Electric Blue: Molecular Evolution of Three-Finger Toxins in the Long-Glanded Coral Snake Species Calliophis bivirgatus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
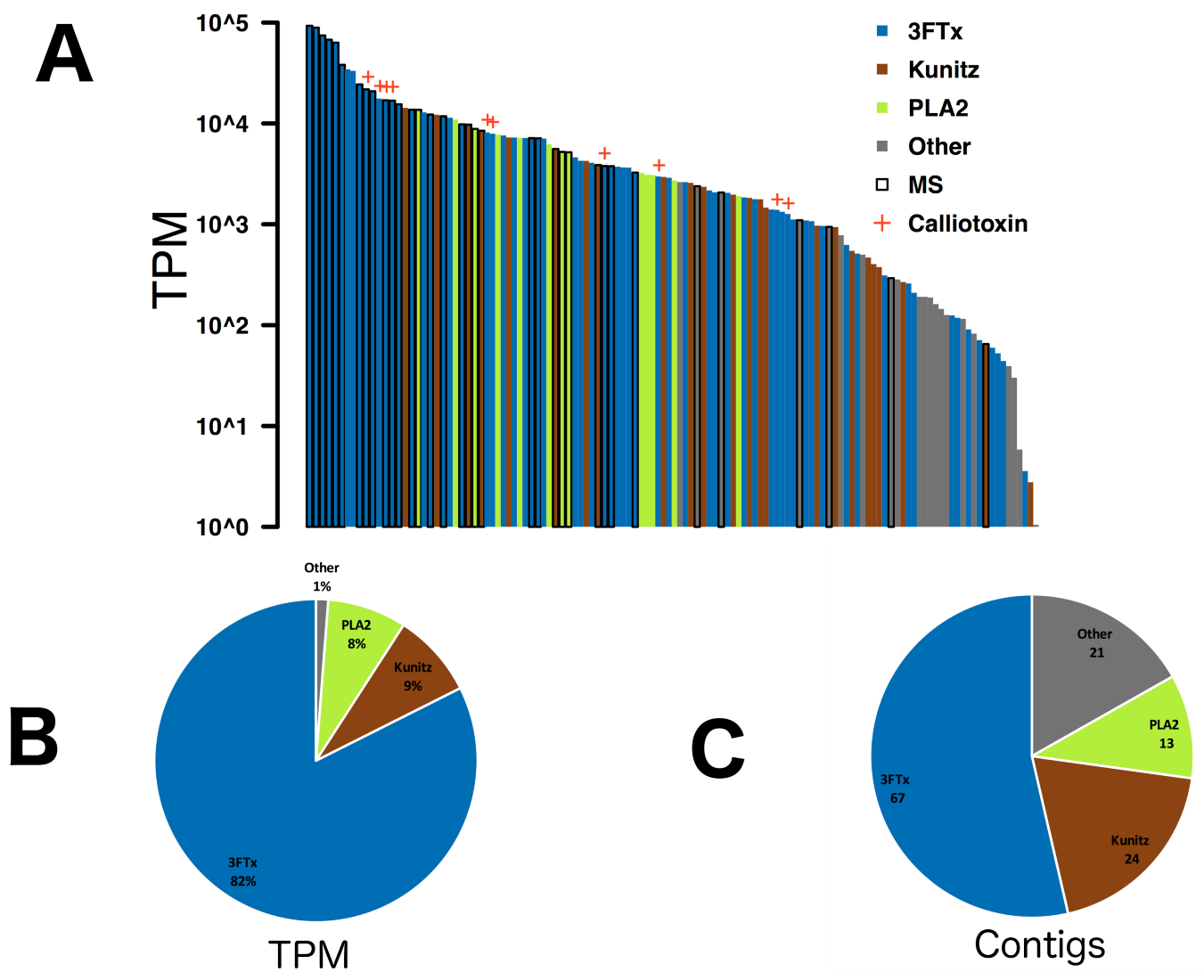
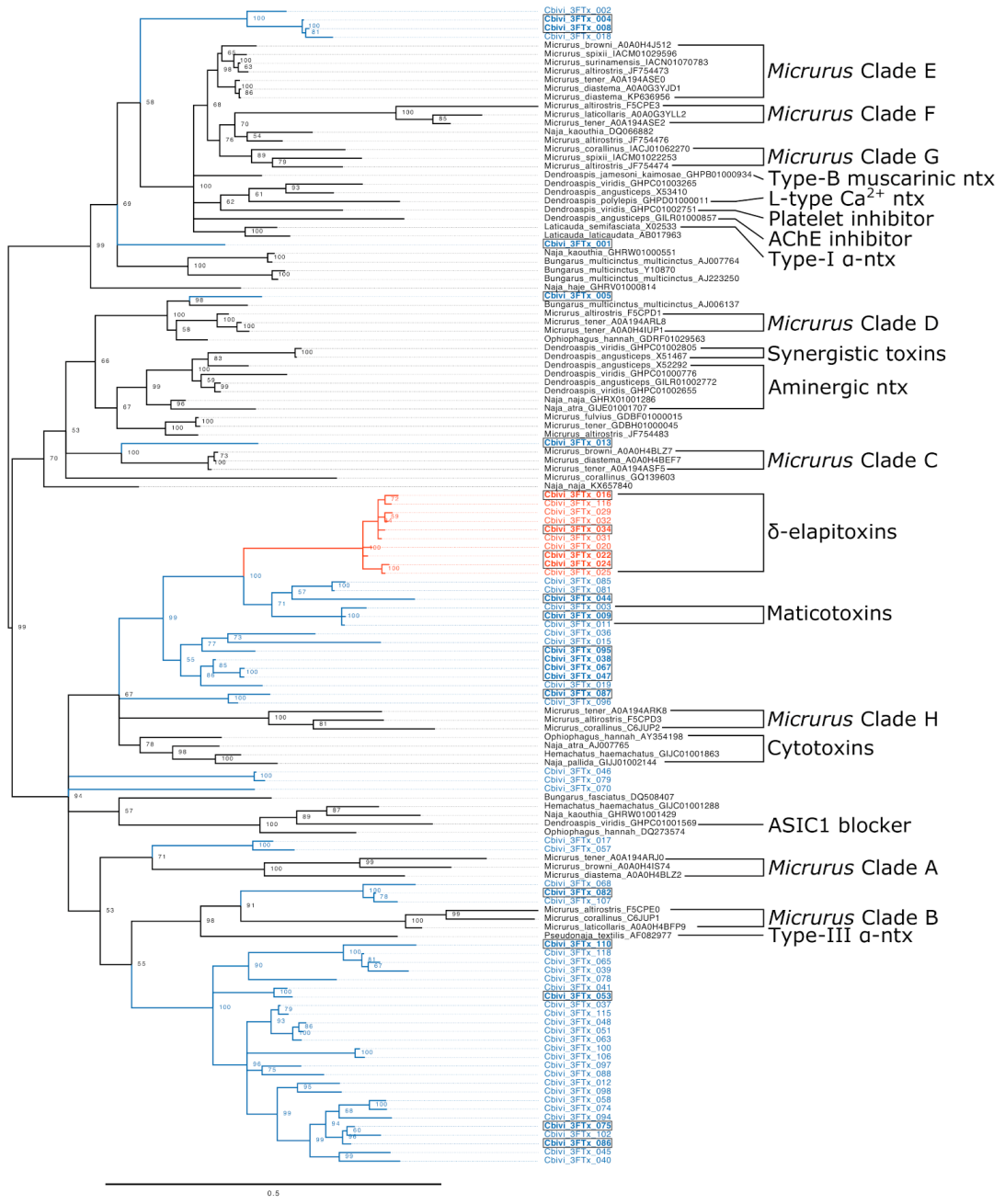
2. Results & Discussion
3. Materials and Methods
3.1. Venom and Tissue Acquisition
3.2. RNA Extraction
3.3. Sequencing
3.4. Assembly
3.5. Annotation
3.6. Tests for Selection
3.7. Phylogenetic Reconstruction
3.8. MS/MS Sample Preparation
3.9. MS/MS
3.10. Visualization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 3FTx | Three-finger toxin |
| NaV | Voltage-gated sodium channel |
| TPM | Transcripts per million |
| CRiSP | Cysteine-rich secretory protein |
| SVMP | Snake venom metalloproteinase |
| MS/MS | Tandem mass spectrometry |
| Ratio of non-synonymous to synonymous substitutions | |
| ORFs | Open reading frames |
References
- Kasturiratne, A.; Wickremasinghe, A.R.; da Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; da Silva, H.J. The Global Burden of Snakebite: A Literature Analysis and Modelling Based on Regional Estimates of Envenoming and Deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warrell, D.A. Snake bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef]
- Uetz, P.; Freed, P.; Hošek, J. (Eds.) The Reptile Database. 2020. Available online: https://www.reptile-database.org (accessed on 12 January 2021).
