Transformation of Selected Trichothecenes during the Wheat Malting Production

 , ,
, ,
Abstract
:1. Introduction
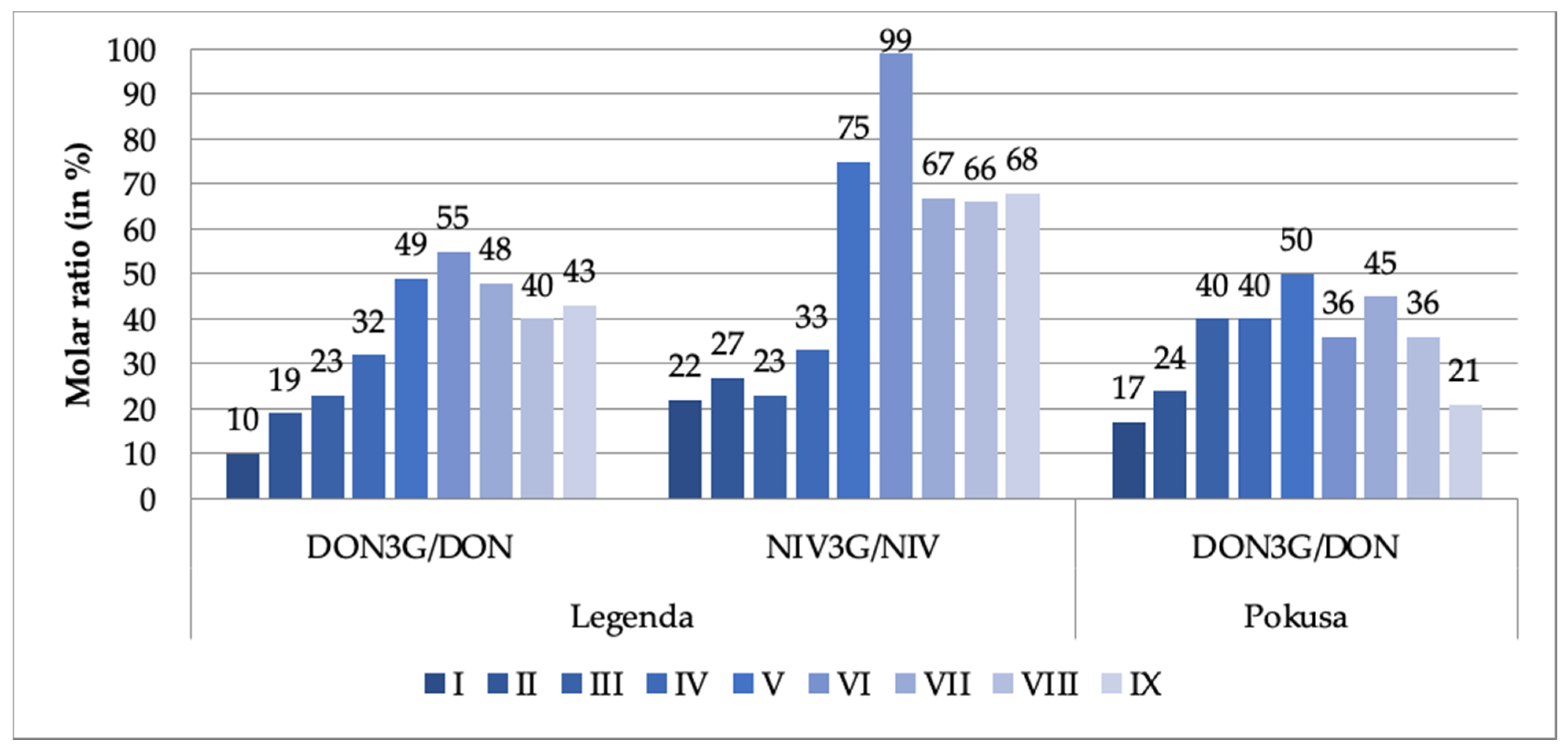
2. Results and Discussion
2.1. Regularly Turned/Aerated Malting Batch
2.2. Unturned Malting Batch
3. Conclusions
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Samples
4.3. Sample Preparation
4.4. UPLC-MS Analysis
4.5. Method Validation
4.6. Statistical Analysis of the Experimental Data
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Krstanovic, V.; Mastanjevic, K.; Velic, N.; Pleadin, J.; Persi, N.; Spanic, V. The influence of Fusarium culmorum contamination level on deoxynivalenol content in wheat, malt and beer. Rom. Biotechnol. Lett. 2015, 20, 10901–10910. [Google Scholar]
- Mastanjevic, K.; Šarkanj, B.; Mastanjevic, K.; Santek, B.; Krstanovic, V. Fusarium culmorum mycotoxin transfer from wheat to malting and brewing products and by-products. World Mycotox. J. 2019, 12, 55–66. [Google Scholar] [CrossRef]
- Spanic, V.; Zdunic, Z.; Drezner, G.; Sarkanj, B. The pressure of Fusarium disease and its relation with mycotoxins in the wheat grain and malt. Toxins 2019, 11, 198. [Google Scholar] [CrossRef] [Green Version]
- Wiśniewska, H.; Stępień, Ł.; Waśkiewicz, A.; Beszterda, M.; Góral, T.; Belter, J. Toxigenic Fusarium species infecting wheat heads in Poland. Cent. Eur. J. Biol. 2014, 9, 163–172. [Google Scholar] [CrossRef]
- Krstanovic, V.; Klapec, T.; Velic, N.; Milakovic, Z. Contamination of malt barley and wheat by Fusarium graminearum and Fusarium culmorum from the crop years 2001–2003 in Eastern Croatia. Microbiol. Res. 2005, 160, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, M.S.; Decundo, J.; Martinez, M.; Dieguez, S.N.; Moreyra, F.; Moreno, M.V.; Stenglein, S.A. Natural contamination with mycotoxins produced by Fusarium graminearum and Fusarium poae in malting barley in Argentina. Toxins 2018, 10, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piacentini, K.C.; Běláková, S.; Benešová, K.; Pernica, M.; Savi, G.D.; Rocha, L.O.; Hartman, I.; Čáslavský, J.; Corrêa, B. Fusarium mycotoxins stability during the malting and brewing processes. Toxins 2019, 11, 257. [Google Scholar] [CrossRef] [Green Version]
- Walder, F.; Schlaeppi, K.; Wittwer, R.; Held, A.Y.; Vogelgsang, S.; Van der Heijden, M.G.A. Community profiling of Fusarium in combination with other plant-associated fungi in different crop species using SMRT sequencing. Front. Plant Sci. 2017, 8, 2019. [Google Scholar] [CrossRef]
- Stenglein, S.A. Fusarium poae: A pathogen that needs more attention. J. Plant. Pathol. 2009, 91, 25–36. [Google Scholar] [CrossRef]
