Potency Testing of Venoms and Antivenoms in Embryonated Eggs: An Ethical Alternative to Animal Testing

Abstract
:1. Introduction
2. Results
2.1. Feasibility of the Embryonated Egg Model
2.2. Measurement of AV Potency
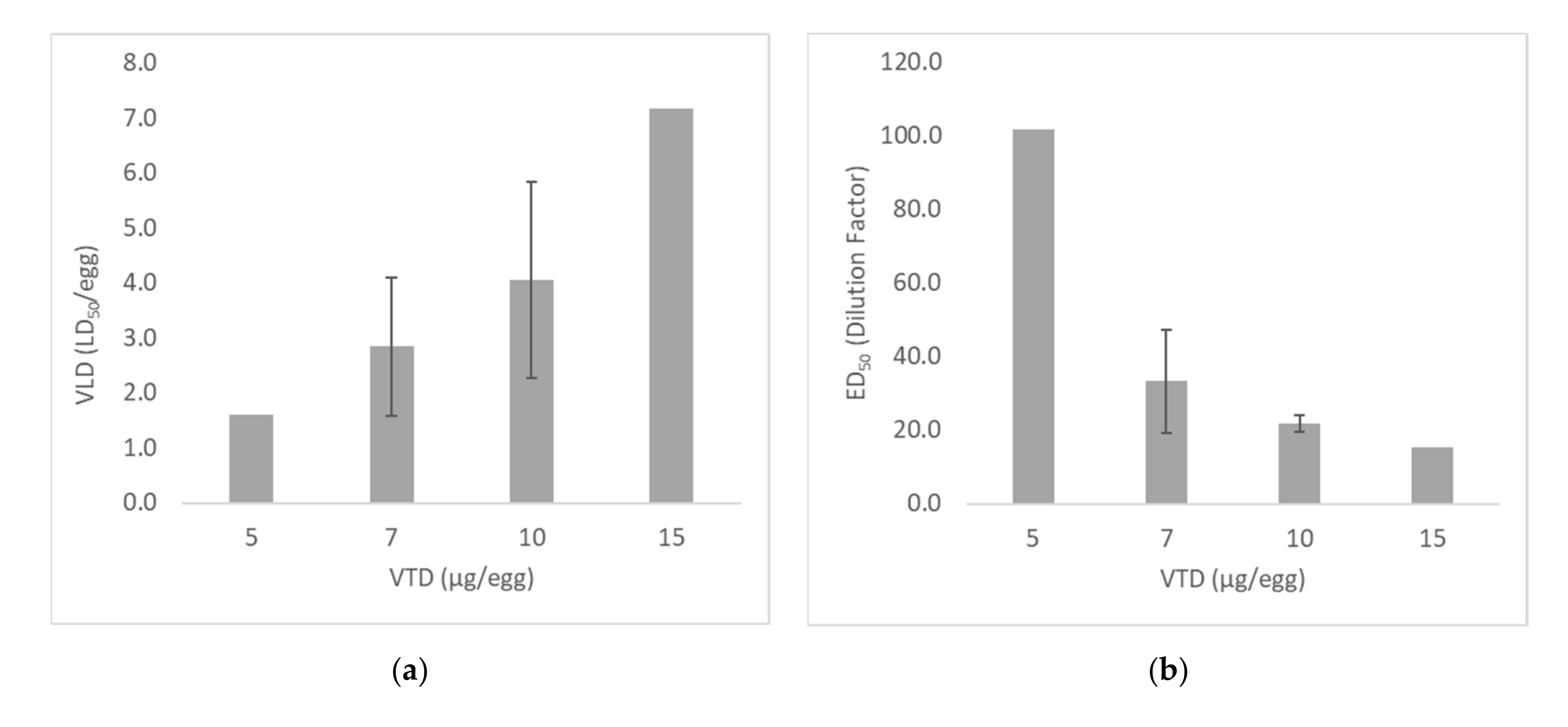
2.2.1. Optimization of Venom Test Dose
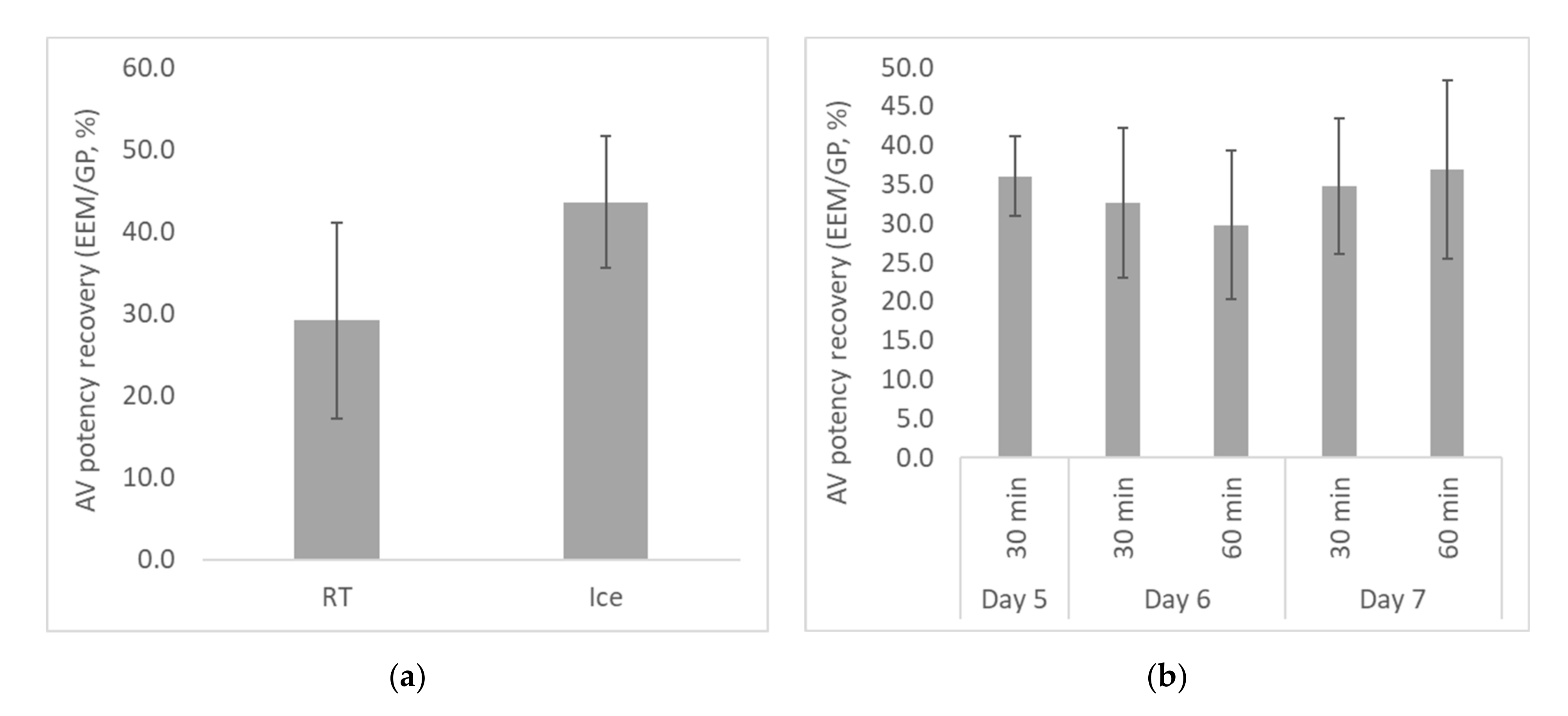
2.2.2. Optimization of Egg Age and Venom/AV Incubation Conditions
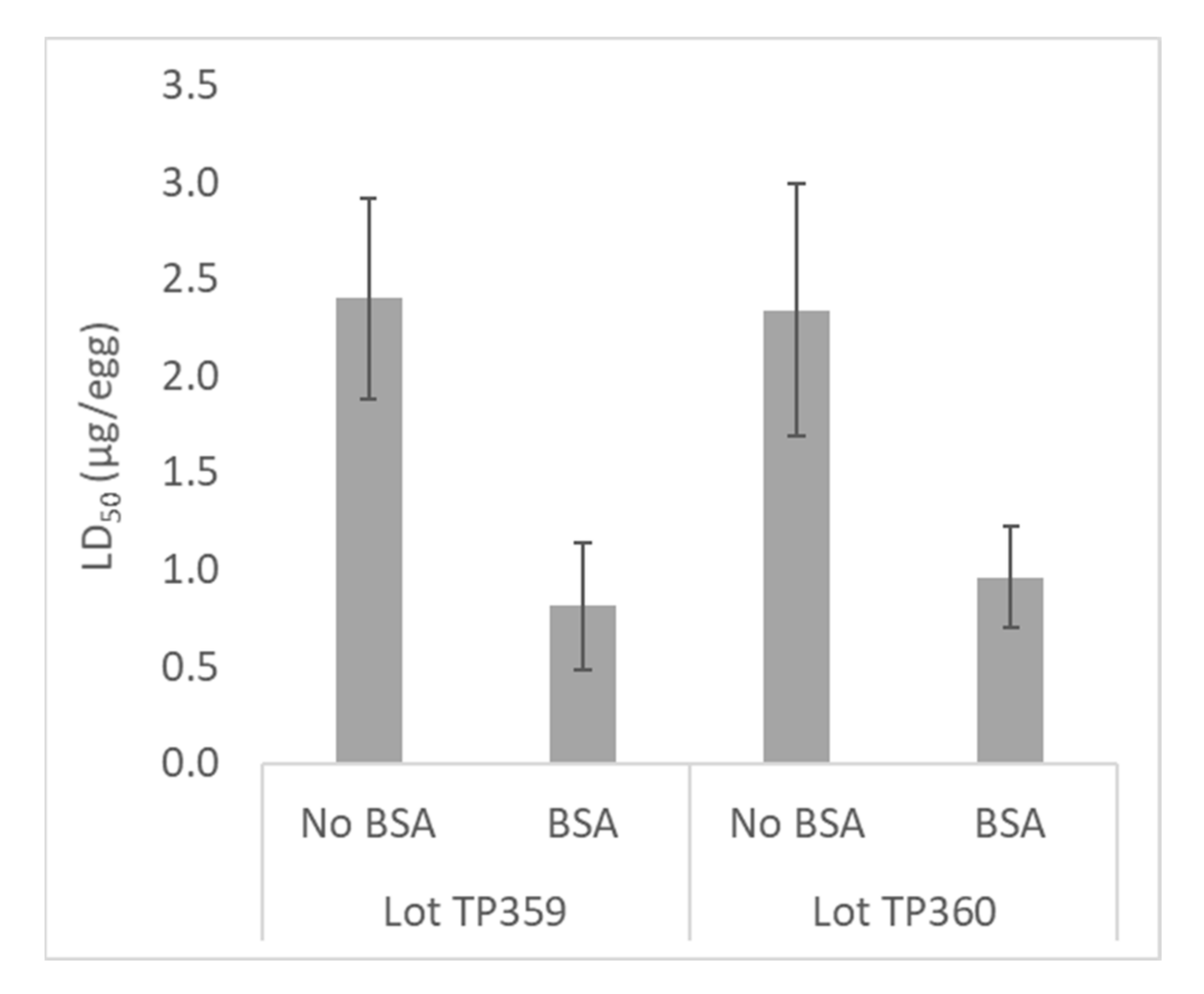
2.2.3. Optimization of Venom Potency by Addition of BSA
2.3. Performance of the EEM Assay
2.3.1. Measurement of AV Potency
2.3.2. Breadth of Venom Activity in the EEM
3. Discussion
4. Materials and Methods
4.1. Venoms, AVs, and Eggs
4.2. Assessment of Venom Potency (LD50)
4.3. Assessment of AV Potency (ED50)
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badre, S. Bioactive toxins from stinging jellyfish. Toxicon 2014, 91, 114–125. [Google Scholar] [CrossRef]
- Barber, C.M.; Madaras, F.; Turnbull, R.K.; Morley, T.; Dunstan, N.; Allen, L.; Kuchel, T.; Mirtschin, P.; Hodgson, W.C. Comparative studies of the venom of a new Taipan species, Oxyuranus temporalis, with other members of its genus. Toxins 2014, 6, 1979–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkman, D.L.; Burnell, J.N. Biochemical and molecular characterisation of cubozoan protein toxins. Toxicon 2009, 54, 1162–1173. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.L.; Jia, X.; Potriquet, J.; Kumar, D.; Dash, D.; Kvaskoff, D.; Mulvenna, J. Transcriptome and venom proteome of the box jellyfish Chironex fleckeri. BMC Genom. 2015, 16, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkman, D.L.; Konstantakopoulos, N.; McInerney, B.V.; Mulvenna, J.; Seymour, J.E.; Isbister, G.K.; Hodgson, W.C. Chironex fleckeri (box jellyfish) venom proteins: Expansion of a cnidarian toxin family that elicits variable cytolytic and cardiovascular effects. J. Biol. Chem. 2014, 289, 4798–4812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgieva, D.; Seifert, J.; Ohler, M.; von Bergen, M.; Spencer, P.; Arni, R.K.; Genov, N.; Betzel, C. Pseudechis australis venomics: Adaptation for a defense against microbial pathogens and recruitment of body transferrin. J. Proteome Res. 2011, 10, 2440–2464. [Google Scholar] [CrossRef]