- Currie, B.J. Snakebite in tropical Australia, Papua New Guinea and Irian Jaya. Emerg. Med. Australas. 2000, 12, 285–294. [Google Scholar] [CrossRef]
- Warrell, D.A. Clinical Toxicology of Snakebite in Africa and the Middle East/Arabian Peninsula. In Handbook of Clinical Toxicology of Animal Venoms and Poisons; CRC Press: Boca Raton, FL, USA, 2017; pp. 433–492. [Google Scholar]
- Warrell, D.A. Clinical Toxicology of Snakebite in Asia. In Handbook of Clinical Toxicology of Animal Venoms and Poisons; CRC Press: Boca Raton, FL, USA, 2017; pp. 493–594. [Google Scholar] [CrossRef]
- Mohapatra, B.; Warrell, D.A.; Suraweera, W.; Bhatia, P.; Dhingra, N.; Jotkar, R.M.; Rodriguez, P.S.; Mishra, K.; Whitaker, R.; Jha, P.; et al. Snakebite Mortality in India: A Nationally Representative Mortality Survey. PLoS Neglected Trop. Dis. 2011, 5, e1018. [Google Scholar] [CrossRef] [PubMed]
- Ariaratnam, C.A.; Sheriff, M.H.R.; Theakston, R.D.G.; Warrell, D.A. Distinctive Epidemiologic and Clinical Features of Common Krait (Bungarus caeruleus) Bites in Sri Lanka. Am. J. Trop. Med. Hyg. 2008, 79, 458–462. [Google Scholar] [CrossRef]
- Warrell, D.A.; Greenwood, B.M.; Davidson, N.M.; Ormerod, L.D.; Prentice, C.R.M. Necrosis, Haemorrhage and Complement Depletion Following Bites by the Spitting Cobra (Naja nigricollis). QJM Int. J. Med. 1976, 45, 1–22. [Google Scholar] [CrossRef]
- Blaylock, R. Epidemiology of snakebite in Eshowe, KwaZulu-Natal, South Africa. Toxicon 2004, 43, 159–166. [Google Scholar] [CrossRef]
- Slowinski, J.B.; Boundy, J.; Lawson, R. The Phylogenetic Relationships of Asian Coral Snakes (Elapidae: Calliophis and Maticora) Based on Morphological and Molecular Characters. Herpetologica 2001, 57, 233–245. [Google Scholar]
- Kelly, C.M.R.; Barker, N.P.; Villet, M.H.; Broadley, D.G. Phylogeny, biogeography and classification of the snake superfamily Elapoidea: A rapid radiation in the late Eocene. Cladistics 2009, 25, 38–63. [Google Scholar] [CrossRef]
- Lee, M.S.Y.; Sanders, K.L.; King, B.; Palci, A. Diversification rates and phenotypic evolution in venomous snakes (Elapidae). Open Sci. 2016, 3, 150277. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G.; Sunagar, K.; Casewell, N.R.; Kochva, E.; Roelants, K.; Scheib, H.; Wüster, W.; Vidal, N.; Young, B.; Burbrink, F.; et al. The origin and evolution of the Toxicofera reptile venom system. In Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Fry, B.G., Ed.; Oxford University Press: Oxford, UK, 2015; pp. 1–31, Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Jackson, K. The evolution of venom-delivery systems in snakes. Zool. J. Linn. Soc. 2003, 137, 337–354. [Google Scholar] [CrossRef] [Green Version]
- Vonk, F.J.; Admiraal, J.F.; Jackson, K.; Reshef, R.; de Bakker, M.A.G.; Vanderschoot, K.; van den Berge, I.; van Atten, M.; Burgerhout, E.; Beck, A.; et al. Evolutionary origin and development of snake fangs. Nature 2008, 454, 630–633. [Google Scholar] [CrossRef]
- Jackson, K. How tubular venom-conducting fangs are formed. J. Morphol. 2002, 252, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Scheib, H.; Weerd, L.v.d.; Young, B.; McNaughtan, J.; Ramjan, S.F.R.; Vidal, N.; Poelmann, R.E.; Norman, J.A. Evolution of an Arsenal Structural and Functional Diversification of the Venom System in the Advanced Snakes (Caenophidia). Mol. Cell. Proteom. 2008, 7, 215–246. [Google Scholar] [CrossRef] [Green Version]
- Kochva, E. On the lateral jaw musculature of the Solenoglypha with remarks on some other snakes. J. Morphol. 1962, 110, 227–284. [Google Scholar] [CrossRef]
