Cyanotoxin Screening in BACA Culture Collection: Identification of New Cylindrospermopsin Producing Cyanobacteria

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Detection of Cyanotoxin(s)-Producing Cyanobacteria
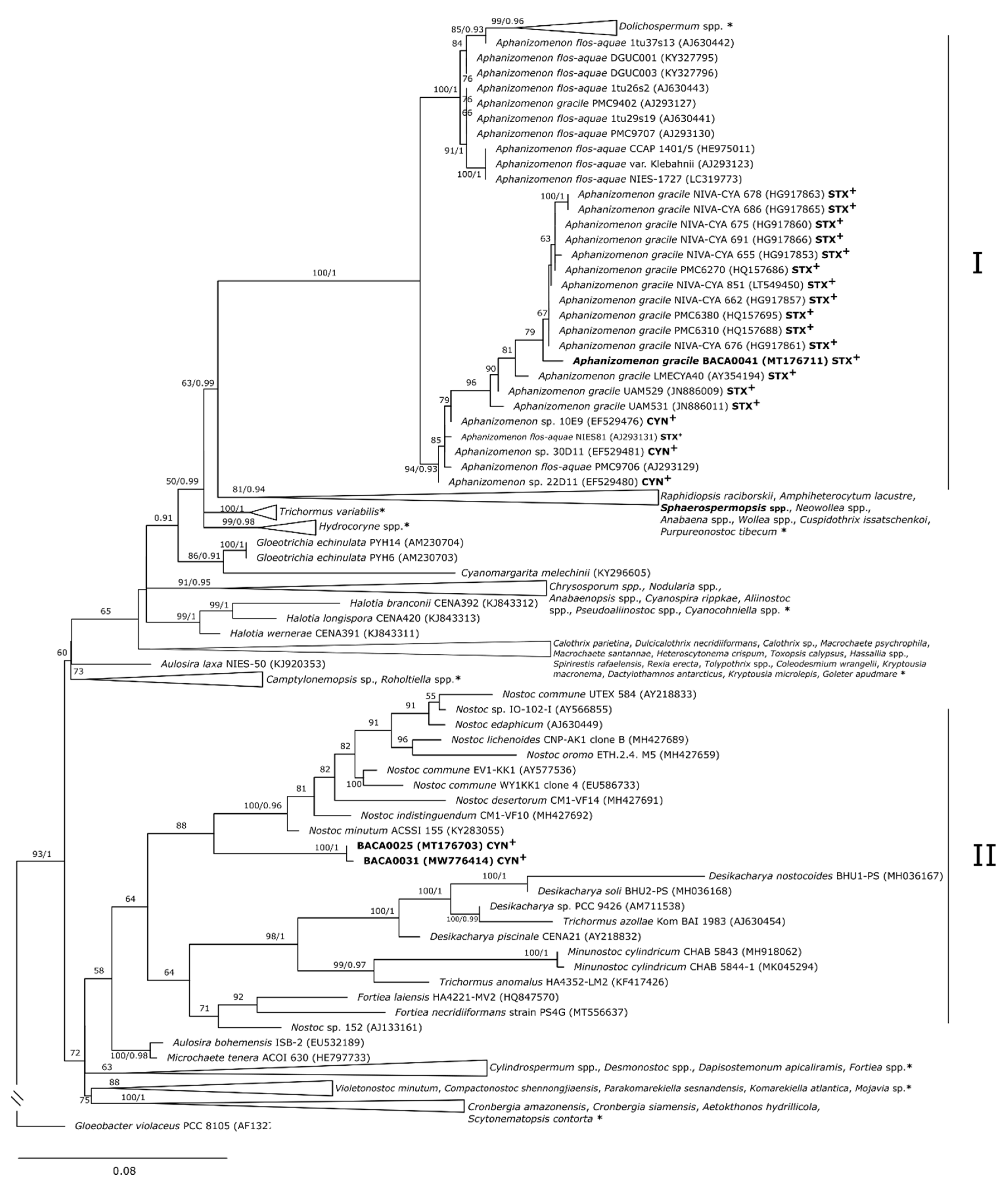
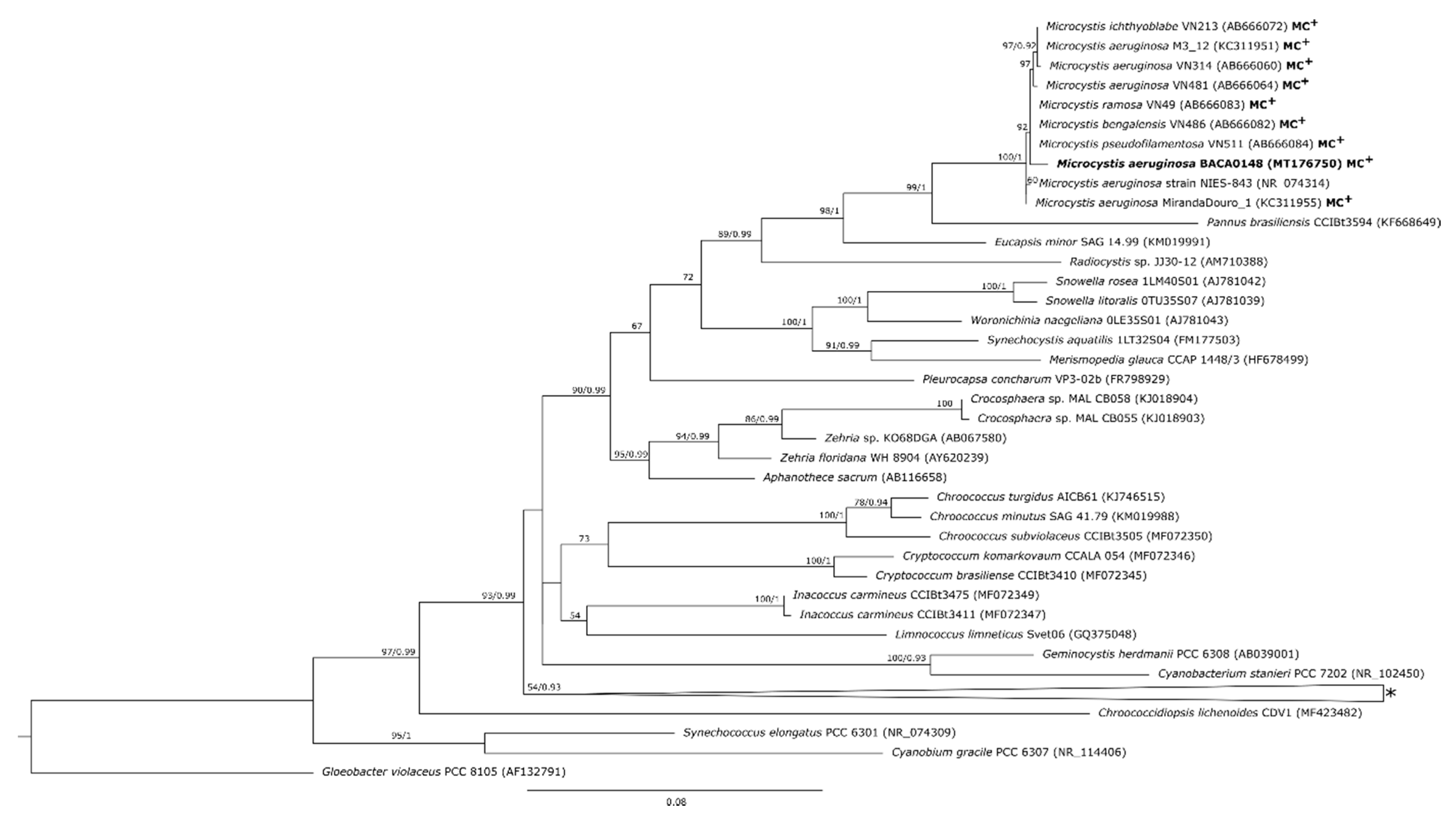
2.2. Phylogenetic Characterization
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. BACA Strains and Growth Conditions
5.2. Cyanotoxins Analysis
5.2.1. Toxin extraction
5.2.2. ESI-LC-MS/MS analysis
5.3. DNA Extraction, PCR Amplification, and Sequencing
5.4. Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamilton, D.P.; Salmaso, N.; Paerl, H.W. Mitigating harmful cyanobacterial blooms: Strategies for control of nitrogen and phosphorus loads. Aquat. Ecol. 2016, 50, 351–366. [Google Scholar] [CrossRef]
- Brooks, B.W.; Lazorchak, J.M.; Howard, M.D.A.; Johnson, M.-V.V.; Morton, S.L.; Perkins, D.A.K.; Reavie, E.D.; Scott, G.I.; Smith, S.A.; Steevens, J.A. Are harmful algal blooms becoming the greatest inland water quality threat to public health and aquatic ecosystems? Environ. Toxicol. Chem. 2016, 35, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Holland, A.; Kinnear, S. Interpreting the Possible Ecological Role(s) of Cyanotoxins: Compounds for Competitive Advantage and/or Physiological Aide? Mar. Drugs 2013, 11, 2239–2258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water, 2nd ed.; CRC Press, on behalf of the World Health Organization: Boca Raton, CH, Geneva, 2021. [Google Scholar]
- Van Apeldoorn, M.E.; Van Egmond, H.P.; Speijers, G.J.A.; Bakker, G.J.I. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Fewer, D.P.; Neilan, B. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Cirés, S.; Casero, M.C.; Quesada, A. Toxicity at the edge of life: A review on cyanobacterial toxins from extreme environments. Mar. Drugs 2017, 15, 233. [Google Scholar] [CrossRef] [PubMed]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.-F. A review of current knowledge on toxic benthic freshwater cyanobacteria–Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef]
- Rantala-Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a synthetase gene cluster of the cyanobacterium Anabaena sp. strain 37 and molecular methods to detect potential producers. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledreux, A.; Thomazeau, S.; Catherine, A.; Duval, C.; Yéprémian, C.; Marie, A.; Bernard, C. Evidence for saxitoxins production by the cyanobacterium Aphanizomenon gracile in a French recreational water body. Harmful Algae 2010, 10, 88–97. [Google Scholar] [CrossRef]