- Jackson, T.N.; Koludarov, I.; Ali, S.A.; Dobson, J.; Zdenek, C.N.; Dashevsky, D.; Op den Brouw, B.; Masci, P.P.; Nouwens, A.; Josh, P.; et al. Rapid Radiations and the Race to Redundancy: An Investigation of the Evolution of Australian Elapid Snake Venoms. Toxins 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Casewell, N.R.; Yanagihara, A.A.; Nouwens, A.; Cribb, B.W.; Whitehead, D.; Jackson, T.N.; Ali, S.A.; Wagstaff, S.C.; Koludarov, I.; et al. Firing the sting: Chemically induced discharge of cnidae reveals novel proteins and peptides from box jellyfish (Chironex fleckeri) venom. Toxins 2015, 7, 936–950. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, G.M.; Little, M.J.; Birinyi-Strachan, L.C. Structure and function of δ-atracotoxins: Lethal neurotoxins targeting the voltage-gated sodium channel. Toxicon 2004, 43, 587–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, V.; White, J.; Schwaner, T.D.; Sparrow, A. Variation in venom proteins from isolated populations of tiger snakes (Notechis ater niger, N. scutatus) in South Australia. Toxicon 1988, 26, 1067–1075. [Google Scholar] [CrossRef]
- Ziegman, R.; Undheim, E.A.B.; Baillie, G.; Jones, A.; Alewood, P.F. Investigation of the estuarine stonefish (Synanceia horrida) venom composition. J. Proteom. 2019, 201, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Ziegman, R.; Alewood, P. Bioactive components in fish venoms. Toxins 2015, 7, 1497–1531. [Google Scholar] [CrossRef]
- World Health Organisation. Annex 5: Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; WHO Technical Report Series, No. 1004; World Health Organisation: Geneva, Switzerland, 2017. [Google Scholar]
- NHMRC. Australian Code for the Care and Use of Animals for Scientific Purposes, 8th ed.; National Health and Medical Research Council: Canberra, Australia, 2013.
- Prevention of Cruelty to Animals (POCTA) Act. Available online: https://agriculture.vic.gov.au/livestock-and-animals/animal-welfare-victoria/pocta-act-1986 (accessed on 31 August 2020).