- Fry, B.G.; Casewell, N.R.; Wüster, W.; Vidal, N.; Young, B.; Jackson, T.N. The structural and functional diversification of the Toxicofera reptile venom system. Toxicon 2012, 60, 434–448. [Google Scholar] [CrossRef]
- Fry, B.G.; Lumsden, N.G.; Wüster, W.; Wickramaratna, J.C.; Hodgson, W.C.; Kini, R.M. Isolation of a Neurotoxin (α-colubritoxin) from a Nonvenomous Colubrid: Evidence for Early Origin of Venom in Snakes. J. Mol. Evol. 2003, 57, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ramjan, S.F.R.; Kuruppu, S.; Fung, K.; Blair Hedges, S.; Richardson, M.K.; et al. Early evolution of the venom system in lizards and snakes. Nature 2006, 439, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.; Sunagar, K.; Jackson, T.N.; Reeks, T.; Fry, B.G. Three-Finger Toxins (3FTxs). In Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Fry, B.G., Ed.; Oxford University Press: Oxford, UK, 2015; pp. 215–227. [Google Scholar]
- Scherf, T.; Balass, M.; Fuchs, S.; Katchalski-Katzir, E.; Anglister, J. Three-dimensional solution structure of the complex of α-bungarotoxin with a library-derived peptide. Proc. Natl. Acad. Sci. USA 1997, 94, 6059–6064. [Google Scholar] [CrossRef] [Green Version]
- Utkin, Y.N. Last decade update for three-finger toxins: Newly emerging structures and biological activities. World J. Biol. Chem. 2019, 10, 17–27. [Google Scholar] [CrossRef]
- Fry, B.G.; Wüster, W.; Ryan Ramjan, S.F.; Jackson, T.; Martelli, P.; Kini, R.M. Analysis of Colubroidea snake venoms by liquid chromatography with mass spectrometry: Evolutionary and toxinological implications: LC/MS analysis of Colubroidea snake venoms. Rapid Commun. Mass Spectrom. 2003, 17, 2047–2062. [Google Scholar] [CrossRef]
- Fry, B.G.; Wüster, W.; Kini, R.M.; Brusic, V.; Khan, A.; Venkataraman, D.; Rooney, A.P. Molecular Evolution and Phylogeny of Elapid Snake Venom Three-Finger Toxins. J. Mol. Evol. 2003, 57, 110–129. [Google Scholar] [CrossRef] [Green Version]
- Kini, R.M.; Doley, R. Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets. Toxicon 2010, 56, 855–867. [Google Scholar] [CrossRef]
- Utkin, Y.N.; Kukhtina, V.V.; Kryukova, E.V.; Chiodini, F.; Bertrand, D.; Methfessel, C.; Tsetlin, V.I. “Weak toxin” from Naja kaouthia is a nontoxic antagonist of α7 and muscle-type nicotinic acetylcholine receptors. J. Biol. Chem. 2001, 276, 15810–15815. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, J.; Mackessy, S.P.; Fry, B.G.; Bhatia, M.; Mourier, G.; Fruchart-Gaillard, C.; Servent, D.; Ménez, R.; Stura, E.; Ménez, A.; et al. Denmotoxin, a Three-finger Toxin from the Colubrid Snake Boiga dendrophila (Mangrove Catsnake) with Bird-specific Activity. J. Biol. Chem. 2006, 281, 29030–29041. [Google Scholar] [CrossRef] [Green Version]
- Mackessy, S.P.; Sixberry, N.M.; Heyborne, W.H.; Fritts, T. Venom of the Brown Treesnake, Boiga irregularis: Ontogenetic shifts and taxa-specific toxicity. Toxicon 2006, 47, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Heyborne, W.H.; Mackessy, S.P. Identification and characterization of a taxon-specific three-finger toxin from the venom of the Green Vinesnake (Oxybelis fulgidus; family Colubridae). Biochimie 2013, 95, 1923–1932. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, J.; Mackessy, S.P.; Sixberry, N.M.; Stura, E.A.; Le Du, M.H.; Ménez, R.; Foo, C.S.; Ménez, A.; Nirthanan, S.; Kini, R.M. Irditoxin, a novel covalently linked heterodimeric three-finger toxin with high taxon-specific neurotoxicity. FASEB J. 2008, 23, 534–545. [Google Scholar] [CrossRef] [Green Version]
