Recombinant Expression of Trametes versicolor Aflatoxin B1-Degrading Enzyme (TV-AFB1D) in Engineering Pichia pastoris GS115 and Application in AFB1 Degradation in AFB1-Contaminated Peanuts

Abstract
:1. Introduction
2. Results
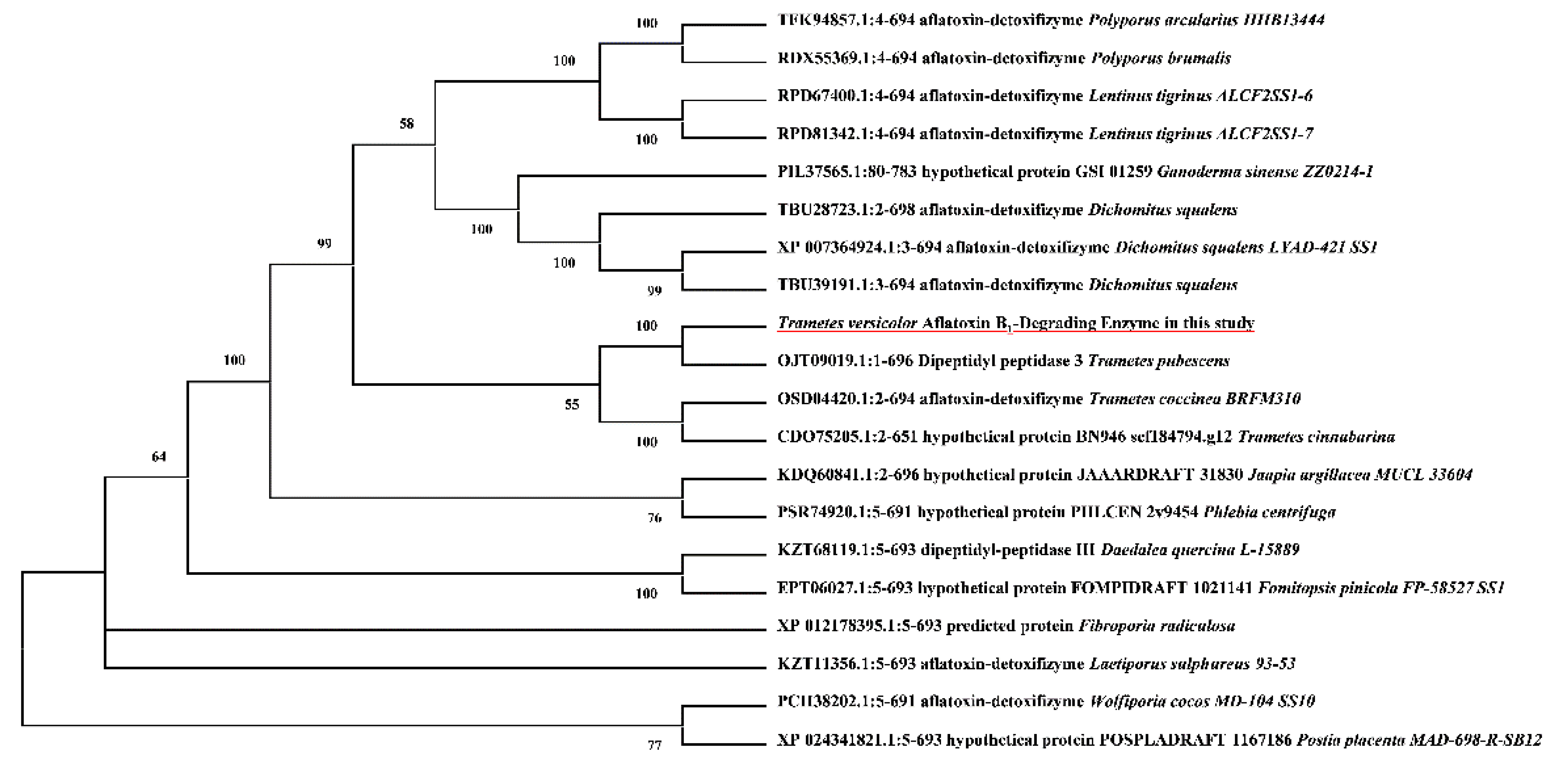
2.1. TV-AFB1D Cloning and Evolutionary Tree Analysis
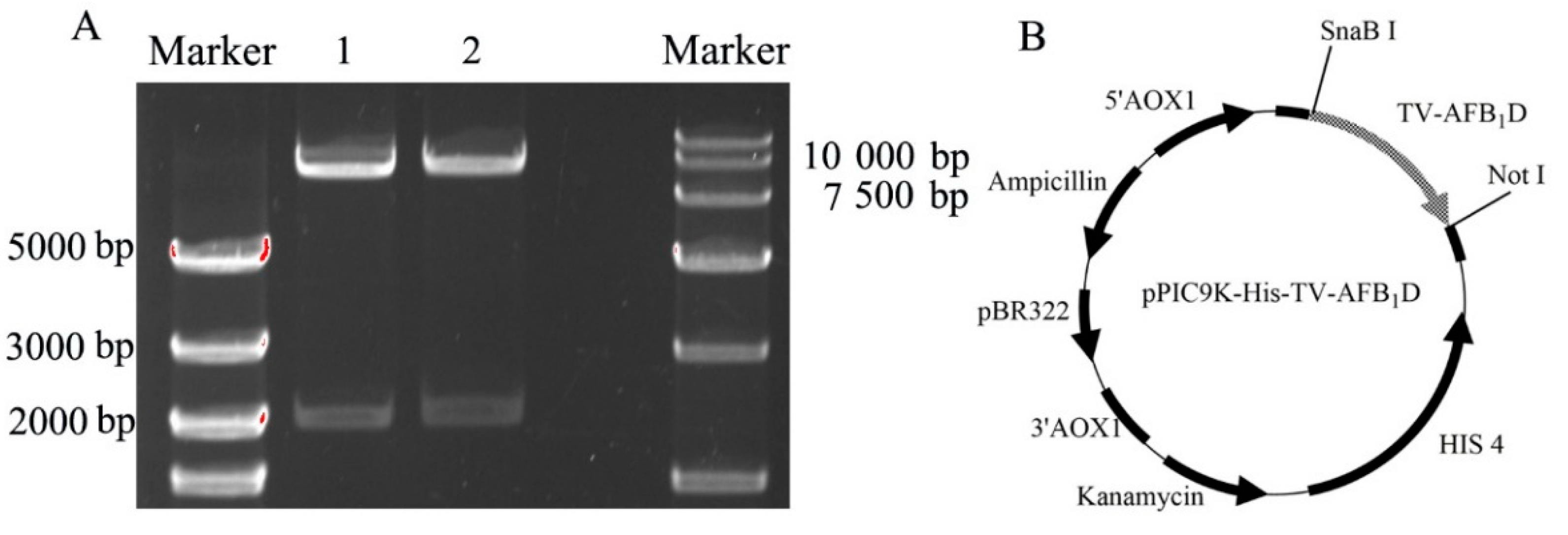
2.2. Expression Vector Construction of TV-AFB1D
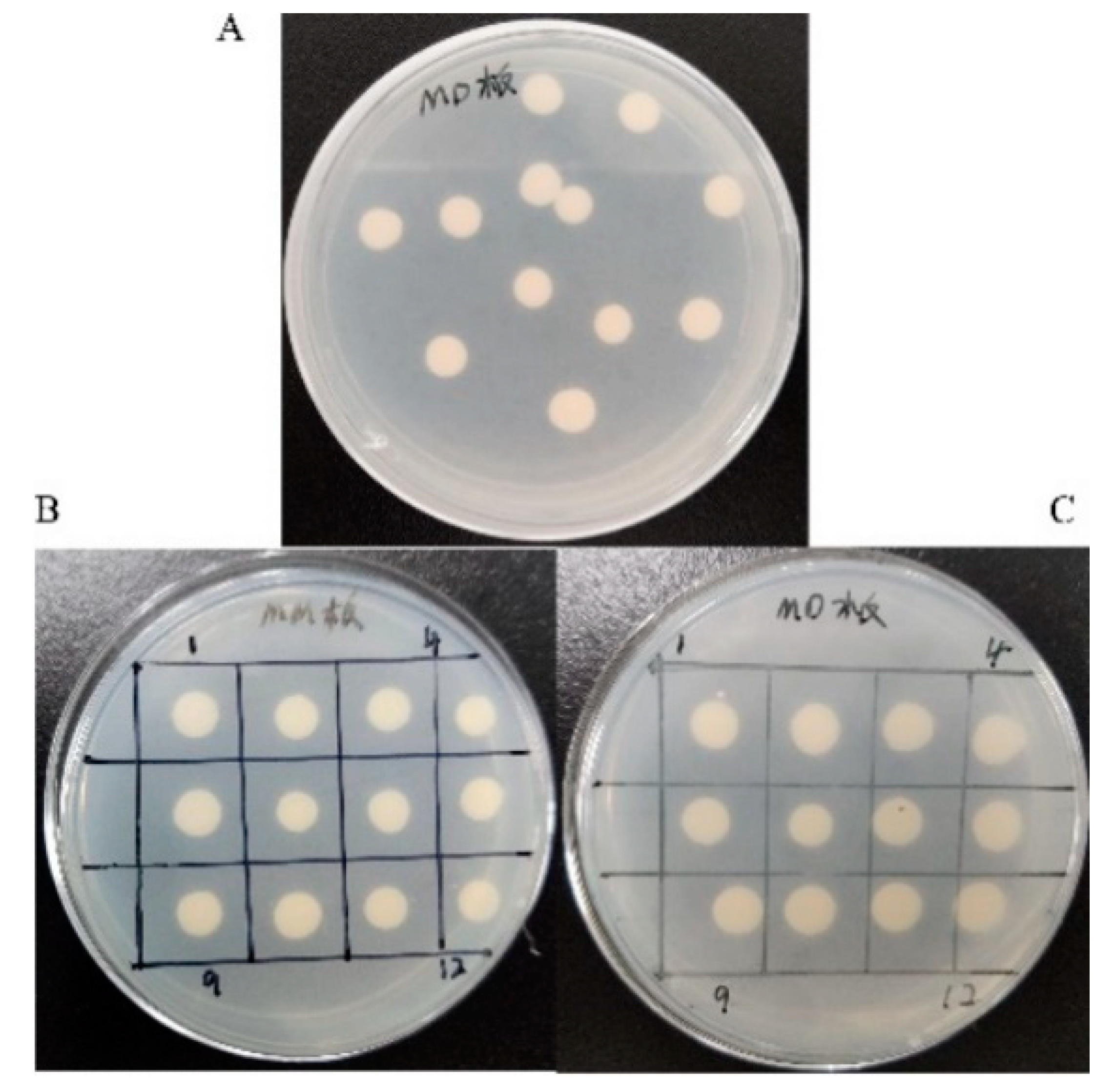
2.3. Transformation of TV-AFB1D into P. pastoris GS115
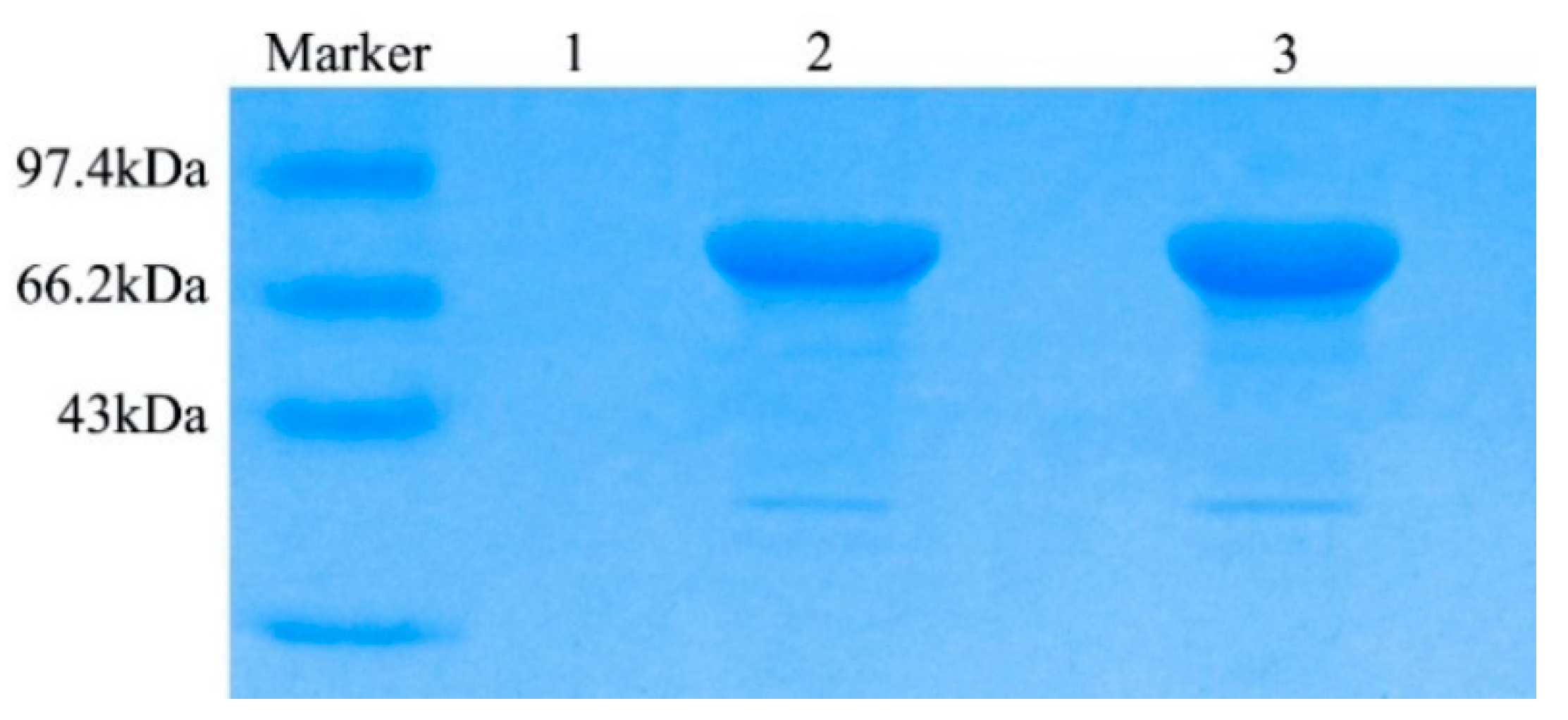
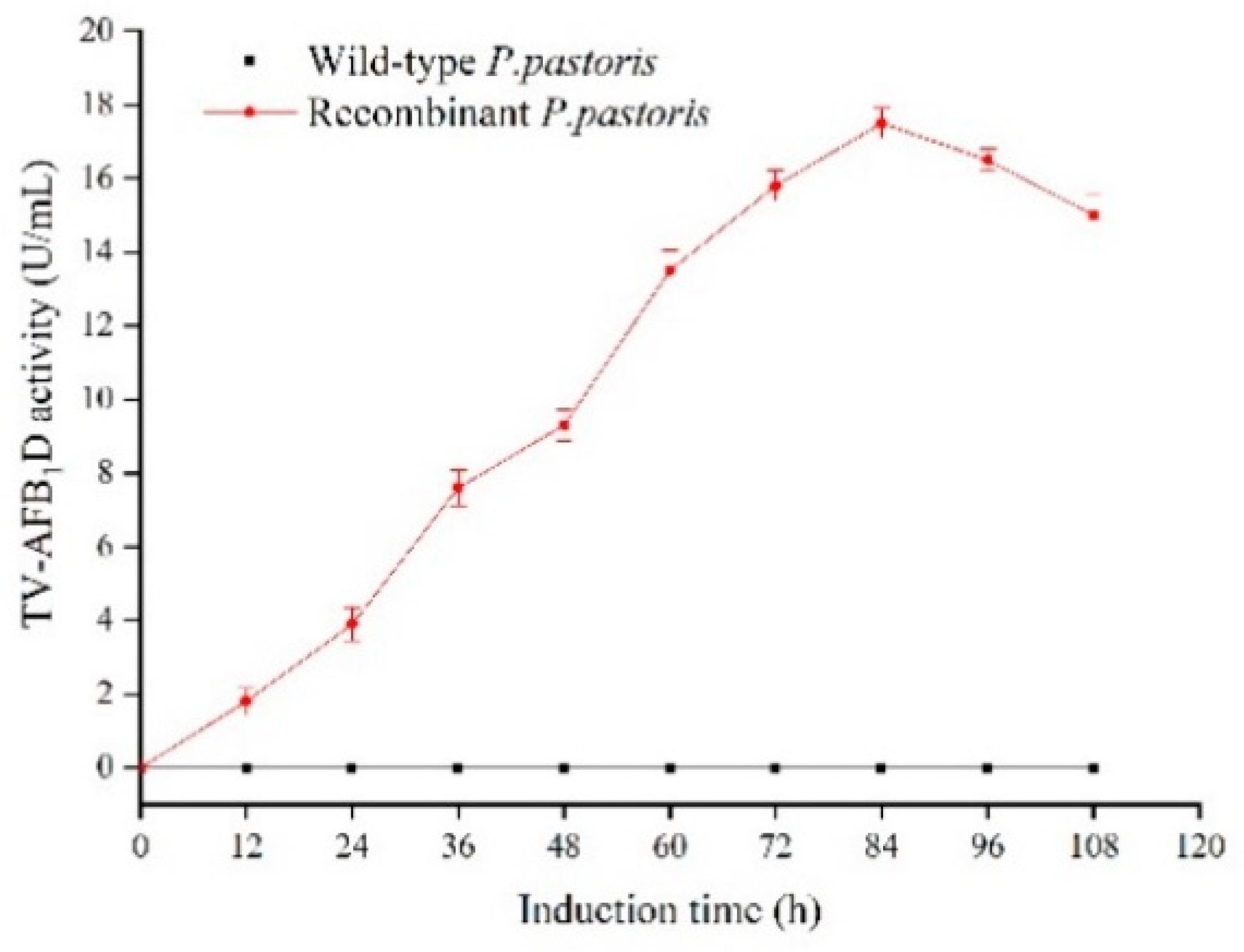
2.4. Recombinant TV-AFB1D Expression in P. pastoris GS115
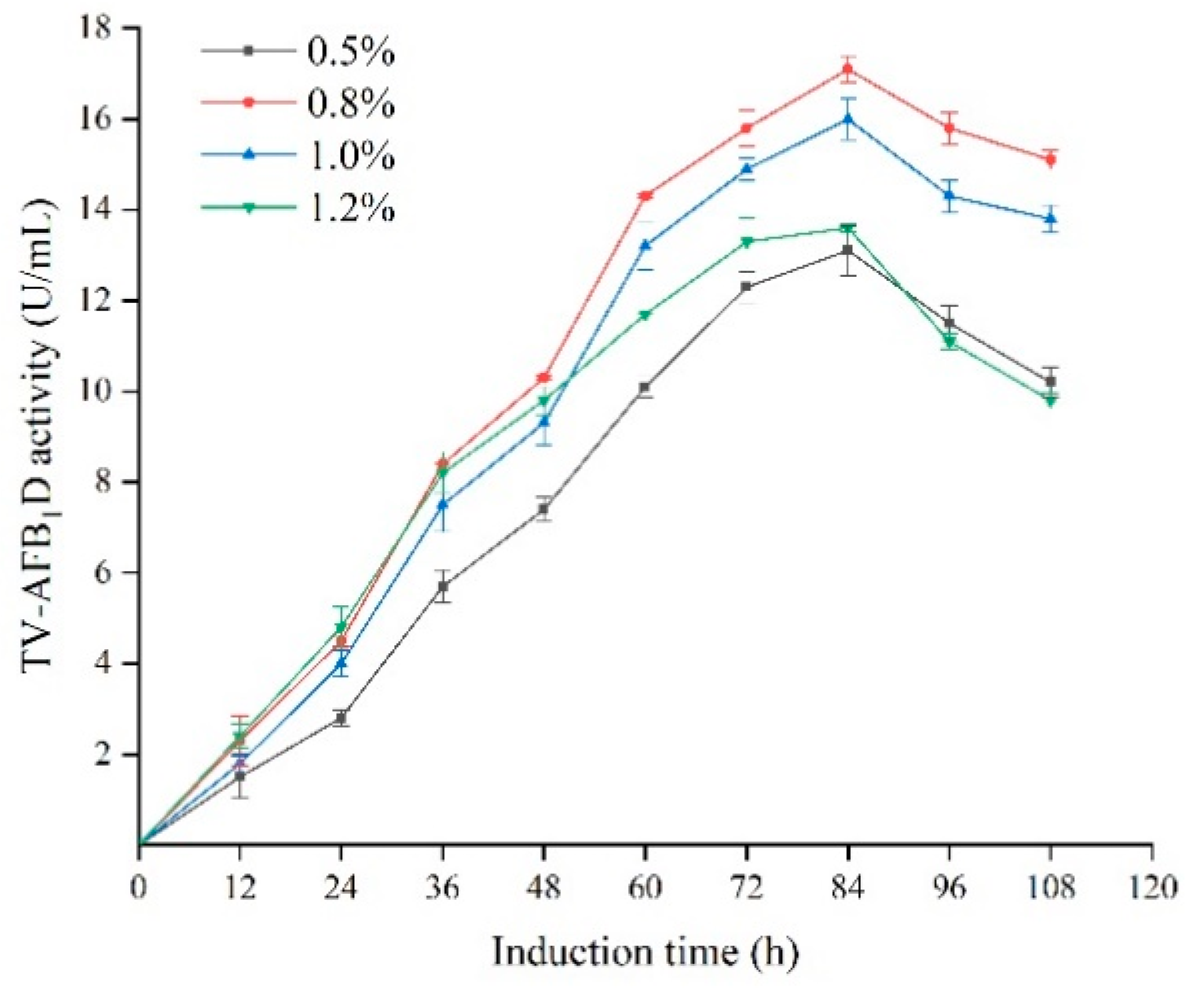
2.5. Effect of Methanol Concentration and Time on the TV-AFB1D Expression
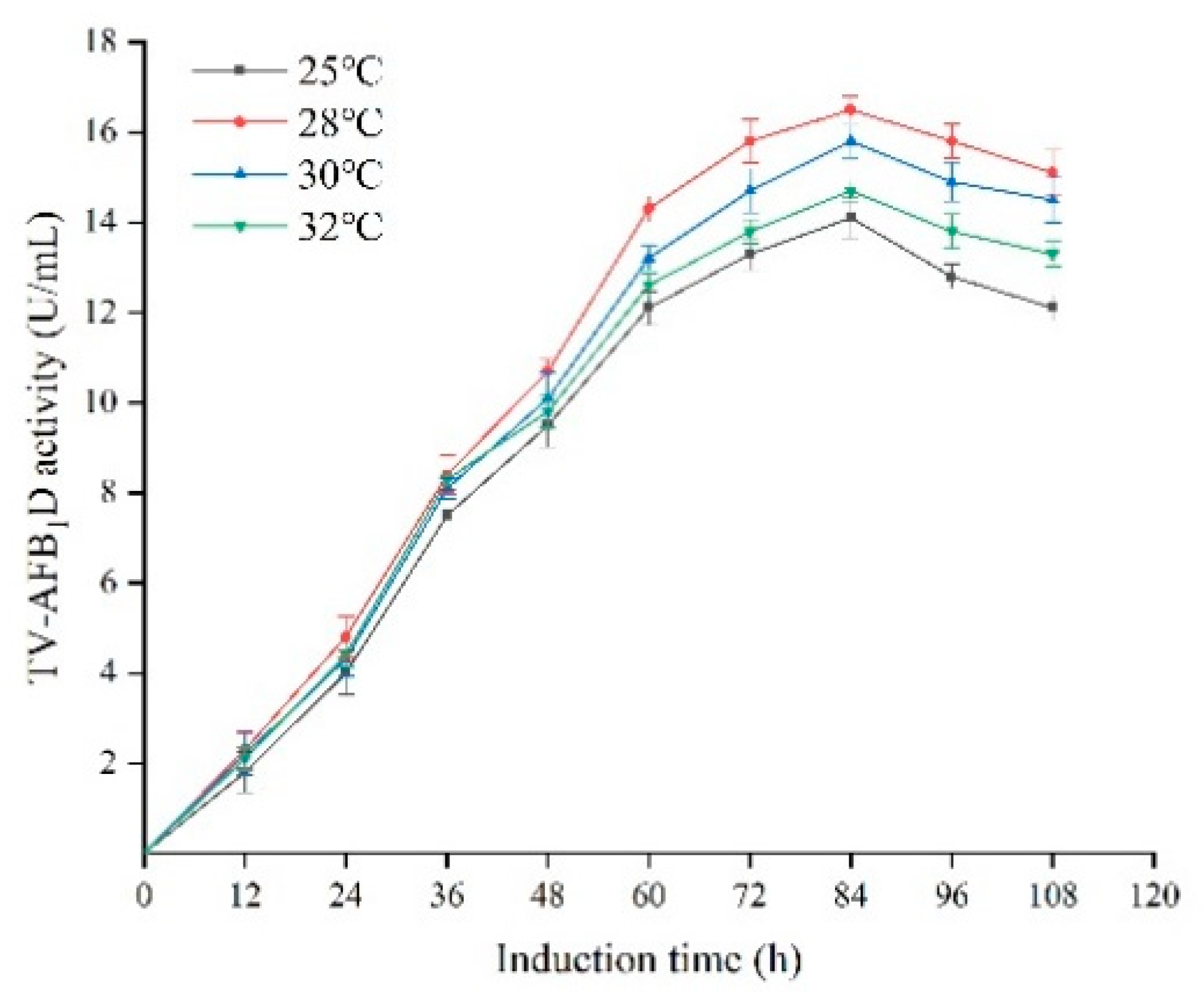