- Mellor, D.J.; Diesch, T.J. Birth and hatching: Key events in the onset of awareness in the lamb and chick. N. Z. Vet. J. 2007, 55, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.L.; Mellor, D.J.; Sandøe, P. How should the welfare of fetal and neurologically immature postnatal animals be protected? Anim. Welf. 2014, 23, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Leary, S.; Underwood, W.; Anthony, R.; Cartner, S.; Grandin, T.; Greenacre, C.; Gwaltney-Brant, S.; McCrackin, M.; Meyer, R.; Miller, D. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition; AVMA American Veterinary Medical Association: Schaumburg, IL, USA, 2020. [Google Scholar]
- Bjornstad, S.; Austdal, L.P.; Roald, B.; Glover, J.C.; Paulsen, R.E. Cracking the Egg: Potential of the Developing Chicken as a Model System for Nonclinical Safety Studies of Pharmaceuticals. J. Pharmacol. Exp. Ther. 2015, 355, 386–396. [Google Scholar] [CrossRef] [Green Version]
- Kain, K.H.; Miller, J.W.; Jones-Paris, C.R.; Thomason, R.T.; Lewis, J.D.; Bader, D.M.; Barnett, J.V.; Zijlstra, A. The chick embryo as an expanding experimental model for cancer and cardiovascular research. Dev. Dyn. 2014, 243, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Mathisen, G.H.; Yazdani, M.; Rakkestad, K.E.; Aden, P.K.; Bodin, J.; Samuelsen, M.; Nygaard, U.C.; Goverud, I.L.; Gaarder, M.; Løberg, E.M. Prenatal exposure to bisphenol A interferes with the development of cerebellar granule neurons in mice and chicken. Int. J. Dev. Neurosci. 2013, 31, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowska, P.; Tayoun, T.; Oulhen, M.; Faugeroux, V.; Rouffiac, V.; Aberlenc, A.; Pommier, A.L.; Honore, A.; Marty, V.; Bawa, O.; et al. Exploitation of the chick embryo chorioallantoic membrane (CAM) as a platform for anti-metastatic drug testing. Sci. Rep. 2020, 10, 16876. [Google Scholar] [CrossRef]
- Vargas, A.; Zeisser-Labouèbe, M.; Lange, N.; Gurny, R.; Delie, F. The chick embryo and its chorioallantoic membrane (CAM) for the in vivo evaluation of drug delivery systems. Adv. Drug Deliv. Rev. 2007, 59, 1162–1176. [Google Scholar] [CrossRef]
