In Vitro and In Vivo Antibacterial Activity of Gliotoxin Alone and in Combination with Antibiotics against Staphylococcus aureus

 , ,
, ,
Abstract
:1. Introduction
2. Results
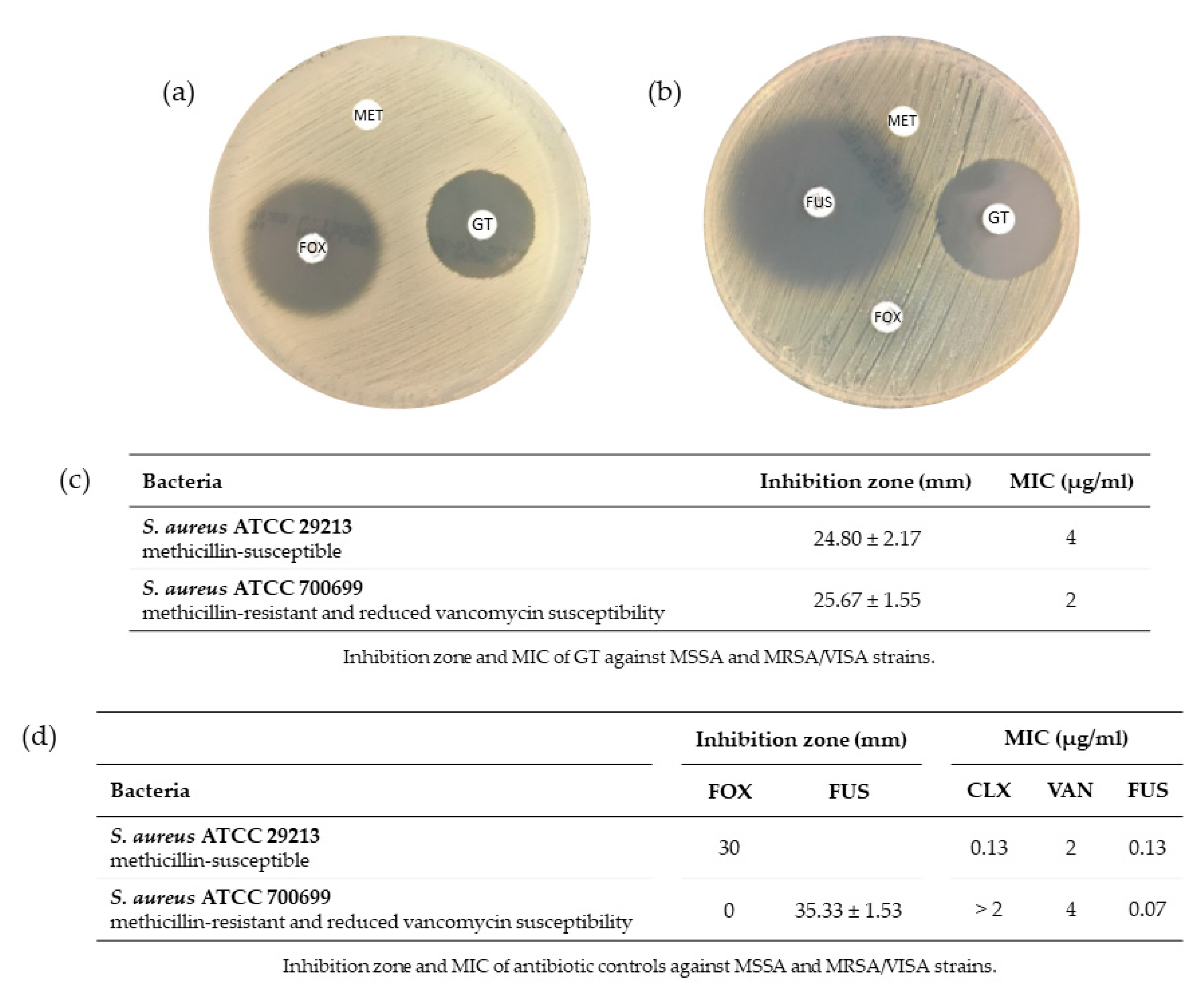
2.1. Antimicrobial Susceptibility of S. aureus against GT
2.1.1. Effect of GT on S. aureus Growth
2.1.2. Minimum Inhibitory Concentration of GT and Antibiotics against S. aureus
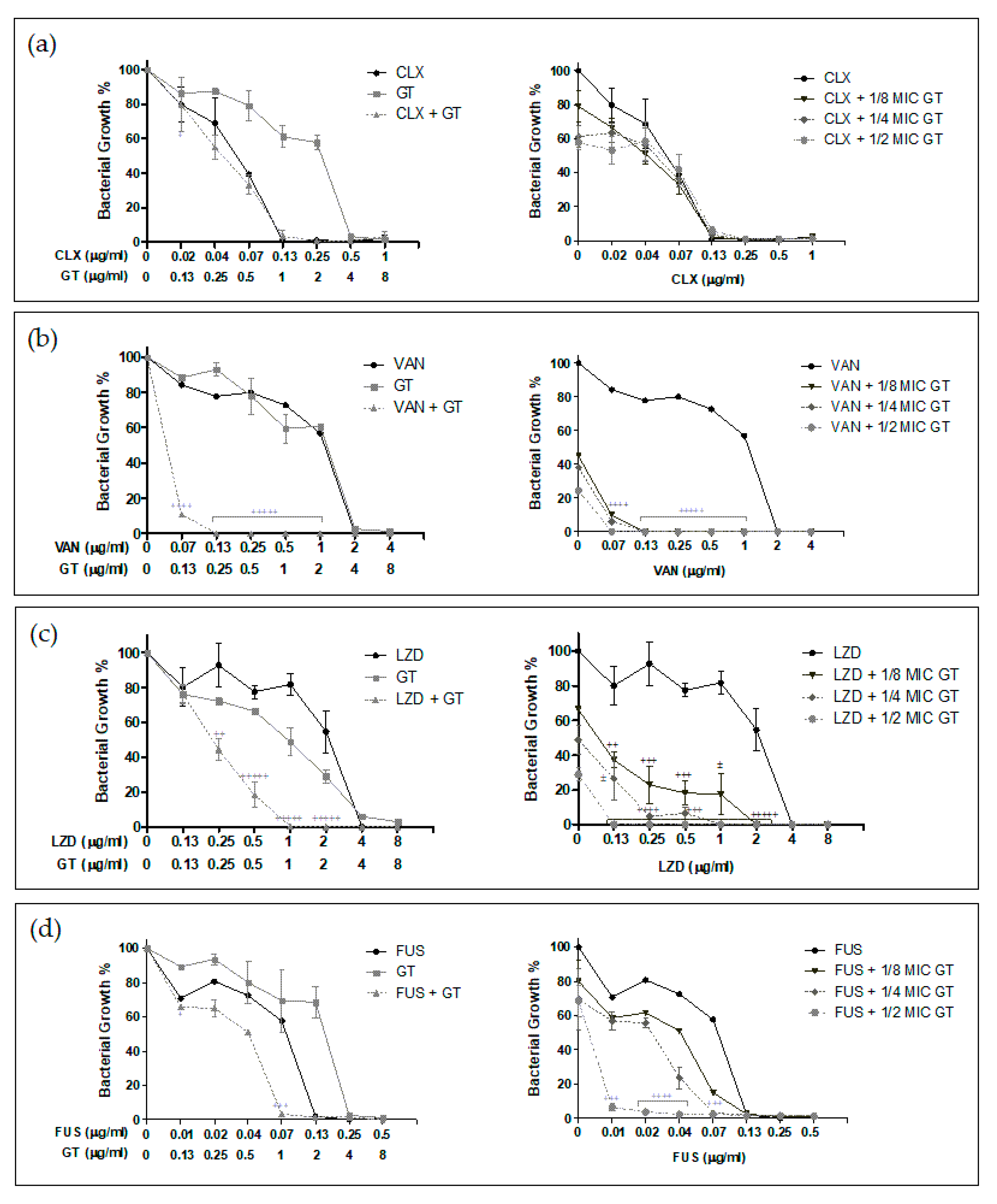
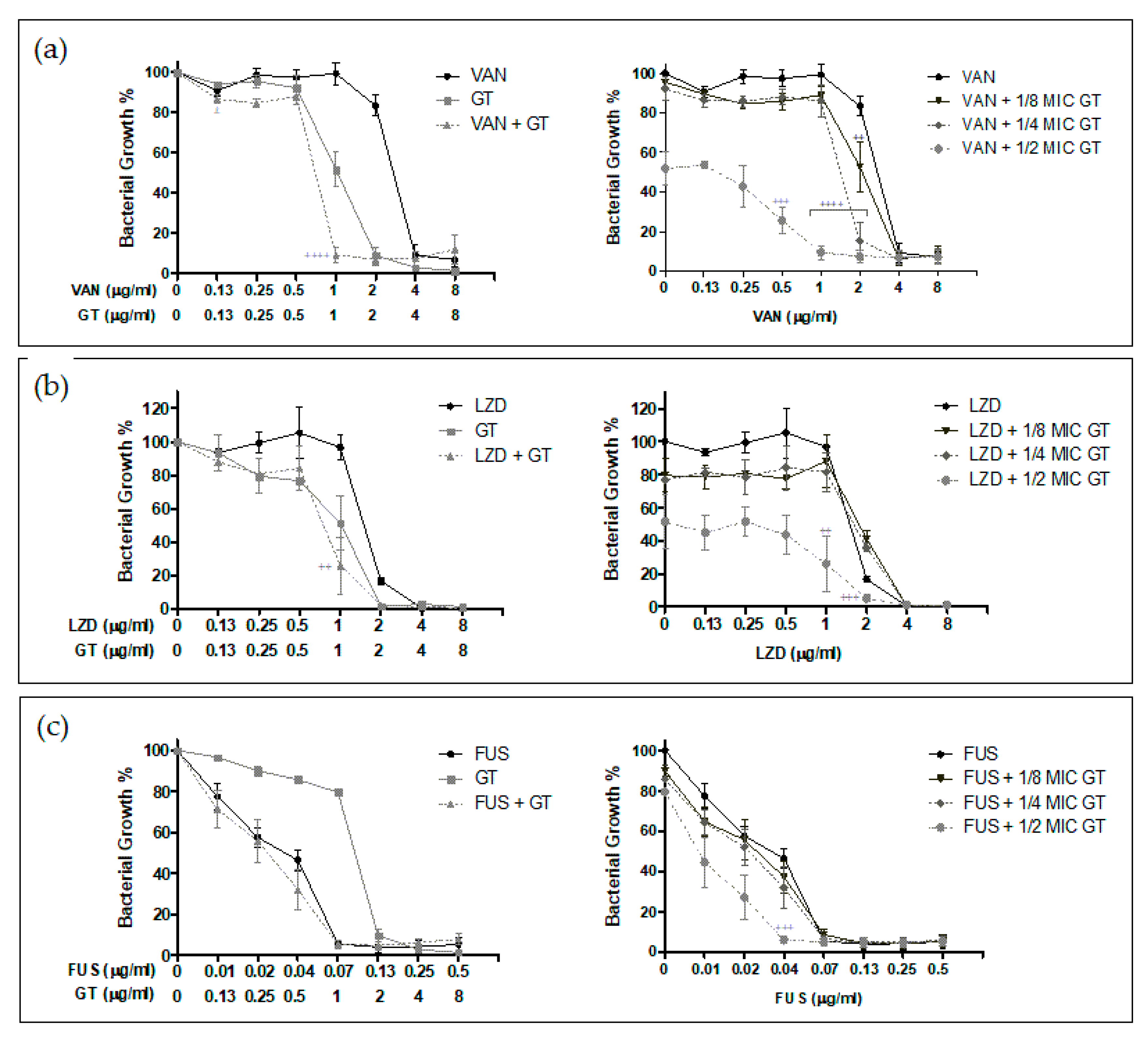
2.2. Antimicrobial Synergy of GT in Combination with Anti-Staphylococcal Drugs
2.3. Mutant Prevention Concentration of GT and Vancomycin Alone or in Combination against S. aureus
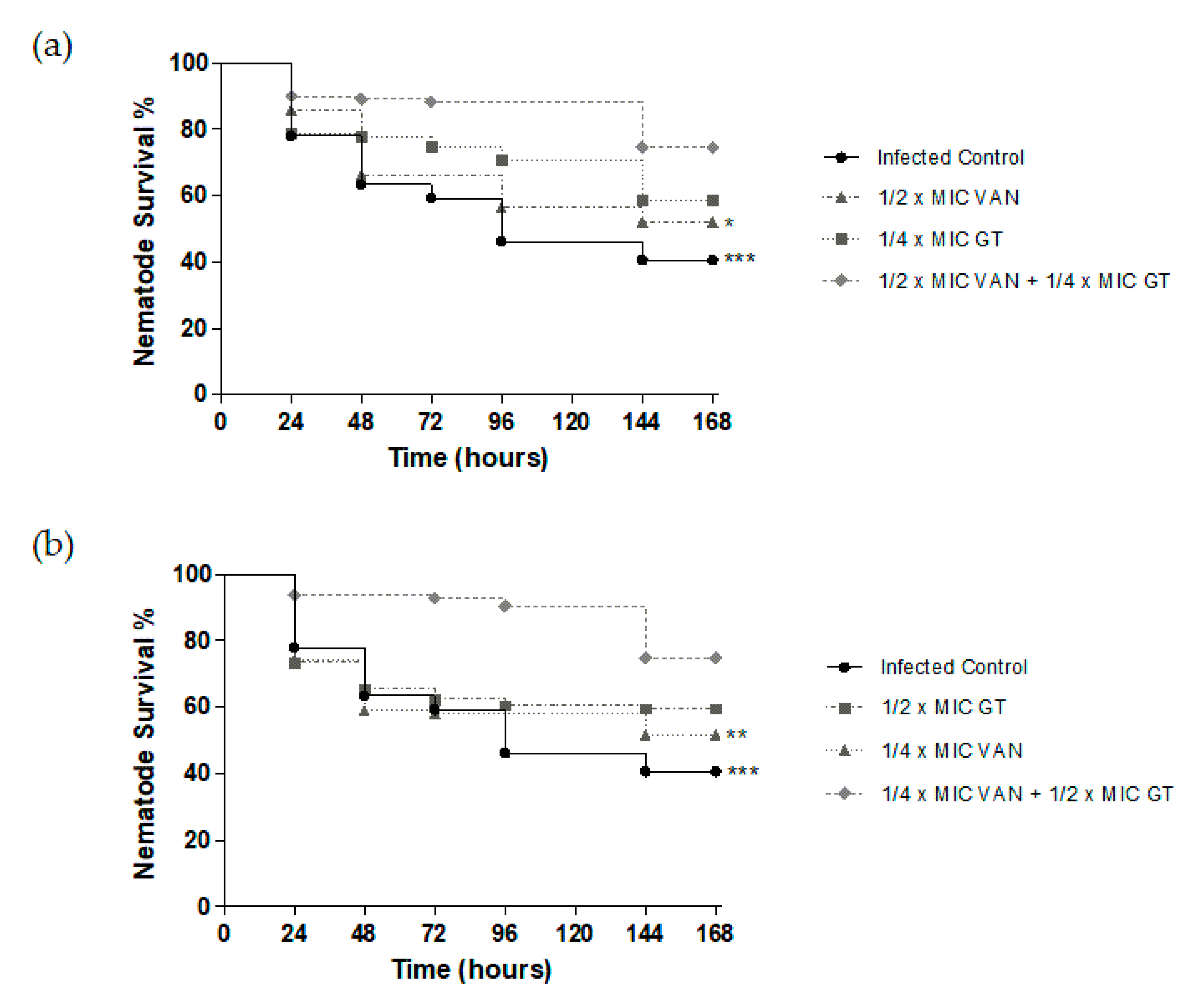
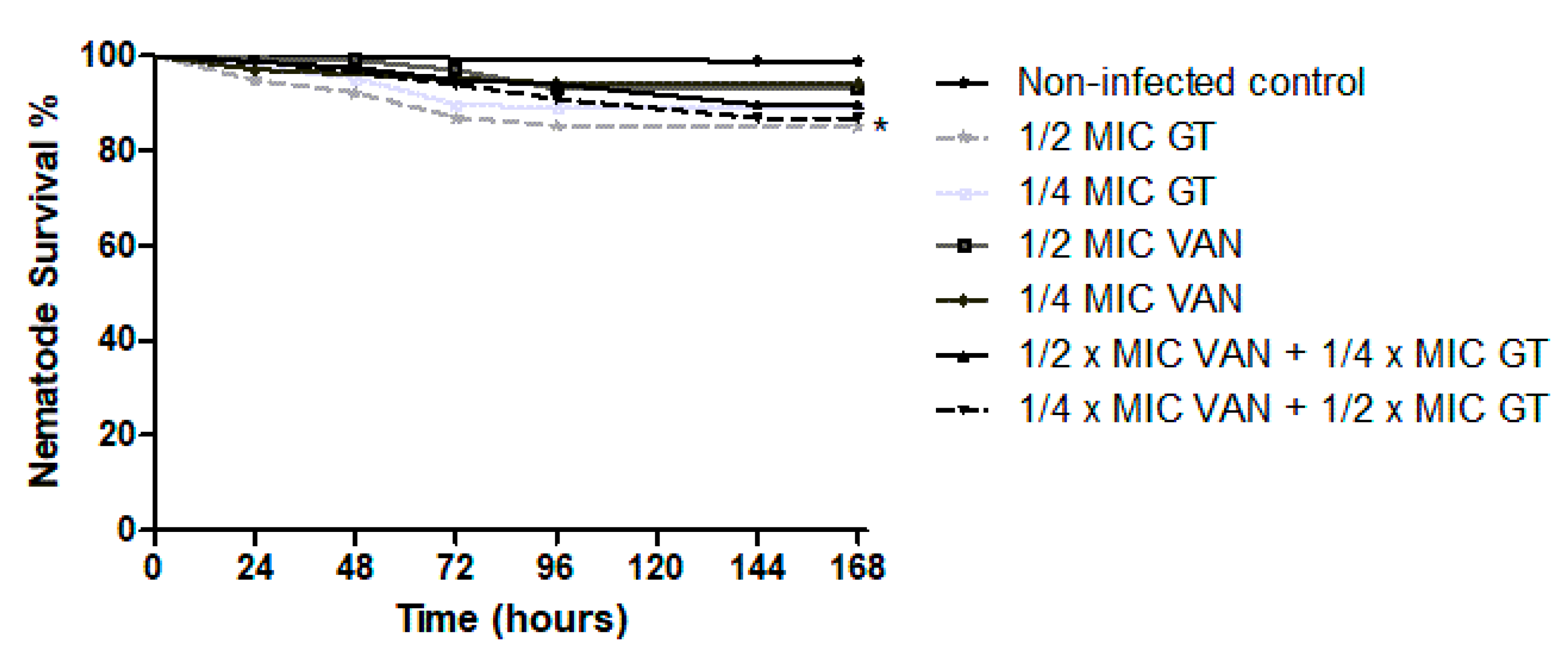
2.4. In Vivo Antibiotic Efficacy of GT as Monotherapy and in Combination with Vancomycin against S. aureus in C. elegans Model
3. Discussion
4. Materials and Methods
4.1. Staphylococcus aureus Strains
4.2. Antimicrobial Susceptibility Assays
4.3. Antimicrobial Synergy Study—Checkerboard Testing
4.4. Determination of the Mutant Prevention Concentration
4.5. C. elegans Culturing and Synchronization
4.6. C. elegans L4 Larvae Culture and Infection
4.7. C. elegans Survival Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gorwitz, R.J.; Kruszon-Moran, D.; McAllister, S.K.; McQuillan, G.; McDougal, L.K.; Fosheim, G.E.; Jensen, B.J.; Killgore, G.; Tenover, F.C.; Kuehnert, M.J. Changes in the prevalence of nasal colonization with Staphylococcus aureus in the United States, 