- Dashevsky, D.; Debono, J.; Rokyta, D.; Nouwens, A.; Josh, P.; Fry, B.G. Three-Finger Toxin Diversification in the Venoms of Cat-Eye Snakes (Colubridae: Boiga). J. Mol. Evol. 2018, 86, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Jackson, T.; Undheim, E.; Ali, S.; Antunes, A.; Fry, B. Three-Fingered RAVERs: Rapid Accumulation of Variations in Exposed Residues of Snake Venom Toxins. Toxins 2013, 5, 2172–2208. [Google Scholar] [CrossRef] [Green Version]
- Kessler, P.; Marchot, P.; Silva, M.; Servent, D. The three-finger toxin fold: A multifunctional structural scaffold able to modulate cholinergic functions. J. Neurochem. 2017, 142, 7–18. [Google Scholar] [CrossRef]
- Chang, C.C. Looking Back on the Discovery of α-Bungarotoxin. J. Biomed. Sci. 1999, 6, 368–375. [Google Scholar] [CrossRef]
- Gold, B.S. Neostigmine for the Treatment of Neurotoxicity Following Envenomation by the Asiatic Cobra. Ann. Emerg. Med. 1996, 28, 87–89. [Google Scholar] [CrossRef]
- da Silva, N.J.; Aird, S.D. Prey specificity, comparative lethality and compositional differences of coral snake venoms. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 128, 425–456. [Google Scholar] [CrossRef]
- Cipriani, V.; Debono, J.; Goldenberg, J.; Jackson, T.N.W.; Arbuckle, K.; Dobson, J.; Koludarov, I.; Li, B.; Hay, C.; Dunstan, N.; et al. Correlation between ontogenetic dietary shifts and venom variation in Australian brown snakes (Pseudonaja). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 197, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, S.; Petras, D.; Engmark, M.; Süssmuth, R.D.; Whiteley, G.; Albulescu, L.O.; Kazandjian, T.D.; Wagstaff, S.C.; Rowley, P.; Wüster, W.; et al. The medical threat of mamba envenoming in sub-Saharan Africa revealed by genus-wide analysis of venom composition, toxicity and antivenomics profiling of available antivenoms. J. Proteom. 2018, 172, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Durban, J.; Sasa, M.; Calvete, J.J. Venom gland transcriptomics and microRNA profiling of juvenile and adult yellow-bellied sea snake, Hydrophis platurus, from Playa del Coco (Guanacaste, Costa Rica). Toxicon 2018, 153, 96–105. [Google Scholar] [CrossRef]
- Figueroa, A.; McKelvy, A.D.; Grismer, L.L.; Bell, C.D.; Lailvaux, S.P. A Species-Level Phylogeny of Extant Snakes with Description of a New Colubrid Subfamily and Genus. PLoS ONE 2016, 11, e0161070. [Google Scholar] [CrossRef]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenetics Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.M.; Smart, U.; Leviton, A.E.; Smith, E.N. A New Species of Long-glanded Coralsnake of the Genus Calliophis (Squamata: Elapidae) from Dinagat Island, with Notes on the Biogeography and Species Diversity of Philippine Calliophis and Hemibungarus. Herpetologica 2018, 74, 89–104. [Google Scholar] [CrossRef]
- Brongersma, L.D. Notes on Maticora bivirgata (Boie) and on Bungarus flaviceps Reinh. Zool. Meded. 1948, 30, 1–29. [Google Scholar]
- Das, I.; Ahmed, N.; Liat, L.B. Venomous Terrestrial Snakes of Malaysia: Their Identity and Biology. In Clinical Toxinology in Asia Pacific and Africa; Gopalakrishnakone, P., Faiz, A., Fernando, R., Gnanathasan, C.A., Habib, A.G., Yang, C.C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 53–69. [Google Scholar] [CrossRef]
- Han, K.K. Blue Malayan coral snake preying on a pink-headed reed snake. Singap. Biodivers. Rec. 2020, 220–222. [Google Scholar]
- Weiting, X.; Yea Tian, T. Blue Malayan coral snake biting barred kukri snake. Singap. Biodivers. Rec. 2013, 82–83. [Google Scholar]
- Leviton, A.E. Contributions to a review of Philippine snakes, III. The genera Maticora and Calliophis. Philipp. J. Sci. 1963, 92, 523–550. [Google Scholar]
- Bernhard-Meyer, A. The poison-glands of Callophis intestinalis and C. bivirgatus. Ann. Mag. Nat. Hist. 1869, 4, 74. [Google Scholar] [CrossRef]