- Sells, P.G.; Ioannou, P.; Theakston, R.D.G. A humane alternative to the measurement of the lethal effects (ld50) of non-neurotoxic venoms using hens’ eggs. Toxicon 1998, 36, 985–991. [Google Scholar] [CrossRef]
- Sells, P.G.; Laing, G.D.; Theakston, R.D.G. An in vivo but insensate model for the evaluation of antivenoms (ED50) using fertile hens’ eggs. Toxicon 2001, 39, 665–668. [Google Scholar] [CrossRef]
- Sells, P.G.; Richards, A.M.; Laing, G.D.; Theakston, R.D.G. The use of hens’ eggs as an alternative to the conventional in vivo rodent assay for antidotes to haemorrhagic venoms. Toxicon 1997, 35, 1413–1421. [Google Scholar] [CrossRef]
- Goebel-Stengel, M.; Stengel, A.; Taché, Y.; Reeve, J.R., Jr. The importance of using the optimal plasticware and glassware in studies involving peptides. Anal. Biochem. 2011, 414, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Broad, A.J.; Sutherland, S.K.; Coulter, A.R. The lethality in mice of dangerous Australian and other snake venom. Toxicon 1979, 17, 661–664. [Google Scholar] [CrossRef]
- Wiener, S. The Sydney funnel-web spider (Atrax robustus). I. Collection of venom and its toxicity in animals. Med J. Aust. 1957, 44, 377–383. [Google Scholar] [CrossRef]
- Herzig, V.; Sunagar, K.; Wilson, D.T.R.; Pineda, S.S.; Israel, M.R.; Dutertre, S.; McFarland, B.S.; Undheim, E.A.B.; Hodgson, W.C.; Alewood, P.F.; et al. Australian funnel-web spiders evolved human-lethal δ-hexatoxins for defense against vertebrate predators. Proc. Natl. Acad. Sci. USA 2020, 117, 24920–24928. [Google Scholar] [CrossRef] [PubMed]
- Habermann, E.; Cheng-Raude, D. Central neurotoxicity of apamin, crotamin, phospholipase A and alpha-amanitin. Toxicon 1975, 13, 465–473. [Google Scholar] [CrossRef]
- Healy, K.; Carbone, C.; Jackson, A.L. Snake venom potency and yield are associated with prey-evolution, predator metabolism and habitat structure. Ecol. Lett. 2019, 22, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Krifi, M.N.; Marrakchi, N.; El Ayeb, M.; Dellagi, K. Effect of Some Variables on the In Vivo Determination of Scorpion and Viper Venom Toxicities. Biologicals 1998, 26, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Lyons, K.; Dugon, M.M.; Healy, K. Diet Breadth Mediates the Prey Specificity of Venom Potency in Snakes. Toxins 2020, 12. [Google Scholar] [CrossRef] [Green Version]