- Codd, G.A.; Meriluoto, J.; Metcalf, J.S. Introduction: Cyanobacteria, Cyanotoxins, Their Human Impact, and Risk Management. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Codd, G.A., Meriluoto, J., Metcalf, J.S., Eds.; John Wiley and Sons, Ltd: Chichester, UK, 2017; pp. 1–8. [Google Scholar]
- Bernard, C.; Ballot, A.; Thomazeau, S.; Maloufi, S.; Furey, A.; Mankiewicz-Boczek, J.; Pawlik-Skowrońska, B.; Capelli, C.; Salmaso, N. Appendix 2: Cyanobacteria associated with the production of cyanotoxins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley and Sons, Ltd: Chichester, UK, 2017. [Google Scholar]
- Tillett, D.; Dittmann, E.; Erhard, M.; Von Döhren, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Heck, K.; Alvarenga, D.O.; Shishido, T.K.; Varani, A.M.; Dörr, F.A.; Pinto, E.; Rouhiainen, L.; Jokela, J.; Sivonen, K.; Fiore, M.F. Biosynthesis of microcystin hepatotoxins in the cyanobacterial genus Fischerella. Toxicon 2018, 141, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bláha, L.; Babica, P.; Maršálek, B. Toxins produced in cyanobacterial water blooms–toxicity and risks. Interdiscip. Toxicol. 2009, 2, 10102–10109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metcalf, J.S.; Codd, G.A. Cyanotoxins. In Ecology of Cyanobacteria II; Whitton, B., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 651–675. [Google Scholar]
- Oksanen, I.; Jokela, J.; Fewer, D.P.; Wahlsten, M.; Rikkinen, J.; Sivonen, K. Discovery of rare and highly toxic microcystins from lichen-associated cyanobacterium Nostoc sp. Strain IO-102-I. Appl. Environ. Microbiol. 2004, 70, 5756–5763. [Google Scholar] [CrossRef] [Green Version]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B. a Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The genetics, biosynthesis and regulation of toxic specialized metabolites of cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Borges, H.L.F.; Branco, L.H.Z.; Martins, M.D.; Lima, C.S.; Barbosa, P.T.; Lira, G.A.S.; Bittencourt-Oliveira, M.C.; Molica, R.J.R. Cyanotoxin production and phylogeny of benthic cyanobacterial strains isolated from the northeast of Brazil. Harmful Algae 2015, 43, 46–57. [Google Scholar] [CrossRef]
- Humpage, A.; Rositano, J.; Bretag, A.; Brown, R.; Baker, P.; Nicholson, B.; Steffensen, D. Paralytic shellfish poisons from Australian cyanobacterial blooms. Mar. Freshw. Res. 1994, 45, 761. [Google Scholar] [CrossRef]
- Sivonen, K.; Himberg, K.; Luukkainen, R.; Niemelä, S.I.; Poon, G.K.; Codd, G.A. Preliminary characterization of neurotoxic cyanobacteria blooms and strains from Finland. Toxic. Assess. 1989, 4, 339–352. [Google Scholar] [CrossRef]
- Kellmann, R.; Mihali, T.K.; Jeon, Y.J.; Pickford, R.; Pomati, F.; Neilan, B. a Biosynthetic Intermediate Analysis and Functional Homology Reveal a Saxitoxin Gene Cluster in Cyanobacteria. Appl. Environ. Microbiol. 2008, 74, 4044–4053. [Google Scholar] [CrossRef] [Green Version]