2001–2004. J. Infect. Dis. 2008, 197, 1226–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, D.; Chamberlaine, A. Staphylococcus aureus: A review of the literature. J. Clin. Nurs. 1995, 4, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Mainous, A.G., 3rd; Hueston, W.J.; Everett, C.J.; Diaz, V.A. Nasal carriage of Staphylococcus aureus and methicillin-resistant S aureus in the United States, 2001–2002. Ann. Fam. Med. 2006, 4, 132–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, D.H.; Howden, B.P. Diagnosis and management of Staphylococcus aureus bacteraemia. Intern. Med. J. 2005, 35, S17–S24. [Google Scholar] [CrossRef]
- Murdoch, D.R.; Corey, G.R.; Hoen, B.; Miró, J.M.; Fowler, V.G., Jr.; Bayer, A.S.; Karchmer, A.W.; Olaison, L.; Pappas, P.A.; Moreillon, P.; et al. Clinical presentation, etiology, and outcome of infective endocarditis in the 21st century: The International Collaboration on Endocarditis-Prospective Cohort Study. Arch. Intern. Med. 2009, 169, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.; Borges, A.; Simões, M. Staphylococcus aureus Toxins and Their Molecular Activity in Infectious Diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.; Chambers, S. Diagnosis and management of Staphylococcus aureus infections of the skin and soft tissue. Intern. Med. J. 2005, 35, S97–S105. [Google Scholar] [CrossRef]