- Yang, D.C.; Deuis, J.R.; Dashevsky, D.; Dobson, J.; Jackson, T.N.W.; Brust, A.; Xie, B.; Koludarov, I.; Debono, J.; Hendrikx, I.; et al. The Snake with the Scorpion’s Sting: Novel Three-Finger Toxin Sodium Channel Activators from the Venom of the Long-Glanded Blue Coral Snake (Calliophis bivirgatus). Toxins 2016, 8, 303. [Google Scholar] [CrossRef]
- Fukuyama, I.; Vogel, G.; Matsui, M.; Eto, K.; Munir, M.; Hossman, M.Y.; Hamidy, A.; Nishikawa, K. Systematics of Calliophis intestinalis with the Resurrection of Calliophis nigrotaeniatus (Elapidae, Serpentes). Zool. Sci. 2020, 37, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.D.; Bokhari, S.; Dashevsky, D. Mild Envenomation by Calliophis maculiceps in Northeast Thailand. Herpetol. Rev. 2020, 51, 429–432. [Google Scholar]
- Mirza, Z.; Pal, S. Notes on the effect of a bite from Calliophis melanurus Shaw, 1802 (Serpents: Elapidae: Calliophinae). Reptile Rap 2010, 9, 7–8. [Google Scholar]
- Silva, A.; Jayakody, S.; Ranasinghe, N.; Manawaduge, A.; Perera, K. First Authenticated Case of a Bite by Rare and Elusive Blood-Bellied Coral Snake (Calliophis haematoetron). Wilderness Environ. Med. 2020, 31, 466–469. [Google Scholar] [CrossRef]
- Maduwage, K.; Buddika, H.; Ranasinghe, J.; Wijesooriya, K. Second case report of slender coral snake (Calliophis melanurus sinhaleyus) envenomation of Sri Lanka. Toxicon 2021, 189, 7–9. [Google Scholar] [CrossRef]
- Jacobson, F. A case of snake bite (Maticora intestinalis). Bull. Raffles Mus. 1937, 13, 77. [Google Scholar]
- Meers, P.D. Snake Bite by the Malayan Coral Snake: Maticora Intestinalis. J. R. Army Med. Corps 1968, 114, 152–153. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.T.; Ab Razak, M.N.; Ismail, A.K. Does the legend speaks the truth? Revisiting the venom profile and the effect of Calliophis intestinalis bite to human. Clin. Toxicol. 2020, 58, 351. [Google Scholar]
- Institute of Medical Research. A Death from Snake Bite in Annual Report of the Institute for Medical Research; Institute of Medical Research: Kuala Lumpur, Malaysia, 1956; pp. 71–72. [Google Scholar]
- Harrison, J.L. The bite of a blue Malaysian Coral Snake or Ular Matahari. Malay. Nat. J. 1957, 11, 130–132. [Google Scholar]
- Reid, H.A. Symptomatology, pathology, and treatment of land snake bite in India and Southeast Asia. In Venomous Animals and Their Venoms; Academic Press: Cambridge, MA, USA, 1968; pp. 611–642. [Google Scholar]
- Tan, C.H.; Fung, S.Y.; Yap, M.K.K.; Leong, P.K.; Liew, J.L.; Tan, N.H. Unveiling the elusive and exotic: Venomics of the Malayan blue coral snake (Calliophis bivirgata flaviceps). J. Proteom. 2016, 132, 1–12. [Google Scholar] [CrossRef]
- Tan, C.H.; Liew, J.L.; Tan, K.Y.; Tan, N.H. Genus Calliophis of Asiatic coral snakes: A deficiency of venom cross-reactivity and neutralization against seven regional elapid antivenoms. Toxicon 2016, 121, 130–133. [Google Scholar] [CrossRef]
- Catania, K.C. The Astonishing Behavior of Electric Eels. Front. Integr. Neurosci. 2019, 13, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catania, K.C. Electric Eels Wield a Functional Venom Analogue. Toxins 2021, 1, 48. [Google Scholar] [CrossRef] [PubMed]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef] [Green Version]
- Roze, J.A. Coral Snakes of the Americas: Biology, Identification, and Venoms; Krieger Publishing Company: Malabar, FL, USA, 1996. [Google Scholar]
- Campbell, J.A.; Lamar, W.W. The Venomous Reptiles of the Western Hemisphere; Cornell University Press: Ithaca, NY, USA, 2004. [Google Scholar]