2.6. Effect of Temperatures and Treatment Time on the Expression of TV-AFB1D
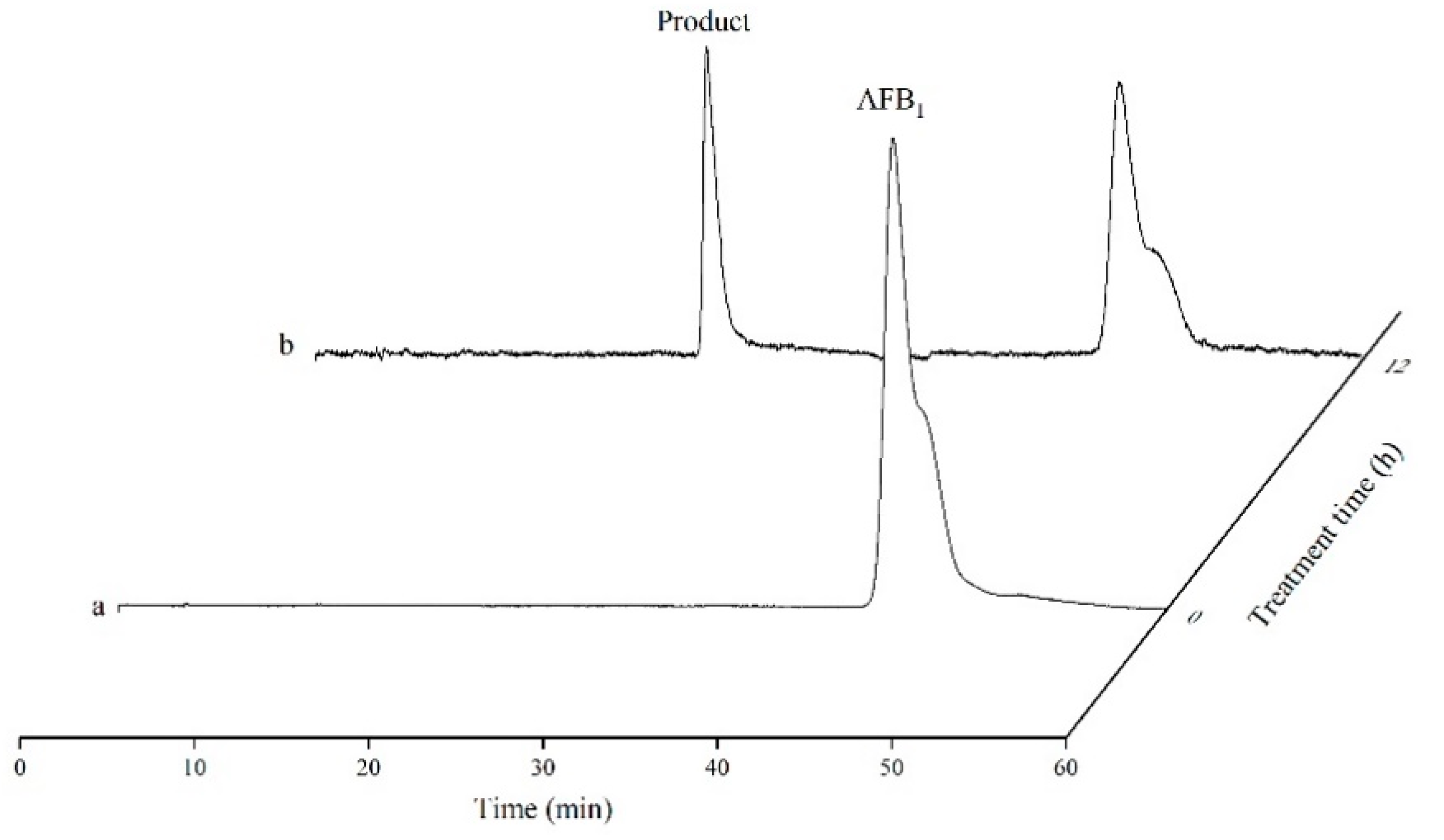
2.7. Product Formation of AFB1-Contaminated Peanuts Catalyzed by TV-AFB1D
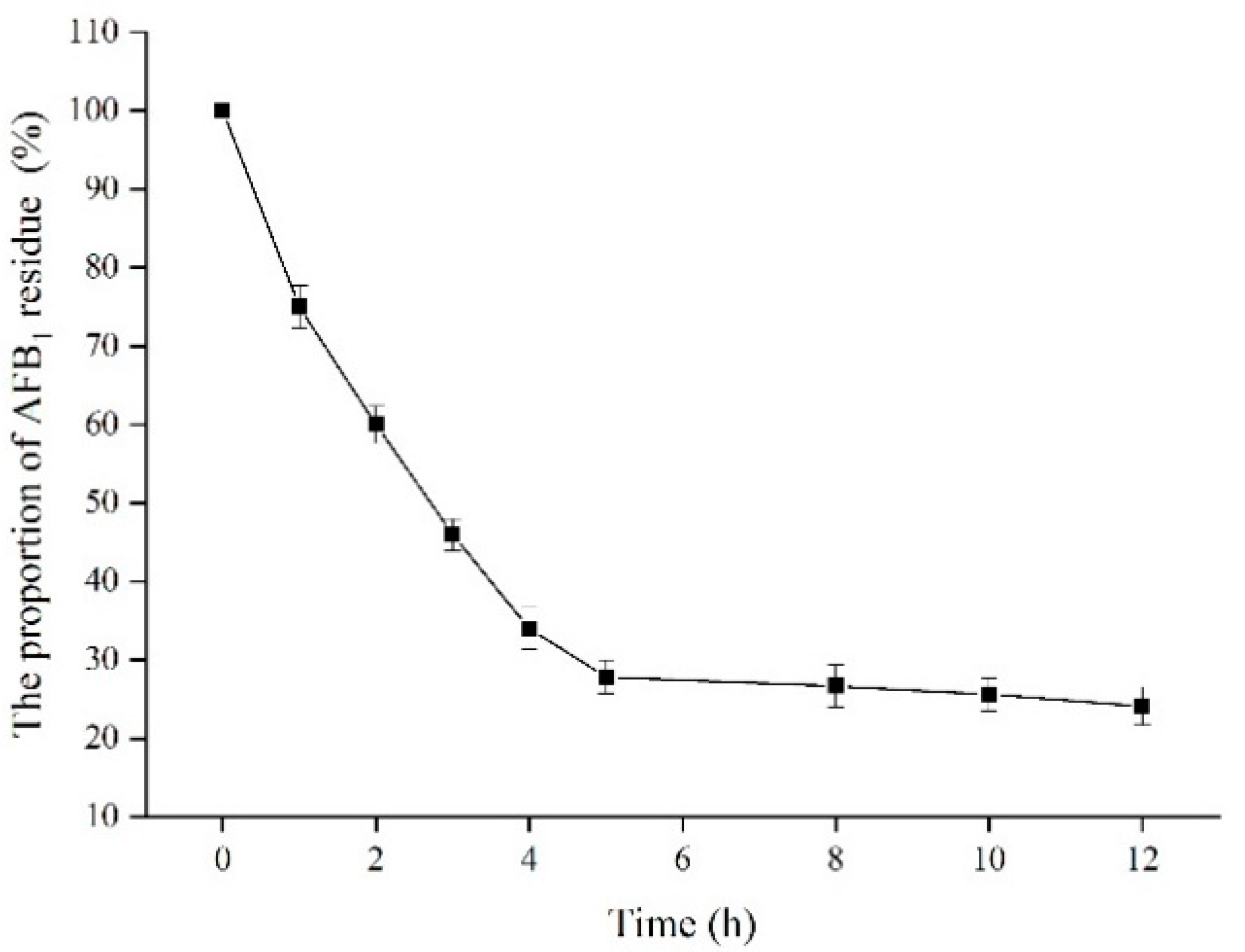
2.8. Effect of Time on the Residue of AFB1 Standard Sample
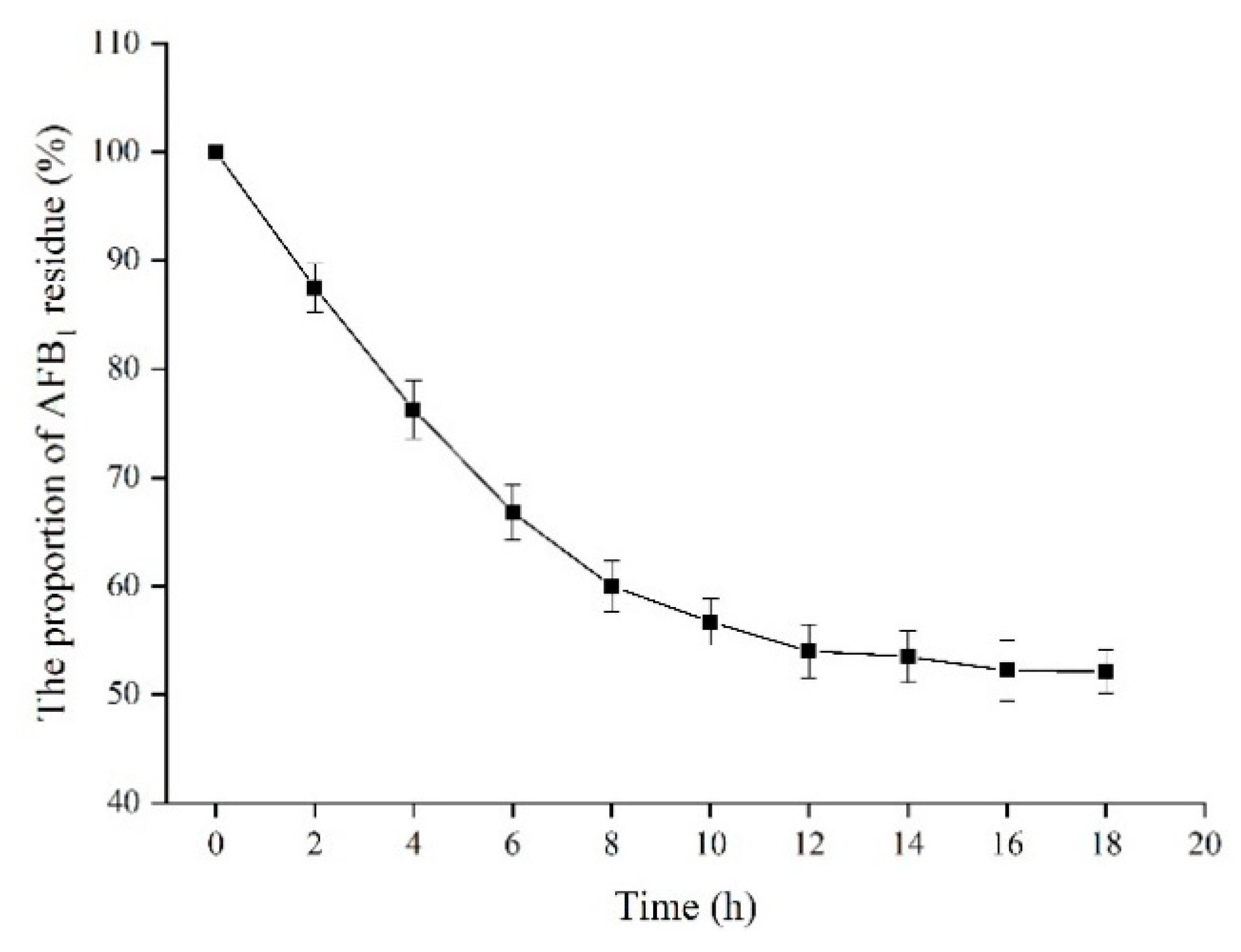
2.9. Effect of Time on the Proportion of AFB1 Residue in Peanuts
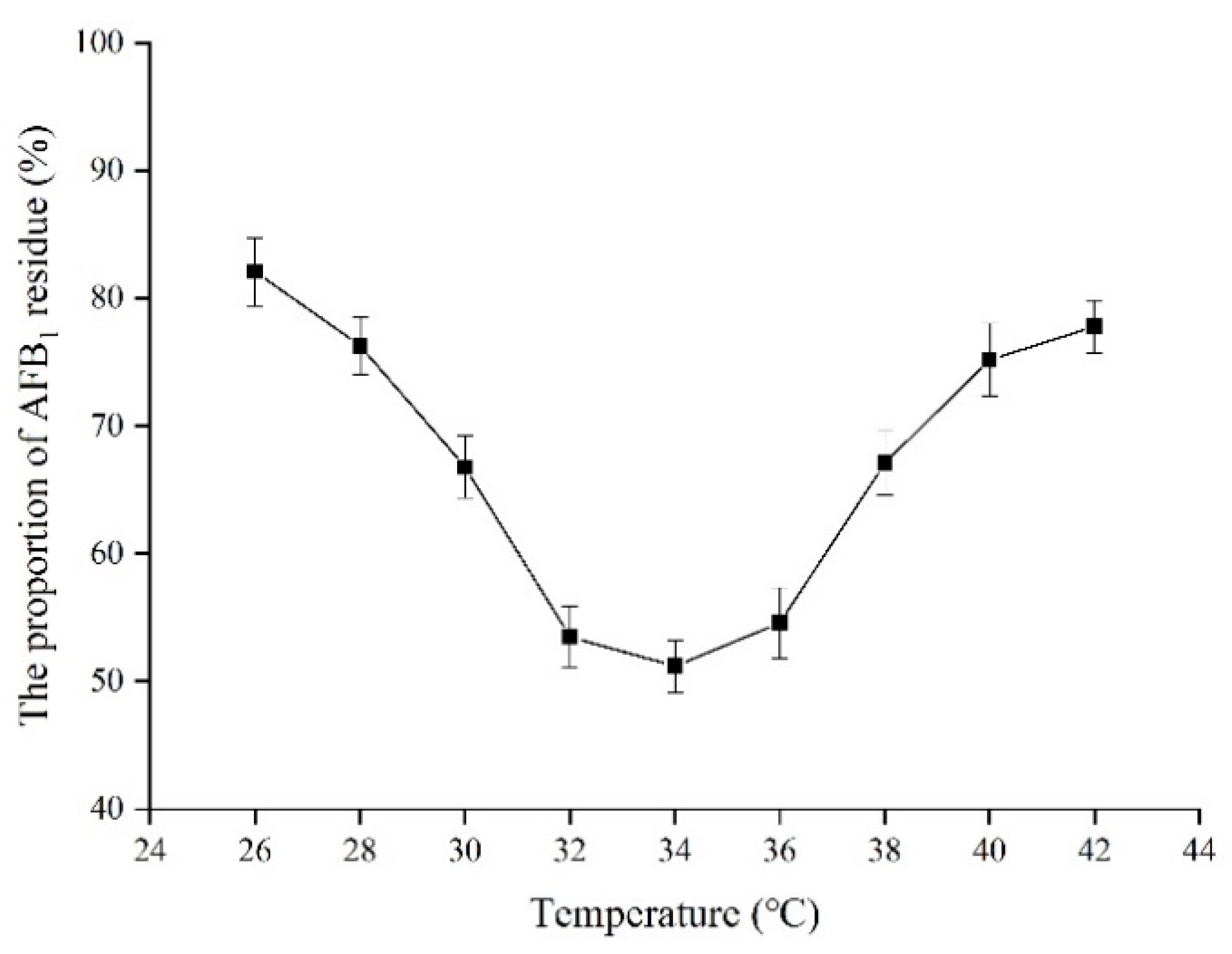
2.10. Effect of Temperature on the Proportion of AFB1 Residue in Peanuts
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials and Reagents
5.2. Construction of pPIC9K-His-TV-AFB1D Vector
5.3. Transformation of pPIC9K-His-TV-AFB1D into P. pastoris GS115
5.4. Inducible Expression and Condition Optimizations of TV-AFB1D
5.5. Purification of Recombinant TV-AFB1D and Definition of Activity
5.6. Measurement of AFB1 Content and Degradation Products
