Characteristic Distribution of Ciguatoxins in the Edible Parts of a Grouper, Variola louti

 , and
, and
Abstract
:1. Introduction
2. Results
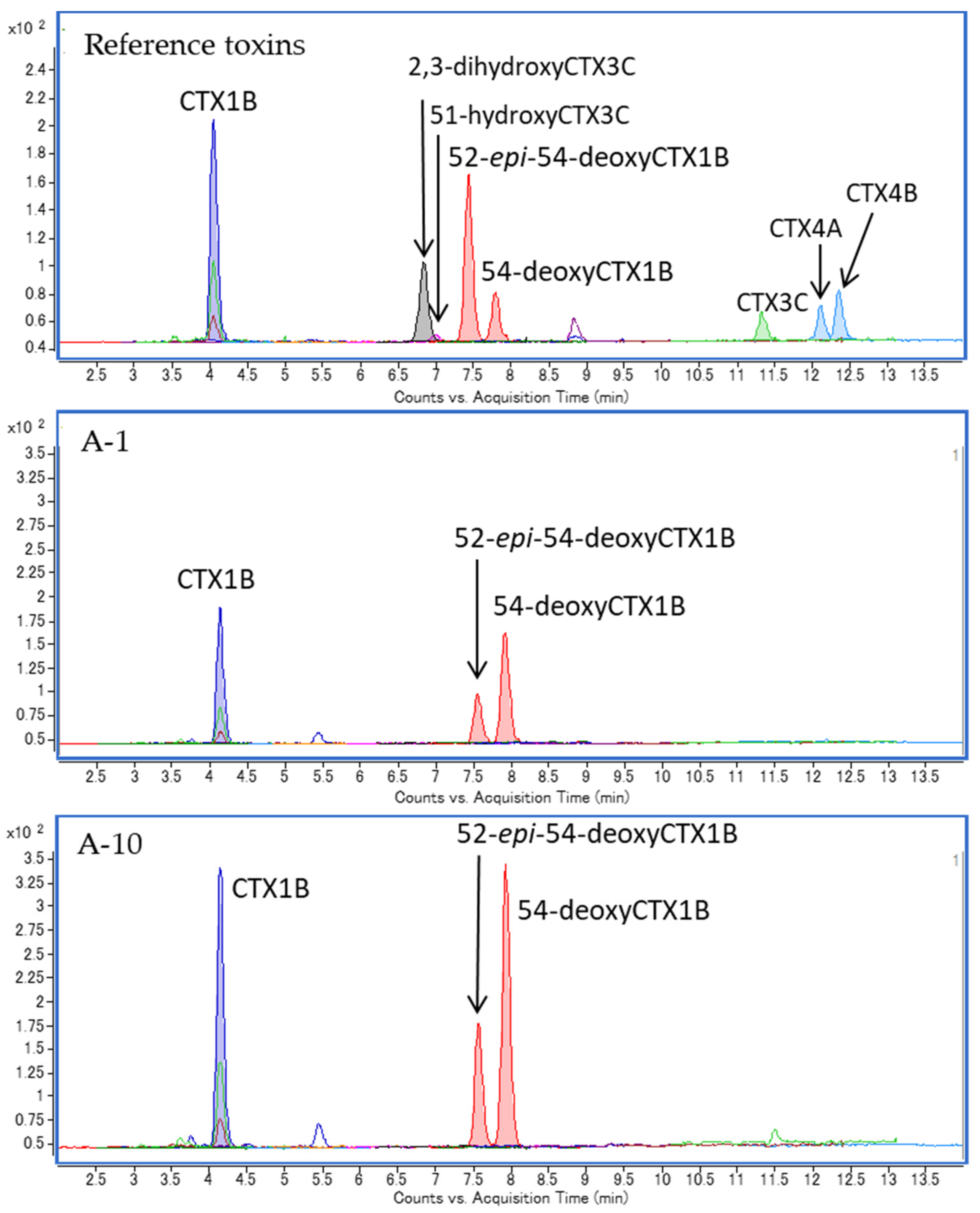
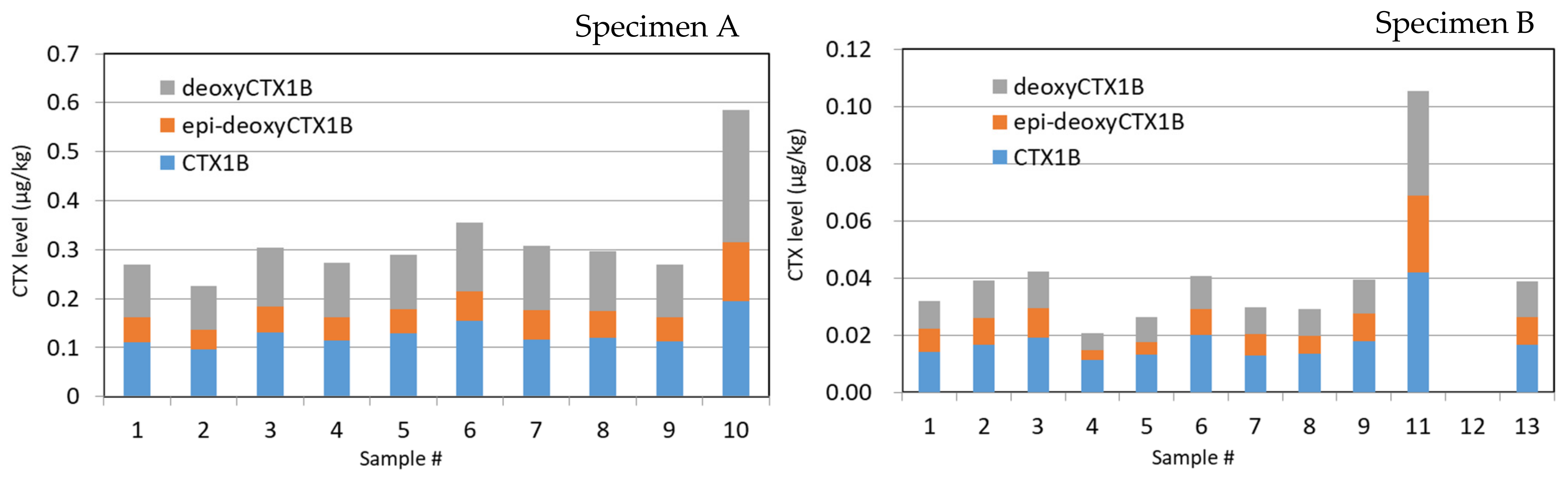
2.1. Specimen A
2.2. Specimen B
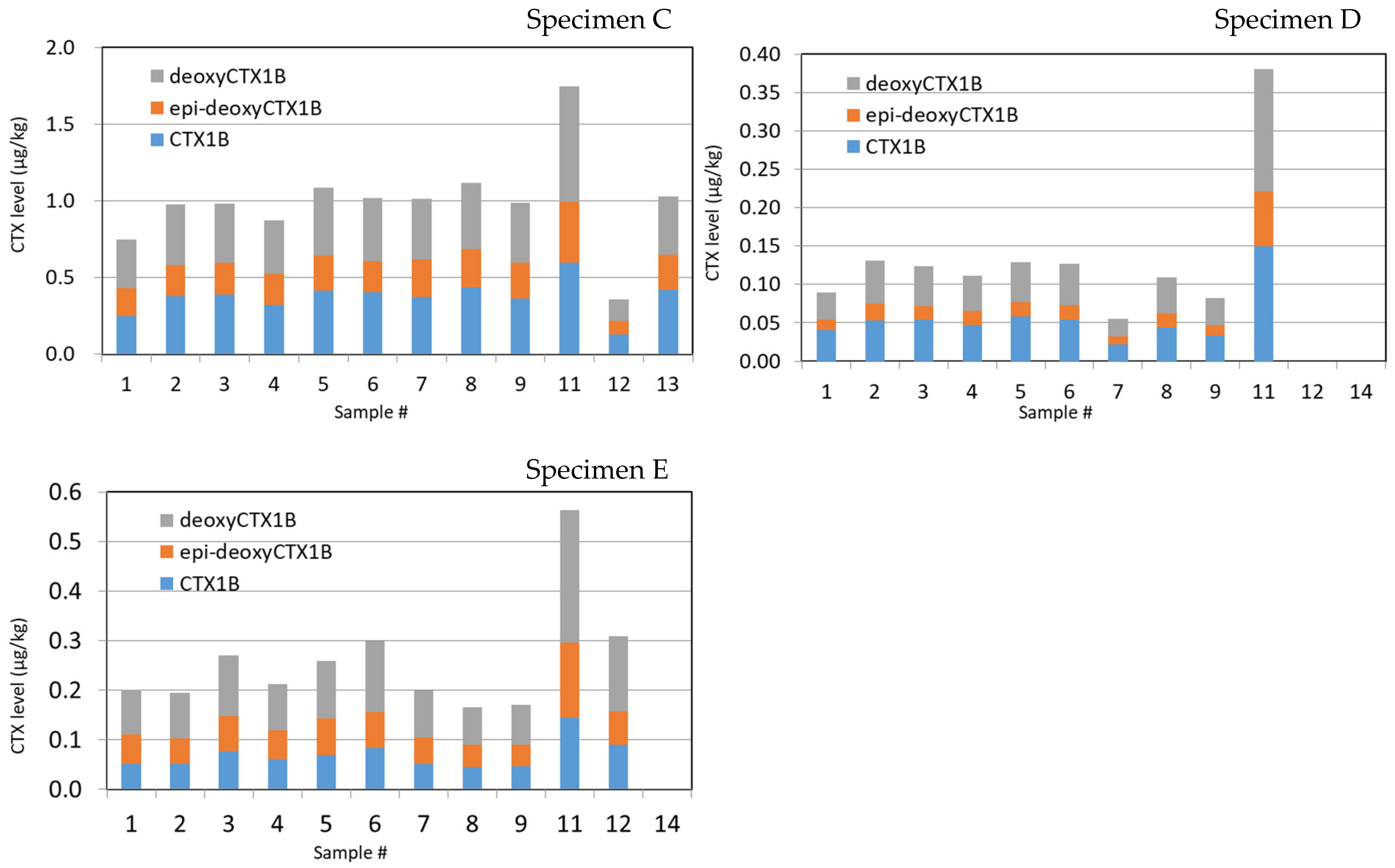
2.3. Specimens C
2.4. Specimens D
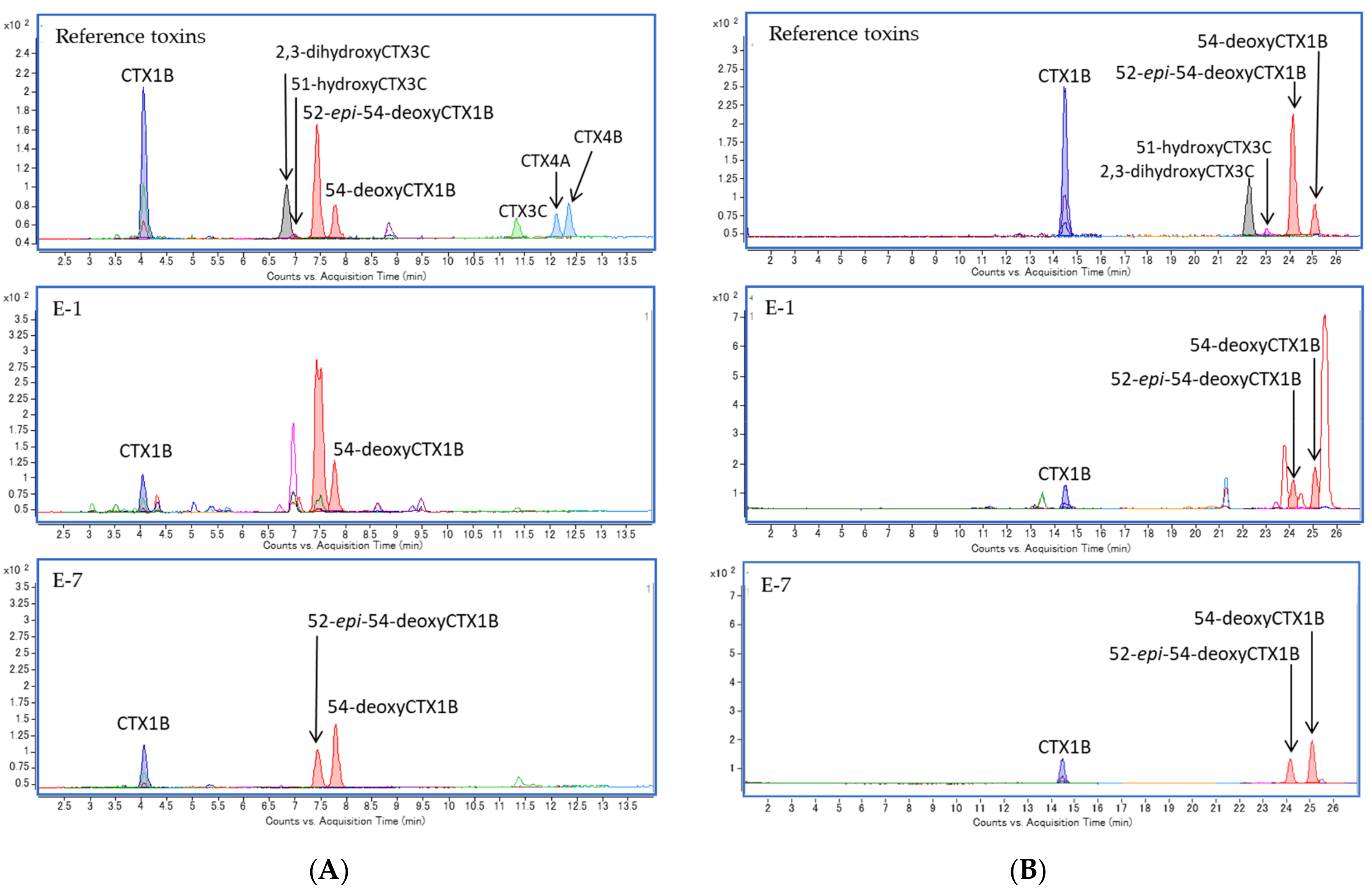
2.5. Specimen E
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reference Toxins and Reagents
5.2. Fish Specimens
5.3. Extraction and Sample Preparation for LC-MS/MS
5.4. LC-MS/MS Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO; WHO. Report of the Expert Meeting on Ciguatera Poisoning: Rome, 19–23 November 2018; FAO: Rome, Italy, 2020; Volume 9, p. 156. [Google Scholar] [CrossRef]
- Yasumoto, T.; Satake, M. Chemistry, Etiology and Determination Methods of Ciguatera Toxins. J. Toxicol. Toxin Rev. 1996, 15, 91–107. [Google Scholar] [CrossRef]
- Yasumoto, T. Chemistry, etiology, and food chain dynamics of marine toxins. Proc. Jpn. Acad. Ser. B 2005, 81, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An Updated Review of Ciguatera Fish Poisoning: Clinical, Epidemiological, Environmental, and Public Health Management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef]
- Pearn, J. Neurology of ciguatera. J. Neurol. Neurosurg. Psychiatry 2001, 70, 4–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinain, M.; Gatti, C.; Darius, H.; Quod, J.-P.; Tester, P. Ciguatera poisonings: A global review of occurrences and trends. Harmful Algae 2020, 101873. [Google Scholar] [CrossRef]