- Eriksen, K.R. [“Celbenin”-resistant staphylococci]. Ugeskr. Laeger 1961, 123, 384–386. [Google Scholar]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 2018, 4, 18033. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybak, M.J.; Le, J.; Lodise, T.P.; Levine, D.P.; Bradley, J.S.; Liu, C.; Mueller, B.A.; Pai, M.P.; Wong-Beringer, A.; Rotschafer, J.C.; et al. Therapeutic monitoring of vancomycin for serious methicillin-resistant Staphylococcus aureus infections: A revised consensus guideline and review by the American Society of Health-System Pharmacists, the Infectious Diseases Society of America, the Pediatric Infectious Diseases Society, and the Society of Infectious Diseases Pharmacists. Am. J. Health Syst. Pharm. 2020, 77, 835–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar] [PubMed]
- Hidayat, L.K.; Hsu, D.I.; Quist, R.; Shriner, K.A.; Wong-Beringer, A. High-dose vancomycin therapy for methicillin-resistant Staphylococcus aureus infections: Efficacy and toxicity. Arch. Intern. Med. 2006, 166, 2138–2144. [Google Scholar] [CrossRef] [Green Version]
- Hiramatsu, K.; Aritaka, N.; Hanaki, H.; Kawasaki, S.; Hosoda, Y.; Hori, S.; Fukuchi, Y.; Kobayashi, I. Dissemination in Japanese hospitals of strains of Staphylococcus aureus heterogeneously resistant to vancomycin. Lancet 1997, 350, 1670–1673. [Google Scholar] [CrossRef]
- Howe, R.A.; Bowker, K.E.; Walsh, T.R.; Feest, T.G.; MacGowan, A.P. Vancomycin-resistant Staphylococcus aureus. Lancet 1998, 351, 602. [Google Scholar] [CrossRef] [Green Version]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, D.M.; Waring, P.; Howlett, B.J. The epipolythiodioxopiperazine (ETP) class of fungal toxins: Distribution, mode of action, functions and biosynthesis. Microbiology 2005, 151, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Waring, P.; Beaver, J. Gliotoxin and related epipolythiodioxopiperazines. Gen. Pharmacol. 