5.7. Application in the AFB1 Degradation of AFB1-Contaminated Peanuts
5.8. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanvi, M.D.; Lawson-Evi, P.; Bouka, E.C.; Eklu-Gadegbeku, K. Aflatoxins in maize dough and dietary exposure in rural populations of Togo. Food Control 2021, 121, 107673. [Google Scholar] [CrossRef]
- Fouche, T.; Claassens, S.; Maboeta, M. Aflatoxins in the soil ecosystem: An overview of its occurrence, fate, effects and future perspectives. Mycotoxin Res. 2020, 36, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Pecorelli, I.; Guarducci, N.; von Holst, C.; Bibi, R.; Pascale, M.; Ciasca, B.; Logrieco, A.F.; Lattanzio, V.M.T. Critical comparison of analytical performances of two immunoassay methods for rapid detection of aflatoxin M-1 in milk. Toxins 2020, 12, 270. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Yu, T.; Qin, W.; Liao, X.; Huang, J.; Liu, Z.; Yu, L.; Liu, X.; Chen, Z.; Yang, C.; et al. Genome-wide association study of the TP53 R249S mutation in hepatocellular carcinoma with aflatoxin B1 exposure and infection with hepatitis B virus. J. Gastrointest. Oncol. 2020, 11, 1333–1339. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.S.; Dunn, J.J.; DeLucca, A.J.; Ciegler, A. Role of lactone ring of aflatoxin B1 in toxicity and mutagenicity. Experientia 1981, 37, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, I.; Audenaert, K.; De Gelder, L. Biodegradation of Mycotoxins: Tales from Known and Unexplored Worlds. Front. Microbiol. 2016, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Guengerich, F.P.; Johnson, W.W.; Ueng, Y.F.; Yamazaki, H.; Shimada, T. Involvement of cytochrome P450, glutathione