- Pérez-Arellano, J.-L.; Luzardo, O.P.; Brito, A.P.; Cabrera, M.H.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera Fish Poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Henao, A.; García-Álvarez, N.; Sergent, F.S.; Estévez, P.; Gago-Martínez, A.; Martín, F.; Ramos-Sosa, M.; Fernández, A.; Diogène, J.; Real, F. Presence of CTXs in moray eels and dusky groupers in the marine environment of the Canary Islands. Aquat. Toxicol. 2020, 221, 105427. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Konosu, S.; Yasumoto, T.; Kamiya, H. Ciguatera in the Ryukyu and Amami Islands. Bull. Jpn. Soc. Sci. Fish. 1969, 35, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Oshiro, N.; Yogi, K.; Asato, S.; Sasaki, T.; Tamanaha, K.; Hirama, M.; Yasumoto, T.; Inafuku, Y. Ciguatera incidence and fish toxicity in Okinawa, Japan. Toxicon 2010, 56, 656–661. [Google Scholar] [CrossRef]
- Oshiro, N.; Matsuo, T.; Sakugawa, S.; Yogi, K.; Matsuda, S.; Yasumoto, T.; Inafuku, Y. Ciguatera Fish Poisoning on Kakeroma Island, Kagoshima Prefecture, Japan. Trop. Med. Health 2011, 39, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Toda, M.; Uneyama, C.; Toyofuku, H.; Morikawa, K. Trends of Food Poisonings Caused by Natural Toxins in Japan, 1989–2011. Shokuhin Eiseigaku Zasshi (Food Hyg. Saf. Sci.) 2012, 53, 105–120. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, T.; Nakajima, I.; Bagnis, R.; Adachi, R. Finding of a dinoflagellate as a likely culprit of ciguatera. Bull. Jpn. Soc. Sci. Fish. 1977, 43, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Chinain, M.; Gatti, C.M.; Roué, M.; Darius, H.T.; Subba Rao, D. Ciguatera-causing dinoflagellates in the genera Gambierdiscus and Fukuyoa: Distribution, ecophysiology and toxicology. In Dinoflagellates: Classification, Evolution, Physiology and Ecological Significance; Durvasula, S.R.V., Ed.; Nova Science Publishers: New York, NY, USA, 2020; pp. 405–457. ISBN 978-1-53617-888-3. [Google Scholar]