- Schwarz, P.B. Fusarium head blight and deoxynivalenol in malting and brewing: Successes and future challenges. Trop. Plant Pathol. 2017, 42, 153–164. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Cano, P.M.; Seeboth, J.; Meurens, F.; Cognie, J.; Abrami, R.; Oswald, I.P.; Guzylack-Piriou, L. Deoxynivalenol as a new factor in the persistence of intestinal inflammatory diseases: An emerging hypothesis through possible modulation of Th17-mediated response. PLoS ONE 2013, 8, e53647. [Google Scholar] [CrossRef]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated mycotoxin deoxynivalenol. Toxins 2013, 5, 784. [Google Scholar] [CrossRef]
- Pestka, J.J.; Zhou, H.R.; Moon, Y.; Chung, Y.J. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: Unraveling a paradox. Toxicol. Lett. 2004, 153, 61. [Google Scholar] [CrossRef]
- Poapolathep, A.; Kumagai, S.; Suzuki, H.; Doi, K. Development of early apoptosis and changes in T-cell subsets in mouse thymocyte primary cultures treated with nivalenol. Exp. Mol. Pathol. 2004, 77, 149–152. [Google Scholar] [CrossRef]
- Kongkapan, J.; Polapothep, A.; Owen, H.; Giorgi, M. A Brief overview of our current understanding of nivalenol: A growing potential danger yet to be fully investigated. Israel J. Vet. Med. 2016, 71, 3–9. [Google Scholar]
- Palumbo, R.; Crisci, A.; Venâncio, A.; Abrahantes, J.C.; Dorne, J.L.; Battilani, P.; Toscano, P. Occurrence and co-occurrence of mycotoxins in cereal-based feed and food. Microorganisms 2020, 8, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryła, M.; Waśkiewicz, A.; Ksieniewicz-Woźniak, E.; Szymczyk, K.; Jędrzejczak, R. Modified Fusarium mycotoxins in cereals and their products—Metabolism, occurrence, and toxicity: An updated review. Molecules 2018, 23, 963. [Google Scholar] [CrossRef] [Green Version]
- Freire, L.; Sant’Ana, A.S. Modified mycotoxins: An uptake review on their formation, detection, occurrence and toxic effects. Food Chem. Toxicol. 2018, 111, 189–205. [Google Scholar] [CrossRef]
- Zachariasova, M.; Vaclavikova, M.; Lacina, O.; Vaclavik, L.; Hajslova, J. Deoxynivalenol oligoglycosides: New “masked” Fusarium toxins occurring in malt, beer, and breadstuff. J. Agric. Food Chem. 2012, 60, 9280–9291. [Google Scholar] [CrossRef] [PubMed]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Yoshinari, T.; Waśkiewicz, A.; Szymczyk, K. Contamination of wheat cultivated in various regions of Poland during 2017 and 2018 agricultural seasons with selected trichothecenes and their modified forms. Toxins 2019, 11, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; et al. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, 4718. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; et al. Appropriateness to set a group health based guidance value for nivalenol and its modified forms. EFSA J. 2017, 15, 4751. [Google Scholar] [CrossRef] [Green Version]