- Mihali, T.K.; Kellmann, R.; Neilan, B. a Characterisation of the paralytic shellfish toxin biosynthesis gene clusters in Anabaena circinalis AWQC131C and Aphanizomenon sp. NH-5. BMC Biochem. 2009, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Casero, M.C.; Ballot, A.; Agha, R.; Quesada, A.; Cirés, S. Characterization of saxitoxin production and release and phylogeny of sxt genes in paralytic shellfish poisoning toxin-producing Aphanizomenon Gracile. Harmful Algae 2014, 37, 28–37. [Google Scholar] [CrossRef]
- Hawkins, P.R.; Runnegar, M.T.C.; Jackson, A.R.B.; Falconer, I.R. Severe hepatotoxicity caused by the tropical cyanobacterium (blue-green alga) Cylindrospermopsis raciborskii (Woloszynska) Seenaya and Subba Raju isolated from a domestic water supply reservoir. Appl. Environ. Microbiol. 1985, 50, 1292–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokociński, M.; Mankiewicz-Boczek, J.; Jurczak, T.; Spoof, L.; Meriluoto, J.; Rejmonczyk, E.; Hautala, H.; Vehniäinen, M.; Pawełczyk, J.; Soininen, J. Aphanizomenon gracile (Nostocales), a cylindrospermopsin-producing cyanobacterium in Polish lakes. Environ. Sci. Pollut. Res. 2013, 20, 5243–5264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banker, R.; Carmeli, S.; Hadas, O.; Teltsch, B.; Porat, R.; Sukenik, A. Identification of cylindrospermopsin in Aphanizomenon ovalisporum (Cyanophyceae) isolated from lake Kinneret, Israel. J. Phycol. 1997, 33, 613–616. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef] [Green Version]
- Whitton, B.; Potts, M. Introduction to the Cyanobacteria. In The Ecology of Cyanobacteria II–Their Diversity in Time and Space; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 1–13. ISBN 978-94-007-3854-6. [Google Scholar]
- Komárek, J.; Kastovsky, J.; Mares, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Mai, T.; Johansen, J.R.; Pietrasiak, N.; Bohunická, M.; Martin, M.P. Revision of the Synechococcales (Cyanobacteria) through recognition of four families including Oculatellaceae fam. nov. and Trichocoleaceae fam. nov. and six new genera containing 14 species. Phytotaxa 2018, 365, 1–59. [Google Scholar] [CrossRef] [Green Version]
- Bagchi, S.N.; Dubey, N.; Singh, P. Phylogenetically distant clade of Nostoc-like taxa with the description of Aliinostoc gen. nov. and Aliinostoc morphoplasticum sp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 3329–3338. [Google Scholar] [CrossRef]
- Genuário, D.B.; Vaz, M.G.M.V.; Hentschke, G.S.; Sant’Anna, C.L.; Fiore, M.F. Halotia gen. nov., a phylogenetically and physiologically coherent cyanobacterial genus isolated from marine coastal environments. Int. J. Syst. Evol. Microbiol. 2015, 65, 663–675. [Google Scholar] [CrossRef] [Green Version]