- Yasumoto, T.; Igarashi, T.; Legrand, A.-M.; Cruchet, P.; Chinain, M.; Fujita, T.; Naoki, H. Structural Elucidation of Ciguatoxin Congeners by Fast-Atom Bombardment Tandem Mass Spectroscopy. J. Am. Chem. Soc. 2000, 122, 4988–4989. [Google Scholar] [CrossRef]
- Yogi, K.; Oshiro, N.; Inafuku, Y.; Hirama, M.; Yasumoto, T. Detailed LC-MS/MS Analysis of Ciguatoxins Revealing Distinct Regional and Species Characteristics in Fish and Causative Alga from the Pacific. Anal. Chem. 2011, 83, 8886–8891. [Google Scholar] [CrossRef] [PubMed]
- Yogi, K.; Sakugawa, S.; Oshiro, N.; Ikehara, T.; Sugiyama, K.; Yasumoto, T. Determination of Toxins Involved in Ciguatera Fish Poisoning in the Pacific by LC/MS. J. AOAC Int. 2014, 97, 398–402. [Google Scholar] [CrossRef]
- Oshiro, N.; Tomikawa, T.; Kuniyoshi, K.; Kimura, K.; Kojima, T.; Yasumoto, T.; Asakura, H. Detection of Ciguatoxins from Fish Introduced into a Wholesale Market in Japan. Shokuhin Eiseigaku Zasshi (Food Hyg. Saf. Sci.) 2021, 62, 8–13. [Google Scholar] [CrossRef]
- Oshiro, N.; Tomikawa, T.; Kuniyoshi, K.; Ishikawa, A.; Toyofuku, H.; Kojima, T.; Asakura, H. LC–MS/MS Analysis of Ciguatoxins Revealing the Regional and Species Distinction of Fish in the Tropical Western Pacific. J. Mar. Sci. Eng. 2021, 9, 299. [Google Scholar] [CrossRef]
- Chateau-Degat, M.-L.; Legrand, A.-M.; Darius, T.; Chansin, R.; Dewailly, E.; Laudon, F.; Chinain, M.; Huin-Blondey, M.-O.; Nguyen, N.L. Prevalence of Chronic Symptoms of Ciguatera Disease in French Polynesian Adults. Am. J. Trop. Med. Hyg. 2007, 77, 842–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, A.; Zammit, A.; Farrell, H. Four recent ciguatera fish poisoning incidents in New South Wales, Australia linked to imported fish. Commun. Dis. Intell. 2019, 43, 43. [Google Scholar] [CrossRef]