- Wallach, V.; Williams, K.L.; Boundy, J. Snakes of the World: A catalogue of Living and Extinct Species; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Kramer, E. Zur Schlangenfauna Nepals. Rev. Suisse Zool. 1977, 84, 721–761. [Google Scholar] [CrossRef]
- Bucaretchi, F.; Capitani, E.M.D.; Vieira, R.J.; Rodrigues, C.K.; Zannin, M.; Da Silva, N.J., Jr.; Casais-e Silva, L.L.; Hyslop, S. Coral snake bites (Micrurus spp.) in Brazil: A review of literature reports. Clin. Toxicol. 2016, 54, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Cañas, C.A.; Castro-Herrera, F.; Castaño Valencia, S. Envenomation by the red-tailed coral snake (Micrurus mipartitus) in Colombia. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 9. [Google Scholar] [CrossRef] [Green Version]
- Anwar, M.; Bernstein, J.N. North American Coral Snake Envenomation. In Clinical Toxinology in Australia, Europe, and Americas; Vogel, C.W., Seifert, S.A., Tambourgi, D.V., Eds.; Springer: Dordrecht, The Netherlands, 2018; pp. 169–178. [Google Scholar] [CrossRef]
- Takasaki, C.; Yoshida, H.; Shimazu, T.; Teruuchi, T.; Toriba, M.; Tamiya, N. Studies on the venom components of the long-glanded coral snake, Maticora bivirgata. Toxicon 1991, 29, 191–200. [Google Scholar] [CrossRef]
- Fry, B.G.; Richards, R.; Earl, S.; Cousin, X.; Jackson, T.N.; Weise, C.; Sunagar, K. Lesser-known or putative reptile toxins. In Venomous Reptiles & Their Toxins; Oxford University Press: New York, NY, USA, 2015; pp. 364–407. [Google Scholar]
- Dashevsky, D.; Fry, B.G. Ancient Diversification of Three-Finger Toxins in Micrurus Coral Snakes. J. Mol. Evol. 2018, 86, 58–67. [Google Scholar] [CrossRef]
- Margres, M.J.; Aronow, K.; Loyacano, J.; Rokyta, D.R. The venom-gland transcriptome of the eastern coral snake (Micrurus fulvius) reveals high venom complexity in the intragenomic evolution of venoms. BMC Genom. 2013, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.J.; Fry, B.G. Electrostatic resistance to alpha-neurotoxins conferred by charge reversal mutations in nicotinic acetylcholine receptors. Proc. R. Soc. Biol. Sci. 2021, 288, 20202703. [Google Scholar] [CrossRef] [PubMed]
- Takacs, Z.; Wilhelmsen, K.C.; Sorota, S. Snake α-Neurotoxin Binding Site on the Egyptian Cobra (Naja haje) Nicotinic Acetylcholine Receptor Is Conserved. Mol. Biol. Evol. 2001, 18, 1800–1809. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Dashevsky, D.; Kerkkamp, H.; Kordiš, D.; de Bakker, M.A.G.; Wouters, R.; van Thiel, J.; op den Brouw, B.; Vonk, F.; Kini, R.M.; et al. Widespread Evolution of Molecular Resistance to Snake Venom α-Neurotoxins in Vertebrates. Toxins 2020, 12, 638. [Google Scholar] [CrossRef]
- Coimbra, F.C.; Dobson, J.; Zdenek, C.N.; op den Brouw, B.; Hamilton, B.; Debono, J.; Masci, P.; Frank, N.; Ge, L.; Kwok, H.F.; et al. Does size matter? Venom proteomic and functional comparison between night adder species (Viperidae: Causus) with short and long venom glands. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 211, 7–14. [Google Scholar] [CrossRef]
- Mebs, D. Toxicity in animals. Trends in evolution? Toxicon 2001, 39, 87–96. [Google Scholar] [CrossRef]
- Gangur, A.N.; Seymour, J.E.; Liddell, M.J.; Wilson, D.; Smout, M.J.; Northfield, T.D. When is overkill optimal? Tritrophic interactions reveal new insights into venom evolution. Theor. Ecol. 2018, 11, 141–149. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Tange, O. GNU Parallel—The Command-Line Power Tool. Usenix Mag. 2011, 36, 42–47. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Li, H. Toolkit for Processing Sequences in FASTA/Q Formats: lh3/seqtk. 2019. Available online: https://github.com/lh3/seqtk (accessed on 4 June 2019).