- Brito, Â.; Ramos, V.; Mota, R.; Lima, S.; Santos, A.; Vieira, J.; Vieira, C.P.; Kaštovský, J.; Vasconcelos, V.M.; Tamagnini, P. Description of new genera and species of marine cyanobacteria from the Portuguese Atlantic coast. Mol. Phylogenet. Evol. 2017, 111, 18–34. [Google Scholar] [CrossRef]
- Cai, F.; Li, X.; Yang, Y.; Jia, N.; Huo, D.; Li, R. Compactonostoc shennongjiaensis gen. & sp. nov. (Nostocales, Cyanobacteria) from a wet rocky wall in China. Phycologia 2019, 58, 200–210. [Google Scholar] [CrossRef]
- Österholm, J.; Popin, R.V.; Fewer, D.P.; Sivonen, K. Phylogenomic Analysis of Secondary Metabolism in the Toxic Cyanobacterial Genera Anabaena, Dolichospermum and Aphanizomenon. Toxins 2020, 12, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humbert, J.-F.; Fastner, J. Ecology of Cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley and Sons, Ltd: Chichester, UK, 2017; pp. 9–18. [Google Scholar]
- Cordeiro, R.; Luz, R.; Vasconcelos, V.; Fonseca, A.; Gonçalves, V. A Critical Review of Cyanobacteria Distribution and Cyanotoxins Occurrence in Atlantic Ocean Islands. Cryptogam. Algol. 2020, 41, 73–89. [Google Scholar] [CrossRef]
- Moreira, C.; Ramos, V.; Azevedo, J.; Vasconcelos, V. Methods to detect cyanobacteria and their toxins in the environment. Appl. Microbiol. Biotechnol. 2014, 98, 8073–8082. [Google Scholar] [CrossRef] [PubMed]
- Sanseverino, I.; António, D.C.; Loos, R.; Lettieri, T. Cyanotoxins: Methods and Approaches for Their Analysis and Detection; Publications Office of the European Union: Luxembourg, 2017. [Google Scholar]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Ballot, A.; Cerasino, L.; Hostyeva, V.; Cirés, S. Variability in the sxt Gene Clusters of PSP Toxin Producing Aphanizomenon gracile Strains from Norway, Spain, Germany and North America. PLoS ONE 2016, 11, 0167552. [Google Scholar] [CrossRef]
- Murray, S.A.; Mihali, T.K.; Neilan, B.A. Extraordinary Conservation, Gene Loss, and Positive Selection in the Evolution of an Ancient Neurotoxin. Mol. Biol. Evol. 2011, 28, 1173–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustafa, A.; Loram, J.E.; Hackett, J.D.; Anderson, D.M.; Plumley, F.G.; Bhattacharya, D. Origin of saxitoxin biosynthetic genes in cyanobacteria. PLoS ONE 2009, 4, 5758. [Google Scholar] [CrossRef] [Green Version]
- Ballot, A.; Fastner, J.; Wiedner, C. Paralytic shellfish poisoning toxin-producing cyanobacterium Aphanizomenon gracile in Northeast Germany. Appl. Environ. Microbiol. 2010, 76, 1173–1180. [Google Scholar] [CrossRef] [Green Version]
- Cirés, S.; Wörmer, L.; Ballot, A.; Agha, R.; Wiedner, C.; Velázquez, D.; Casero, M.C.; Quesada, A. Phylogeography of cylindrospermopsin and paralytic shellfish toxin-producing nostocales cyanobacteria from mediterranean europe (Spain). Appl. Environ. Microbiol. 2014, 80, 1359–1370. [Google Scholar] [CrossRef] [Green Version]