- Chateau-Degat, M.-L.; Dewailly, E.; Cerf, N.; Nguyen, N.L.; Huin-Blondey, M.-O.; Hubert, B.; Laudon, F.; Chansin, R. Temporal trends and epidemiological aspects of ciguatera in French Polynesia: A 10-year analysis. Trop. Med. Int. Health 2007, 12, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Muecke, C.; Hamper, L.; Skinner, A.; Osborne, C. Ciguatera fish poisoning in an international ship crew in Saint John Canada: 2015. Can. Commun. Dis. Rep. 2015, 41, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.Y.K. Characteristic Features and Contributory Factors in Fatal Ciguatera Fish Poisoning—Implications for Prevention and Public Education. Am. J. Trop. Med. Hyg. 2016, 94, 704–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, Y.; Fusetani, N. A Preliminary Report on the Toxicity of an Amberjack, Seriola aureovittata. Bull. Jpn. Soc. Sci. Fish. 1968, 34, 618–626. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, T.; Scheuer, P.J. Marine toxins of the pacific—VIII ciguatoxin from moray eel livers. Toxicon 1969, 7, 273–276. [Google Scholar] [CrossRef]
- Dickey, R.W.; Plakas, S.M. Ciguatera: A public health perspective. Toxicon 2010, 56, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H.; Mak, Y.L.; Wu, J.J.; Jin, L.; Sit, W.H.; Lam, J.C.W.; De Mitcheson, Y.S.; Chan, L.L.; Lam, P.K.S.; Murphy, M.B. Spatial distribution of ciguateric fish in the Republic of Kiribati. Chemosphere 2011, 84, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mak, Y.L.; Chang, Y.-H.; Xiao, C.; Chen, Y.-M.; Shen, J.; Wang, Q.; Ruan, Y.; Lam, P.K.S. Uptake and Depuration Kinetics of Pacific Ciguatoxins in Orange-Spotted Grouper (Epinephelus coioides). Environ. Sci. Technol. 2020, 54, 4475–4483. [Google Scholar] [CrossRef]