- Kircher, M.; Sawyer, S.; Meyer, M. Double indexing overcomes inaccuracies in multiplex sequencing on the Illumina platform. Nucleic Acids Res. 2012, 40, e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marçais, G.; Kingsford, C. A fast, lock-free approach for efficient parallel counting of occurrences of k-mers. Bioinformatics 2011, 27, 764–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapleson, D.; Garcia Accinelli, G.; Kettleborough, G.; Wright, J.; Clavijo, B.J. KAT: A K-mer analysis toolkit to quality control NGS datasets and genome assemblies. Bioinformatics 2017, 33, 574–576. [Google Scholar] [CrossRef]
- Krueger, F. Trim Galore!: A Wrapper Tool around Cutadapt and FastQC to Consistently Apply Quality and Adapter Trimming to FastQ Files. Available online: https://github.com/lh3/seqtk (accessed on 17 June 2019).
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2013, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokyta, D.R.; Lemmon, A.R.; Margres, M.J.; Aronow, K. The venom-gland transcriptome of the eastern diamondback rattlesnake (Crotalus adamanteus). BMC Genom. 2012, 13, 312. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Wu, G.; Tang, J.; Luo, R.; Patterson, J.; Liu, S.; Huang, W.; He, G.; Gu, S.; Li, S. SOAPdenovo-Trans: De novo transcriptome assembly with short RNA-Seq reads. Bioinformatics 2014, 30, 1660–1666. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-Seq: Reference generation and analysis with Trinity. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, T.U. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar] [CrossRef] [Green Version]
- Madden, T.L.; Tatusov, R.L.; Zhang, J. Applications of network BLAST server. Methods Enzymol. 1996, 266, 131–141. [Google Scholar] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows—Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Milne, I.; Stephen, G.; Bayer, M.; Cock, P.J.A.; Pritchard, L.; Cardle, L.; Shaw, P.D.; Marshall, D. Using Tablet for visual exploration of second-generation sequencing data. Briefings Bioinform. 2013, 14, 193–202. [Google Scholar] [CrossRef]
- Wagner, G.P.; Kin, K.; Lynch, V.J. Measurement of mRNA abundance using RNA-seq data: RPKM measure is inconsistent among samples. Theory Biosci. 2012, 131, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Pond, S.L.K.; Frost, S.D.W.; Muse, S.V. HyPhy: Hypothesis testing using phylogenies. Bioinformatics 2005, 21, 676–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. rambaut/figtree. 2020. Available online: https://github.com/rambaut/figtree (accessed on 9 December 2020).
- Mellacheruvu, D.; Wright, Z.; Couzens, A.L.; Lambert, J.P.; St-Denis, N.; Li, T.; Miteva, Y.V.; Hauri, S.; Sardiu, M.E.; Low, T.Y.; et al. The CRAPome: A Contaminant Repository for Affinity Purification Mass Spectrometry Data. Nat. Methods 2013, 10, 730–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- The GIMP Development Team. GIMP. 2019. Available online: https://www.gimp.org (accessed on 12 September 2020).
- Inkscape Project. Inkscape. 2020. Available online: https://inkscape.org (accessed on 12 September 2020).




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dashevsky, D.; Rokyta, D.; Frank, N.; Nouwens, A.; Fry, B.G. Electric Blue: Molecular Evolution of Three-Finger Toxins in the Long-Glanded Coral Snake Species Calliophis bivirgatus. Toxins 2021, 13, 124. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020124
Dashevsky D, Rokyta D, Frank N, Nouwens A, Fry BG. Electric Blue: Molecular Evolution of Three-Finger Toxins in the Long-Glanded Coral Snake Species Calliophis bivirgatus. Toxins. 2021; 13(2):124. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020124
Chicago/Turabian StyleDashevsky, Daniel, Darin Rokyta, Nathaniel Frank, Amanda Nouwens, and Bryan G. Fry. 2021. "Electric Blue: Molecular Evolution of Three-Finger Toxins in the Long-Glanded Coral Snake Species Calliophis bivirgatus" Toxins 13, no. 2: 124. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020124