- Lemmens, M.; Buerstmayr, H.; Krska, R.; Schuhmacher, R.; Grausgruber, H.; Ruckenbauer, P. The effect of inoculation treatment and long-term application of moisture on Fusarium head blight symptoms and deoxynivalenol contamination in wheat grains. Eur. J. Plant Pathol. 2004, 110, 299–308. [Google Scholar] [CrossRef]
- Cowger, C.; Arellano, C. Fusarium graminearum infection and deoxynivalenol concentrations during development of wheat spikes. Phytopathology 2013, 103, 460–471. [Google Scholar] [CrossRef] [Green Version]
- Bryła, M.; Waśkiewicz, A.; Podolska, G.; Szymczyk, K.; Jędrzejczak, R.; Damaziak, K.; Sułek, A. Occurrence of 26 mycotoxins in the grain of cereals cultivated in Poland. Toxins 2016, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Van Nierop, S.; Rautenbach, M.; Axcell, B.; Cantrell, I. The impact of microorganisms on barley and malt—A review. J. Am Soc. Brew. Chem. 2006, 64, 69–78. [Google Scholar] [CrossRef]
- Habler, K.; Hofer, K.; Geissinger, C.; Schuler, J.; Huckelhoven, R.; Hess, M.; Gastl, M.; Rychlik, M. Fate of Fusarium toxins during the malting process. J Agric. Food Chem. 2016, 64, 1377–1384. [Google Scholar] [CrossRef]
- Pascari, X.; Gil-Samarra, S.; Marín, S.; Ramos, A.J.; Sanchis, V. Fate of zearalenone, deoxynivalenol and deoxynivalenol-3-glucoside during malting process. LWT Food Sci. Tech. 2019, 99, 540–546. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, P.B.; Casper, H.H.; Beattie, S. Fate and development of naturally occurring Fusarium mycotoxins during malting and brewing. J. Am. Soc. Brew. Chem. 1995, 53, 121–127. [Google Scholar]
- Huff, W.E.; Hagler, W.M. Density segregation of corn and wheat naturally contaminated with aflatoxin, deoxynivalenol and zearalenone. J. Food Prot. 1985, 48, 416–420. [Google Scholar] [CrossRef]
- Berthiller, F.; Dall’Asta, C.; Corradini, R.; Marchelli, R.; Sulyok, M.; Krska, R.; Adam, G.; Schuhmacher, R. Occurrence of deoxynivalenol and its 3-β-D-glucoside in wheat and maize. Food Addit. Contam. A 2009, 26, 507–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall’Asta, C.; Dall’Erta, A.; Mantovani, P.; Massi, A.; Galaverna, G. Occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in durum wheat. World Mycotox. J. 2013, 6, 83–91. [Google Scholar] [CrossRef]