- Kohli, G.S.; Haslauer, K.; Sarowar, C.; Kretzschmar, A.L.; Boulter, M.; Harwood, D.; Laczka, O.; Murray, S.A. Qualitative and quantitative assessment of the presence of ciguatoxin, P-CTX-1B, in Spanish Mackerel (Scomberomorus commerson) from waters in New South Wales (Australia). Toxicol. Rep. 2017, 4, 328–334. [Google Scholar] [CrossRef]
- Legrand, A.M.; Litaudon, M.; Genthon, J.N.; Bagnis, R.; Yasumoto, T. Isolation and some properties of ciguatoxin. Environ. Boil. Fishes 1989, 1, 183–188. [Google Scholar] [CrossRef]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Sawada, T.; Takahashi, K.; Hatano, M. Molecular Species Analysis of Fish Oil Triglyceride by Light Scattering Mass Detector Equipped Liquid Chromatograph II. Triglyceride Composition of Tuna and Bonito Orbital Fats. Nippon Suisan Gakkaishi 1993, 59, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, K.; Saito, H. The Docosahexaenoic Acid Content of the Lipid of Juvenile Bluefin Tuna Thunnus thynnus Caught in the Sea off Japanese Coast. Fish. Sci. 1996, 62, 840–841. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, Y.; Nomura, A.; Wada, S. Seasonal Variation in Lipid Content and Fatty Acid Composition of Round Frigate Mackerel, Auxis rochei Risso, in Tosa Bay. J. Jpn. Oil Chem. Soc. 1996, 45, 1267–1270. [Google Scholar] [CrossRef]
- Wang, J.; Ou, S.-W.; Wang, Y.-J. Distribution and function of voltage-gated sodium channels in the nervous system. Channels 2017, 11, 534–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Yasumoto, T. Structures of ciguatoxin and its congener. J. Am. Chem. Soc. 1989, 111, 8929–8931. [Google Scholar] [CrossRef]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Fukui, M.; Yasumoto, T. Structures and configurations of ciguatoxin from the moray eel Gymnothorax