- Meier, J.; Theakston, R.D.G. Approximate LD50 determinations of snake venoms using eight to ten experimental animals. Toxicon 1986, 24, 395–401. [Google Scholar] [CrossRef]
- Carrette, T.; Seymour, J. A rapid and repeatable method for venom extraction from cubozoan nematocysts. Toxicon 2004, 44, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.I.; Ryan, N.M.; Page, C.B.; Buckley, N.A.; Brown, S.G.; O’Leary, M.A.; Isbister, G.K. The Australian Snakebite Project, 2005-2015 (ASP-20). Med. J. Aust. 2017, 207, 119–125. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neurotoxins | Coagulants | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Common Name | Species | α- 1 | β- 2 | Other | PFT 3 | CT 4 | MT 5 | Pro- | Anti- |
| Black snake (King Brown/Mulga) | Pseudechis australis | + | + | + | + | ||||
| Coastal taipan | Oxyuranus scutellatus | + | + | + | + | ||||
| Death adder | Acanthophis antarcticus | + | + | + | |||||
| Tiger snake | Notechis scutatus | + | + | + | + | ||||
| Eastern brown snake | Pseudonaja textilis | + | + | + | |||||
| Sea snake | Enhydrina schistosa | + | + | ||||||
| Funnel-web spider (FWS) | Atrax robustus | + | |||||||
| Redback spider (RBS) | Latrodectus hasselti | + | + | ||||||
| Box jellyfish (BJF) | Chironex fleckeri | + | + | + | + | ||||
| Stonefish (SF) | Synanceia sp. | + | + | + | + | + | |||
| Venom | Susceptibility of Chick Embryos in ovo | Venom Dose Causing 100% Mortality (µg/egg) |
|---|---|---|
| Black snake | + | 46 |
| Taipan | + | 4 |
| Death adder | + | 26 |
| Tiger snake | + | 4 |
| Sea snake | + | 20 |
| Funnel-web spider | + | 2 |
| Box jellyfish | + | 37.5 U/egg 1 |
| EEM | Small Animal Assay | ||||
|---|---|---|---|---|---|
| Venom | Venom lot | LD50 (µg/egg) | LD50 (mg/kg) | LD50 (µg/animal) | LD50 (mg/kg) |
| Black snake | PA284 | 0.24 | 0.0043 | 295.4 | 0.739 |
| Brown snake | BR262 | 6.00 | 0.1105 | 6.9 | 0.017 |
| Death Adder | DA416 | 0.89 | 0.0161 | 60.7 | 0.152 |