1996, 27, 1311–1316. [Google Scholar] [CrossRef]
- Davis, C.; Carberry, S.; Schrettl, M.; Singh, I.; Stephens, J.C.; Barry, S.M.; Kavanagh, K.; Challis, G.L.; Brougham, D.; Doyle, S. The role of glutathione S-transferase GliG in gliotoxin biosynthesis in Aspergillus fumigatus. Chem. Biol. 2011, 18, 542–552. [Google Scholar] [CrossRef] [Green Version]
- Müllbacher, A.; Moreland, A.F.; Waring, P.; Sjaarda, A.; Eichner, R.D. Prevention of graft-versus-host disease by treatment of bone marrow with gliotoxin in fully allogeneic chimeras and their cytotoxic T cell repertoire. Transplantation 1988, 46, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Braithwaite, A.W.; Eichner, R.D.; Waring, P.; Müllbacher, A. The immunomodulating agent gliotoxin causes genomic DNA fragmentation. Mol. Immunol. 1987, 24, 47–55. [Google Scholar] [CrossRef]
- Golden, M.C.; Hahm, S.J.; Elessar, R.E.; Saksonov, S.; Steinberg, J.J. DNA damage by gliotoxin from Aspergillus fumigatus. An occupational and environmental propagule: Adduct detection as measured by 32P DNA radiolabelling and two-dimensional thin-layer chromatography. Mycoses 1998, 41, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, S.M.; Mäki-Paakkanen, J.; Hirvonen, M.R.; Roponen, M.; von Wright, A. Genotoxicity of gliotoxin, a secondary metabolite of Aspergillus fumigatus, in a battery of short-term test systems. Mutat. Res. 2002, 520, 161–170. [Google Scholar] [CrossRef]