- Ksieniewicz-Woźniak, E.; Bryła, M.; Waśkiewicz, A.; Yoshinari, T.; Szymczyk, K. Selected trichothecenes in barley malt and beer from Poland and an assessment of dietary risks associated with their consumption. Toxins 2019, 11, 715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habschied, K.; Krska, R.; Sulyok, M.; Lukinac, J.; Jukić, M.; Šarkanj, B.; Krstanović, V.; Mastanjević, K. The influence of steeping water change during malting on the multi-toxin content in malt. Foods 2019, 8, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancova, K.; Hajslova, J.; Poustka, J.; Krplova, A.; Zachariasova, M.; Dostalek, P.; Sachambula, L. Transfer of Fusarium mycotoxins and ‘masked’ deoxynivalenol (deoxynivalenol-3-glucoside) from field barley through malt to beer. Food Addit. Contam. A 2008, 25, 732–744. [Google Scholar] [CrossRef] [Green Version]
- Yoshinari, T.; Sakuda, S.; Furihata, K.; Furusawa, H.; Ohnishi, T.; Sugita-Konishi, Y.; Ishizaki, N.; Terajima, J. Structural determination of a nivalenol glucoside and development of an analytical method for the simultaneous determination of nivalenol and deoxynivalenol, and their glucosides, in wheat. J. Agric. Food Chem. 2014, 62, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EC) No 401/2006 of 23 February 2006 laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Off. J. Eur. Union 2006, L70, 12–34. [Google Scholar]



{kind=link}
{kind=link}
| Wheat Variety | Mycotoxin | Concentration ± SD (µg/kg) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | ||
| Dry Grain | Grain after Steeping | Day 2 | Day 3 | Day 4 | Day 5 | Day 6 | Day 7 | Day 7 (Unturned Batch) | ||
| Legenda | DON | a 1540 ± 332 | a 986 ± 103 | a 1178 ± 98 | a 939 ± 105 | a 1190 ± 238 | a 1349 ± 122 | b 2349 ± 267 | b 3010 ± 338 | c 7077 ± 1006 |
| DON3G | a 236 ± 12 | a 285 ± 15 | a 428 ± 46 | b 467 ± 17 | c 907 ± 222 | c 1146 ± 191 | d 1744 ± 191 | d 1850 ± 397 | e 4751 ± 569 | |
| NIV | d 702 ± 64 | c 519 ± 6 | c 453 ± 10 | ab 328 ± 59 | ab 314 ± 34 | a 270 ± 24 | bc 411 ± 39 | d 691 ± 65 | e 841 ± 62 | |
| NIV3G | b 239 ± 45 | ab 220 ± 27 | a 163 ± 15 | ab 169 ± 11 | c 365 ± 90 | c 413 ± 27 | c 426 ± 44 | d 705 ± 13 | e 881 ± 54 | |
| Pokusa | DON | bc 2164 ± 210 | ab 1642 ± 48 | a 1239 ± 178 | a 1145 ± 106 | ab 1216 ± 119 | c 2770 ± 375 | c 2953 ± 135 | d 4678 ± 963 | e 20,444 ± 873 |
| DON3G | a 584 ± 143 | a 620 ± 87 | a 773 ± 113 | a 710 ± 97 | a 938 ± 101 | b 1533 ± 229 | c 2050 ± 144 | d 2609 ± 477 | e 6672 ± 667 | |