- Akcaalan, R.; Köker, L.; Oğuz, A.; Spoof, L.; Meriluoto, J.; Albay, M. First report of cylindrospermopsin production by two cyanobacteria (Dolichospermum mendotae and Chrysosporum ovalisporum) in Lake Iznik, Turkey. Toxins 2014, 6, 3173–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballot, A.; Ramm, J.; Rundberget, T.; Kaplan-Levy, R.N.; Hadas, O.; Sukenik, A.; Wiedner, C. Occurrence of non-cylindrospermopsin-producing Aphanizomenon ovalisporum and Anabaena bergii in Lake Kinneret (Israel). J. Plankton Res. 2011, 33, 1736–1746. [Google Scholar] [CrossRef]
- Gantar, M.; Sekar, R.; Richardson, L.L. Cyanotoxins from black band disease of corals and from other coral reef environments. Microb. Ecol. 2009, 58, 856–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.D.; Guo, H.; Zhang, H. The diversity of cyanobacterial toxins on structural characterization, distribution and identification: A systematic review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayumi, T.; Kato, H.; Imanishi, S.; Kawasaki, Y.; Hasegawa, M.; Harada, K. Structural Characterization of Microcystins by LC/MS/MS under Ion Trap Conditions. J. Antibiot. 2006, 59, 710–719. [Google Scholar] [CrossRef] [Green Version]
- Rantala, A.; Fewer, D.P.; Hisbergues, M.; Rouhiainen, L.; Vaitomaa, J.; Börner, T.; Sivonen, K. Phylogenetic evidence for the early evolution of microcystin synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 568–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meriluoto, J.; Spoof, L.; Codd, G.A. Appendix 3: Tables of Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley and Sons, Ltd: Chichester, UK, 2017; pp. 526–537. [Google Scholar]
- Galetović, A.; Azevedo, J.; Castelo-Branco, R.; Oliveira, F.; Gómez-Silva, B.; Vasconcelos, V. Absence of Cyanotoxins in Llayta, Edible Nostocaceae Colonies from the Andes Highlands. Toxins 2020, 12, 382. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R. Isolation and purification of cyanobacteria. In Methods in Enzymology; Elsvier: Amsterdam, The Netherlands, 1988; Volume 167, pp. 3–27. [Google Scholar]
- Rippka, R.; Waterbury, J.; Stanier, R. Isolation and Purification of Cyanobacteria: Some General Principles. In The Prokaryotes: A Handbook on Habitats, Isolation and Identification of Bacteria.; Starr, M.P., Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: Berlin/Heidelberg, Germany, 1981. [Google Scholar]
- Cordeiro, R.; Luz, R.; Vasconcelos, V.; Gonçalves, V.; Fonseca, A. Cyanobacteria Phylogenetic Studies Reveal Evidence for Polyphyletic Genera from Thermal and Freshwater Habitats. Diversity 2020, 12, 298. [Google Scholar] [CrossRef]