S-transferase, and epoxide hydrolase in the metabolism of aflatoxin B1 and relevance to risk of human liver cancer. Environ. Health Perspect. 1996, 104 (Suppl. S3), 557–562. [Google Scholar] [CrossRef] [PubMed]
- Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.C.; Nebbia, C.S.; et al. Risk assessment of aflatoxins in food. EFSA J. 2020, 18, 1–112. [Google Scholar] [CrossRef]
- Ji, J.M.; Xie, W.L. Removal of aflatoxin B-1 from contaminated peanut oils using magnetic attapulgite. Food Chem. 2021, 339, 128072. [Google Scholar] [CrossRef]
- Javanmardi, F.; Khodaei, D.; Sheidaei, Z.; Bashiry, M.; Nayebzadeh, K.; Vasseghian, Y.; Khaneghah, A.M. Decontamination of aflatoxins in edible oils: A comprehensive review. Food Rev. Int. 2020, 17, 1–17. [Google Scholar] [CrossRef]
- Magagnoli, S.; Lanzoni, A.; Masetti, A.; Depalo, L.; Albertini, M.; Ferrari, R.; Spadola, G.; Degola, F.; Restivo, F.M.; Burgio, G. Sustainability of strategies for Ostrinia nubilalis management in Northern Italy: Potential impact on beneficial arthropods and aflatoxin contamination in years with different meteorological conditions. Crop Prot. 2021, 142, 105529. [Google Scholar] [CrossRef]
- Adeyeye, S.A.O. Aflatoxigenic fungi and mycotoxins in food: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 709–721. [Google Scholar] [CrossRef]
- Kabak, B. The fate of mycotoxins during thermal food processing. J. Sci. Food Agric. 2009, 89, 549–554. [Google Scholar] [CrossRef]
- Mao, J.; He, B.; Zhang, L.X.; Li, P.W.; Zhang, Q.; Ding, X.X.; Zhang, W. A Structure Identification and Toxicity Assessment of the Degradation Products of Aflatoxin B-1 in Peanut Oil under UV Irradiation. Toxins 2016, 8, 332. [Google Scholar] [CrossRef] [PubMed]