- Liang, W.L.; Le, X.; Li, H.J.; Yang, X.L.; Chen, J.X.; Xu, J.; Liu, H.L.; Wang, L.Y.; Wang, K.T.; Hu, K.C.; et al. Exploring the chemodiversity and biological activities of the secondary metabolites from the marine fungus Neosartorya pseudofischeri. Mar. Drugs 2014, 12, 5657–5676. [Google Scholar] [CrossRef] [Green Version]
- Svahn, K.S.; Göransson, U.; El-Seedi, H.; Bohlin, L.; Larsson, D.G.; Olsen, B.; Chryssanthou, E. Antimicrobial activity of filamentous fungi isolated from highly antibiotic-contaminated river sediment. Infect. Ecol. Epidemiol. 2012, 2. [Google Scholar] [CrossRef]
- Sahoo, K.C.; Sahoo, S.; Marrone, G.; Pathak, A.; Lundborg, C.S.; Tamhankar, A.J. Climatic factors and community—Associated methicillin-resistant Staphylococcus aureus skin and soft-tissue infections—A time-series analysis study. Int. J. Environ. Res. Public Health 2014, 11, 8996–9007. [Google Scholar] [CrossRef] [Green Version]
- Reece, E.; Doyle, S.; Greally, P.; Renwick, J.; McClean, S. Aspergillus fumigatus Inhibits Pseudomonas aeruginosa in Co-culture: Implications of a Mutually Antagonistic Relationship on Virulence and Inflammation in the CF Airway. Front. Microbiol. 2018, 9, 1205. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.J.; Ghosh, S.; Okoli, I.; Mylonakis, E. Antifungal activity of microbial secondary metabolites. PLoS ONE 2011, 6, e25321. [Google Scholar] [CrossRef] [Green Version]