javanicus and its likely precursor from the dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. The structure of CTX3C, a ciguatoxin congener isolated from cultured Gambierdiscus toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- Satake, M.; Ishibashi, Y.; Legrand, A.-M.; Yasumoto, T. Isolation and Structure of Ciguatoxin-4A, a New Ciguatoxin Precursor, from Cultures of Dinoflagellate Gambierdiscus toxicus and Parrotfish Scarus gibbus. Biosci. Biotechnol. Biochem. 1996, 60, 2103–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satake, M.; Fukui, M.; Legrand, A.-M.; Cruchet, P.; Yasumoto, T. Isolation and structures of new ciguatoxin analogs, 2,3-dihydroxyCTX3C and 51-hydroxyCTX3C, accumulated in tropical reef fish. Tetrahedron Lett. 1998, 39, 1197–1198. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen | ID 1 | SL 2 (mm) | Weight (g) | Eye Sample Preparation 3 |
|---|---|---|---|---|
| A | 160024 | 490 | 2934 | whole parts were mixed (#10) |
| B | 160085 | 410 | 1734 | surrounding tissue (#11), inner contents, including a lens (#12) and the outer membrane of the eyeball (#13) |
| C | 163077 | 456 | 2719 | surrounding tissue (#11), inner contents, including a lens (#12) and the outer membrane of the eyeball (#13) |
| D | 160135 | 490 | 2594 | surrounding tissue (#11), inner contents of the eyeball (#12) and lens (#14) |
| E | 160136 | 450 | 2430 | surrounding tissue (#11), inner contents of the eyeball (#12) and lens (#14) |
| Specimen | CTXs Levels (μg/kg) in the Fillet 1 | CTXs Levels (μg/kg) in the Head 1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| #1 | #2 | #3 | #4 | #5 | #6 | Average | SD 2 | #7 | #8 | #9 | Average | SD | |
| A | 0.270 | 0.226 | 0.305 | 0.274 | 0.290 | 0.355 | 0.287 | 0.043 | 0.308 | 0.297 | 0.270 | 0.292 | 0.019 |