| Wheat Variety | Mycotoxin | II/I | III/II | IV/III | V/IV | VI/V | VII/VI | VIII/VII | VIII/II | IX/II | VIII/I | IX/I | IX/VIII |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Legenda | DON | −36 | +19 | –20 | +27 | +13 | +74 * | +28 | +205 * | +617 * | +95 * | +360 * | +135 * |
| DON3G | +21 | +50 | +9 | +94 * | +26 | +52 * | +6 | +548 * | +1565 * | +683 * | +1913 * | +157 * | |
| NIV | –26 * | –13 | –28 * | –4 | –14 | +52 * | +68 * | +33 * | +62 * | −2 | +20 * | +22 * | |
| NIV3G | –8 | –26 | +3 | +116 * | +13 | +3 | +65 * | +220 * | +300 * | +196 * | +270 * | +25 * | |
| Pokusa | DON | –24 | –25 | –8 | +6 | +128 * | +7 | +58 * | +185 * | +1145 * | +116 * | +845 * | +337 * |
| DON3G | +6 | +25 | –8 | +32 | +63 * | +34 | +27 | +321 * | +976 * | +347 * | +1042 * | +156 * |
| Analyte | Ion Mass (m/z) | Retention Time (min) | LOD (μg/kg) | LOQ (μg/kg) |
|---|---|---|---|---|
| DON | 341.2 (M+FA-H)− | 4.19 | 167 | 557 |
| DON3G | 503.2 (M+FA-H)− | 4.33 | 83 | 277 |
| NIV | 357.2 (M+FA-H)− | 2.30 | 83 | 277 |
| NIV3G | 519.2 (M+FA-H)− | 2.42 | 33 | 110 |
| Recovery Rate (R) and Relative Standard Deviation (RSD) | DON | DON3G | NIV | NIV3G |
|---|---|---|---|---|
| fortification level (n = 4) | 1200 | 400 | 125 | 50 |
| R (%) | 97.8 | 82.3 | 90.6 | 105.8 |
| RSD (%) | 18.3 | 10.1 | 14.0 | 13.4 |
| fortification level (n = 4) | 5000 | 1500 | 500 | 198 |
| R (%) | 92.2 | 86.0 | 92.3 | 88.0 |
| RSD (%) | 13.1 | 10.1 | 9.2 | 11.3 |
| fortification level (n = 4) | 10,000 | 3000 | 1000 | 396 |
| R (%) | 103.3 | 98.3 | 97.1 | 95.4 |
| RSD (%) | 5.4 | 6.6 | 9.4 | 11.3 |
| fortification level (n = 4) | 20,000 | 6000 | 2000 | 792 |
| R (%) | 91.1 | 84.4 | 93.3 | 86.3 |
| RSD (%) | 4.1 | 9.8 | 9.2 | 11.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryła, M.; Ksieniewicz-Woźniak, E.; Michałowska, D.; Waśkiewicz, A.; Yoshinari, T.; Gwiazdowski, R. Transformation of Selected Trichothecenes during the Wheat Malting Production. Toxins 2021, 13, 135. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020135
Bryła M, Ksieniewicz-Woźniak E, Michałowska D, Waśkiewicz A, Yoshinari T, Gwiazdowski R. Transformation of Selected Trichothecenes during the Wheat Malting Production. Toxins. 2021; 13(2):135. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020135
Chicago/Turabian StyleBryła, Marcin, Edyta Ksieniewicz-Woźniak, Dorota Michałowska, Agnieszka Waśkiewicz, Tomoya Yoshinari, and Romuald Gwiazdowski. 2021. "Transformation of Selected Trichothecenes during the Wheat Malting Production" Toxins 13, no. 2: 135. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020135