| Taipan | TP359 | 0.74 | 0.0134 | 5.4 | 0.014 |
| Tiger snake | 286048 | 0.35 | 0.0063 | 6.3 | 0.016 |
| Sea snake | VS0004 | 5.00 | 0.0943 | 23.3 | 0.058 |
| Funnel-web spider | FW123 | 4.47 | 0.0808 | 3.8 | 1.900 |
| Redback spider | BO057/ BO058 | NR 1 | NR 1 | 1.0 sac/mouse | 50.0 sacs/mouse |
| Box jellyfish | BJF-020206 | 37.5 U/egg | 678.1 U/kg | 1.7 U/mouse | 85.0 U/kg |
| Stonefish | 266344 | 5.2 | 0.0956 | 16.7 | 0.835 |
| Antivenom Type | Venom Species | Venom Supplier | Species AV Raised in | Species AV Tested in |
|---|---|---|---|---|
| Black snake | Pseudechis australis | ARP 1 | Horses | Guinea pigs |
| Taipan | Oxyuranus scutellatus | ARP 1 | Horses | Guinea pigs |
| Death Adder | Acanthophis antarcticus | ARP 1 | Horses | Guinea pigs |
| Tiger Snake | Notechis scutatus | ARP 1 | Horses | Guinea pigs |
| Brown Snake | Pseudonaja textilis | ARP 1 | Horses | Guinea pigs |
| Polyvalent Snake | All above | As Above | Horses | Guinea pigs |
| Sea Snake | Enhydrina schistosa | VS 2 | Horses | Guinea pigs |
| Funnel-Web Spider | Atrax robustus | ARP 1 | Rabbits | Suckling mice |
| Redback Spider | Latrodectus hasselti | VS 2 | Horses | Mice |
| Box Jellyfish | Chironex fleckeri | Vision 3 | Sheep | Mice |
| Stonefish | Synanceia horrida, S. verrucosa | JCU 4 | Horses | Mice |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verity, E.E.; Stewart, K.; Vandenberg, K.; Ong, C.; Rockman, S. Potency Testing of Venoms and Antivenoms in Embryonated Eggs: An Ethical Alternative to Animal Testing. Toxins 2021, 13, 233. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040233
Verity EE, Stewart K, Vandenberg K, Ong C, Rockman S. Potency Testing of Venoms and Antivenoms in Embryonated Eggs: An Ethical Alternative to Animal Testing. Toxins. 2021; 13(4):233. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040233
Chicago/Turabian StyleVerity, Erin E., Kathy Stewart, Kirsten Vandenberg, Chi Ong, and Steven Rockman. 2021. "Potency Testing of Venoms and Antivenoms in Embryonated Eggs: An Ethical Alternative to Animal Testing" Toxins 13, no. 4: 233. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040233