- Carberry, S.; Molloy, E.; Hammel, S.; O’Keeffe, G.; Jones, G.W.; Kavanagh, K.; Doyle, S. Gliotoxin effects on fungal growth: Mechanisms and exploitation. Fungal Genet. Biol. 2012, 49, 302–312. [Google Scholar] [CrossRef] [Green Version]
- Anaam, F.; Pirko, E.Y.; Abaas, A.F.; Al-aajem, B.M.R. Antimicrobial Activity of Aspergillus Gliotoxin on S.aureus in Diabetic’s Patient’s Type-2. Br. J. Med Health Res. 2018, 5, 1–5. [Google Scholar] [CrossRef]
- Li, X.; Kim, S.K.; Nam, K.W.; Kang, J.S.; Choi, H.D.; Son, B.W. A new antibacterial dioxopiperazine alkaloid related to gliotoxin from a marine isolate of the fungus Pseudallescheria. J. Antibiot. 2006, 59, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Gardete, S.; Tomasz, A. Mechanisms of vancomycin resistance in Staphylococcus aureus. J. Clin. Investig. 2014, 124, 2836–2840. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Kim, E.S.; Kim, H.S.; Park, S.J.; Bang, K.M.; Park, H.J.; Park, S.Y.; Moon, S.M.; Chong, Y.P.; Kim, S.H.; et al. Comparison of the clinical features, bacterial genotypes and outcomes of patients with bacteraemia due to heteroresistant vancomycin-intermediate Staphylococcus aureus and vancomycin-susceptible S. aureus. J. Antimicrob. Chemother. 2012, 67, 1843–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridkin, S.K.; Hageman, J.; McDougal, L.K.; Mohammed, J.; Jarvis, W.R.; Perl, T.M.; Tenover, F.C. Epidemiological and microbiological characterization of infections caused by Staphylococcus aureus with reduced susceptibility