- Neilan, B.A.; Jacobs, D.; Del Dot, T.; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman, A.E. rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol. 1997, 47, 693–697. [Google Scholar] [CrossRef]
- Nübel, U.; Garcia-Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef] [Green Version]
- Ouahid, Y.; Pérez-Silva, G.; Del Campo, F.F. Identification of potentially toxic environmental Microcystis by individual and multiple PCR amplification of specific microcystin synthetase gene regions. Environ. Toxicol. 2005, 20, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Schembri, M.A.; Neilan, B.A.; Saint, C.P. Identification of genes implicated in toxin production in the cyanobacterium Cylindrospermosis raciborskii. Environ. Toxicol. 2001, 16, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Foss, A.J.; Selwood, A.I.; Özen, M.; Boundy, M. Paralytic shellfish toxin producing Aphanizomenon gracile strains isolated from Lake Iznik, Turkey. Toxicon 2018, 148, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Ballot, A.; Swe, T.; Mjelde, M.; Cerasino, L.; Hostyeva, V.; Miles, C.O. Cylindrospermopsin- and Deoxycylindrospermopsin-Producing Raphidiopsis raciborskii and Microcystin-Producing Microcystis spp. in Meiktila Lake, Myanmar. Toxins 2020, 12, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuken, A.; Campbell, R.J.; Quesada, A.; Sukenik, A.; Dadheech, P.K.; Wiedner, C. Genetic and morphologic characterization of four putative cylindrospermopsin producing species of the cyanobacterial genera Anabaena and Aphanizomenon. J. Plankton Res. 2009, 31, 465–480. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, W.; Li, D.; Shen, Y.; Liu, Y.; Song, L. Analysis of paralytic shellfish toxins in Aphanizomenon DC-1 from Lake Dianchi, China. Environ. Toxicol. 2006, 21, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Lyra, C.; Suomalainen, S.; Gugger, M.; Vezie, C.; Sundman, P.; Paulin, L.; Sivonen, K. Molecular characterization of planktic cyanobacteria of Anabaena, Aphanizomenon, Microcystis and Planktothrix genera. Int. J. Syst. Evol. Microbiol. 2001, 51, 513–526. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, 232–235. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Strains | Habitat | Species | PCR | ESI-LC-MS/MS | ||||
|---|---|---|---|---|---|---|---|---|
| SXT | CYN | MC | SXT | CYN | MC-LR | |||
| BACA0007 | Lake | Kamptonema sp. | G | - | - | - | - | - |
| BACA0025 | Lake | unidentified species | - | B, CC | - | - | + | - |
| BACA0031 | Lake | unidentified species | - | B, CC | - | - | + | - |
| BACA0041 | Lake | Aphanizomenon gracile | A, G, H, I | - | - | + | - | - |
| BACA0081 | Lake | Cylindrospermum sp. | H | - | - | - | - | - |