| B | 0.032 | 0.039 | 0.042 | 0.021 | 0.026 | 0.041 | 0.033 | 0.009 | 0.030 | 0.029 | 0.039 | 0.033 | 0.006 |
| C | 0.749 | 0.978 | 0.984 | 0.873 | 1.088 | 1.018 | 0.948 | 0.120 | 1.011 | 1.117 | 0.989 | 1.039 | 0.068 |
| D | 0.089 | 0.131 | 0.124 | 0.111 | 0.129 | 0.126 | 0.118 | 0.016 | 0.055 | 0.110 | 0.082 | 0.082 | 0.027 |
| E | 0.201 | 0.195 | 0.270 | 0.212 | 0.259 | 0.300 | 0.239 | 0.043 | 0.200 | 0.166 | 0.171 | 0.179 | 0.018 |
| Specimen | CTXs Levels (μg/kg) 1 | ||||||
|---|---|---|---|---|---|---|---|
| Fillet 2 | Head 3 | #10 | #11 | #12 | #13 | #14 | |
| A | 0.287 | 0.292 | 0.585 | - 4 | - | - | - |
| B | 0.033 | 0.033 | - | 0.106 | <LOD 5 | 0.039 | - |
| C | 0.948 | 1.039 | - | 1.747 | 0.360 | 1.031 | - |
| D | 0.118 | 0.082 | - | 0.380 | <LOD | - | <LOD |
| E | 0.239 | 0.179 | - | 0.563 | 0.308 | - | <LOD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oshiro, N.; Nagasawa, H.; Kuniyoshi, K.; Kobayashi, N.; Sugita-Konishi, Y.; Asakura, H.; Yasumoto, T. Characteristic Distribution of Ciguatoxins in the Edible Parts of a Grouper, Variola louti. Toxins 2021, 13, 218. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13030218
Oshiro N, Nagasawa H, Kuniyoshi K, Kobayashi N, Sugita-Konishi Y, Asakura H, Yasumoto T. Characteristic Distribution of Ciguatoxins in the Edible Parts of a Grouper, Variola louti. Toxins. 2021; 13(3):218. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13030218
Chicago/Turabian StyleOshiro, Naomasa, Hiroya Nagasawa, Kyoko Kuniyoshi, Naoki Kobayashi, Yoshiko Sugita-Konishi, Hiroshi Asakura, and Takeshi Yasumoto. 2021. "Characteristic Distribution of Ciguatoxins in the Edible Parts of a Grouper, Variola louti" Toxins 13, no. 3: 218. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13030218