to vancomycin, United States, 1997–2001. Clin. Infect. Dis. 2003, 36, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Howden, B.P.; Davies, J.K.; Johnson, P.D.; Stinear, T.P.; Grayson, M.L. Reduced vancomycin susceptibility in Staphylococcus aureus, including vancomycin-intermediate and heterogeneous vancomycin-intermediate strains: Resistance mechanisms, laboratory detection, and clinical implications. Clin. Microbiol. Rev. 2010, 23, 99–139. [Google Scholar] [CrossRef] [Green Version]
- Liñares, J. The VISA/GISA problem: Therapeutic implications. Clin. Microbiol. Infect. 2001, 7, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Pang, L.; Ma, X.X.; Hu, J.; Tian, Y.; Yang, Y.L.; Sun, D.D. Phenotypic and Molecular Characterisation of Staphylococcus Aureus with Reduced Vancomycin Susceptibility Derivated in Vitro. Open Med. 2018, 13, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Pardo, J.; Urban, C.; Galvez, E.M.; Ekert, P.G.; Müller, U.; Kwon-Chung, J.; Lobigs, M.; Müllbacher, A.; Wallich, R.; Borner, C.; et al. The mitochondrial protein Bak is pivotal for gliotoxin-induced apoptosis and a critical host factor of Aspergillus fumigatus virulence in mice. J. Cell Biol. 2006, 174, 509–519. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing (EUCAST). Disk Diffusion Method for Antimicrobial Susceptibility Testing, Version 8.0, 2020. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Disk_test_documents/2021_manuals/Reading_guide_v_8.0_EUCAST_Disk_Test_2021.pdf (accessed on 23 January 2021).