| BACA0091 | Lake | Nostoc sp. | - | - | E | - | - | - |
| BACA0109 | Lake | Nostoc sp. | - | B | - | - | - | - |
| BACA0142 | Thermal | Leptolyngbya sp. | - | B | - | - | - | - |
| BACA0146 | Thermal | Leptolyngbya sp. | - | B | - | - | - | - |
| BACA0148 | Lake | Microcystis aeruginosa | - | - | C, D, E, G | - | - | + |
| BACA0203 | Lake | Leptodesmis sp. | G, H | - | - | - | - | - |
| BACA0204 | Lake | Leptolyngbya sp. | G | - | - | - | - | - |
| BACA0223 | Lake | Anathece minutissima | G | - | - | - | - | - |
| Gene | Primer | Fragment length (bp) | Sequence (5’–3’) | References |
|---|---|---|---|---|
| mcyC | PSCF1 | 674 | GCAACATCCCAAGAGCAAAG | Ouahid et al. [61] |
| PSCR1 | CCGACAACATCACAAAGGC | |||
| mcyD | PKDF1 | 647 | GACGCTCAAATGATGAAAC | |
| PKDR1 | GCAACCGATAAAAACTCCC | |||
| mcyE | PKEF1 | 755 | CGCAAACCCGATTTACAG | |
| PKER1 | CCCCTACCATCTTCATCTTC | |||
| mcyG | PKGF1 | 425 | ACTCTCAAGTTATCCTCCCTC | |
| PKGR1 | AATCGCTAAAACGCCACC | |||
| sxtA | sxtA F | 602 | AGGTCTTTGACTTGCATCCAA | Ledreux et al. [10] |
| sxtA R | AACCGGCGACATAGATGATA | |||
| sxtG | sxtGf | 893 | AGGAATTCCCTATCCACCGGAG | Casero et al. [25] |
| sxtGr | CGGCGAACATCTAACGTTGCAC | |||
| sxtH | sxtHf | 812 | AAGACCACTGTCCCCACCGAGG | |
| sxtHr | CTGTGCAGCGATCTGATGGCAC | |||
| sxtI | sxtIf | 910 | AGCGCTGCCGCTATGGTTGTCG | |
| sxtIr | ACGCAATTGAGGGCGACACCAC | |||
| cyrB | M13 | 597 | GGCAAATTGTGATAGCCACGAGC | Schembri et al. [62] |
| M14 | GATGGAACATCGCTCACTGGTG | |||
| cyrC | M4 | 650 | GAAGCTCTGGAATCCGGTAA | |
| M5 | AATCCTTACGGGATCCGGTGC | |||
| 16S rRNA | 27F | AGAGTTTGATCCTGGCTCAG | Neilan et al. [59] | |
| CYA359F | GGGGAATYTTCCGCAATG GG | Nubel et al. [60] | ||
| 1494R | TACGGCTACCTTGTTACGAC | Neilan et al. [59] | ||
| CYA781R | GACTACTGGGGTATCTAATCCCATT | Nubel et al. [60] | ||
| CYA781F | AATGGGATTAGATACCCCAGTAGTC | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordeiro, R.; Azevedo, J.; Luz, R.; Vasconcelos, V.; Gonçalves, V.; Fonseca, A. Cyanotoxin Screening in BACA Culture Collection: Identification of New Cylindrospermopsin Producing Cyanobacteria. Toxins 2021, 13, 258. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040258
Cordeiro R, Azevedo J, Luz R, Vasconcelos V, Gonçalves V, Fonseca A. Cyanotoxin Screening in BACA Culture Collection: Identification of New Cylindrospermopsin Producing Cyanobacteria. Toxins. 2021; 13(4):258. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040258
Chicago/Turabian StyleCordeiro, Rita, Joana Azevedo, Rúben Luz, Vitor Vasconcelos, Vítor Gonçalves, and Amélia Fonseca. 2021. "Cyanotoxin Screening in BACA Culture Collection: Identification of New Cylindrospermopsin Producing Cyanobacteria" Toxins 13, no. 4: 258. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040258