | Pneumonia | 7 | Tracheal aspirate (2), blood culture, sputum (2), pleural effusion, protected brush | Yes (74) | Yes |
| 116 | Pneumonia | 2 | Tracheal aspirate | No | Yes |
| 121 | Pneumonia | 3 | Tracheal aspirate | Yes (8) | No |
| 124 | Tracheobronchitis | 2 | Tracheal aspirate | Yes (21) | No |
| 126 | Pneumonia | 4 | Tracheal aspirate | Yes (28) | Yes |
| 133 | Pneumonia | 2 | Tracheal aspirate | Yes (11) | No |
| 144 | Tracheobronchitis | 3 | Tracheal aspirate | Yes (12) | No |
| 146 | Tracheobronchitis | 5 | Tracheal aspirate | Yes (176) | No |
| 181 | Tracheobronchitis | 2 | Tracheal aspirate | Yes (92) | No |
| 184 | Bronchial colonization | 4 | Tracheal aspirate | Yes (15) | No |
| 185 | Bronchial colonization | 4 | Tracheal aspirate (3), nasal swab | Yes (10) | No |
| 191 | Bronchial colonization | 3 | Tracheal aspirate, central venous catheter, blood culture | No | No |
| 193 | Pneumonia | 2 | Tracheal aspirate | No | Yes |
| Patient ID | Number of Isolates | Study Group | Related Isolates Identified | MLST (In Silico) of Earliest Isolate | Number of Within-Host Variants Identified |
|---|---|---|---|---|---|
| 9 | 2 | Bronchial colonization | Yes | 8 | 3 |
| 15 | 3 | Tracheobronchitis | No | 5 | NA a |
| 17 | 2 | Bronchial colonization | Yes | 10 | 3 |
| 28 | 2 | Tracheobronchitis | Yes | 45 | 11 |
| 32 | 2 | Bronchial colonization | Yes | 30 | 31 |
| 41 | 3 | Bronchial colonization | Yes | 15 | 3 |
| 46 | 2 | Tracheobronchitis | Yes | 7 | 1 |
| 48 | 2 | Bronchial colonization | Yes | 121 | 2 |
| 50 | 2 | Tracheobronchitis | Yes | 398 | 1 |
| 54 | 2 | Tracheobronchitis | Yes | 125 | 1 |
| 59 | 3 | Pneumonia | Yes | NF | 0 |
| 103 | 6 | Bronchial colonization | Yes | 125 | 17 |
| 116 | 2 | Pneumonia | Yes | NF | 2 |
| 121 | 3 | Pneumonia | Yes | 22 | 3 |
| 124 | 2 | Tracheobronchitis | Yes | 125 | 0 |
| 126 | 4 | Pneumonia | Yes | NF | 17 |
| 133 | 2 | Pneumonia | Yes | 188 | 0 |
| 144 | 3 | Tracheobronchitis | Yes | 15 | 10 |
| 146 | 5 | Tracheobronchitis | Yes (first 2 of 5) | 12 | 6 |
| 181 | 2 | Tracheobronchitis | No | 7 | NA |
| 184 | 4 | Bronchial colonization | Yes | 97 | 15 |
| 185 | 4 | Bronchial colonization | Yes (first 3 of 4) | 133 | 6 |
| 193 | 2 | Pneumonia | Yes | 5 | 0 |
| 37 | 3 | Bronchial colonization | Yes | 146 | 0 |
| 115 | 6 | Pneumonia | Yes | 146 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lacoma, A.; Laabei, M.; Sánchez-Herrero, J.F.; Young, B.; Godoy-Tena, G.; Gomes-Fernandes, M.; Sumoy, L.; Plans, O.; Arméstar, F.; Prat, C. Genotypic and Phenotypic Characterization of Staphylococcus aureus Isolates from the Respiratory Tract in Mechanically-Ventilated Patients. Toxins 2021, 13, 122. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020122
Lacoma A, Laabei M, Sánchez-Herrero JF, Young B, Godoy-Tena G, Gomes-Fernandes M, Sumoy L, Plans O, Arméstar F, Prat C. Genotypic and Phenotypic Characterization of Staphylococcus aureus Isolates from the Respiratory Tract in Mechanically-Ventilated Patients. Toxins. 2021; 13(2):122. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020122
Chicago/Turabian StyleLacoma, Alicia, Maisem Laabei, Jose Francisco Sánchez-Herrero, Bernadette Young, Gerard Godoy-Tena, Meissiner Gomes-Fernandes, Lauro Sumoy, Oriol Plans, Fernando Arméstar, and Cristina Prat. 2021. "Genotypic and Phenotypic Characterization of Staphylococcus aureus Isolates from the Respiratory Tract in Mechanically-Ventilated Patients" Toxins 13, no. 2: 122. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020122