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk Susceptibility Tests, 12th ed.; Approved Standard; Supplement M02-A12; CLSI: Wayne, PA, USA, 2015. [Google Scholar]
- Clinical and Laboratory Standard Institute. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-First Informational Supplement M100-S21; CLSI: Wayne, PA, USA, 2011. [Google Scholar]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, T.; Hayball, M. CalcuSyn for Windows: Multiple-Drug Dose-Effect Analyzer and Manual; Biosoft: Cambridge, UK, 1996. [Google Scholar]
- Dong, Y.; Zhao, X.; Domagala, J.; Drlica, K. Effect of fluoroquinolone concentration on selection of resistant mutants of Mycobacterium bovis BCG and Staphylococcus aureus. Antimicrob. Agents Chemother. 1999, 43, 1756–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondeau, J.M. New concepts in antimicrobial susceptibility testing: The mutant prevention concentration and mutant selection window approach. Vet. Dermatol. 2009, 20, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Beanan, M.J.; Strome, S. Characterization of a germ-line proliferation mutation in C. elegans. Development 1992, 116, 755–766. [Google Scholar]
- Kong, C.; Yehye, W.A.; Abd Rahman, N.; Tan, M.W.; Nathan, S. Discovery of potential anti-infectives against Staphylococcus aureus using a Caenorhabditis elegans infection model. BMC Complement. Altern. Med. 2014, 14, 4. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Individually | Combination | |
|---|---|---|---|
| Vancomycin | 64 | 0 | 32 |
| Gliotoxin | 0 | 256 | 2 |
| Range of combination index (CI) | Symbol | Description |
|---|---|---|
| <0.1 | +++++ | Very strong synergism |
| 0.1–0.3 | ++++ | Strong synergism |
| 0.3–0.7 | +++ | Synergism |
| 0.7–0.85 | ++ | Moderate synergism |
| 0.85–0.90 | + | Slight synergism |
| 0.90–1.10 | ± | Nearly additive |
| 1.10–1.20 | – | Slight antagonism |
| 1.20–1.45 | – – | Moderate antagonism |
| 1.45–3.3 | – – – | Antagonism |
| 3.3–10 | – – – – | Strong antagonism |
| >10 | – – – – – | Very strong antagonism |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esteban, P.; Redrado, S.; Comas, L.; Domingo, M.P.; Millán-Lou, M.I.; Seral, C.; Algarate, S.; Lopez, C.; Rezusta, A.; Pardo, J.; et al. In Vitro and In Vivo Antibacterial Activity of Gliotoxin Alone and in Combination with Antibiotics against Staphylococcus aureus. Toxins 2021, 13, 85. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020085
Esteban P, Redrado S, Comas L, Domingo MP, Millán-Lou MI, Seral C, Algarate S, Lopez C, Rezusta A, Pardo J, et al. In Vitro and In Vivo Antibacterial Activity of Gliotoxin Alone and in Combination with Antibiotics against Staphylococcus aureus. Toxins. 2021; 13(2):85. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020085
Chicago/Turabian StyleEsteban, Patricia, Sergio Redrado, Laura Comas, M. Pilar Domingo, M. Isabel Millán-Lou, Cristina Seral, Sonia Algarate, Concepción Lopez, Antonio Rezusta, Julian Pardo, and et al. 2021. "In Vitro and In Vivo Antibacterial Activity of Gliotoxin Alone and in Combination with Antibiotics against Staphylococcus aureus" Toxins 13, no. 2: 85. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020085