- Samarajeewa, U.; Sen, A.C.; Cohen, M.D.; Wei, C.I. Detoxification of Aflatoxins in Foods and Feeds by Physical and Chemical Methods 1. J. Food Prot. 1990, 53, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Desheng, Q.; Fan, L.; Yanhu, Y.; Niya, Z. Adsorption of aflatoxin B-1 on montmorillonite. Poult. Sci. 2005, 84, 959–961. [Google Scholar] [CrossRef] [PubMed]
- Pankaj, S.K.; Shi, H.; Keener, K.M. A review of novel physical and chemical decontamination technologies for aflatoxin in food. Trends Food Sci. Technol. 2018, 71, 73–83. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, L.; Ma, Q.; Ji, C. Novel strategies for degradation of aflatoxins in food and feed: A review. Food Res. Int. 2021, 140, 109878. [Google Scholar] [CrossRef]
- Mendez-Albores, A.; Martinez-Bustos, F.; Gaytan-Martinez, M.; Moreno-Martinez, E. Effect of lactic and citric acid on the stability of B-aflatoxins in extrusion-cooked sorghum. Lett. Appl. Microbiol. 2008, 47, 1–7. [Google Scholar] [CrossRef]
- Ismail, A.; Goncalves, B.L.; de Neeff, D.V.; Ponzilacqua, B.; Coppa, C.; Hintzsche, H.; Sajid, M.; Cruz, A.G.; Corassin, C.H.; Oliveira, C.A.F. Aflatoxin in foodstuffs: Occurrence and recent advances in decontamination. Food Res. Int. 2018, 113, 74–85. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Gbashi, S.; Nwinyi, O.C.; Mavumengwana, V. Review on microbial degradation of aflatoxins. Crit. Rev. Food Sci. Nutr. 2017, 57, 3208–3217. [Google Scholar] [CrossRef]
- Guo, Y.P.; Qin, X.J.; Tang, Y.; Ma, Q.G.; Zhang, J.Y.; Zhao, L.H. CotA laccase, a novel aflatoxin oxidase from Bacillus licheniformis, transforms aflatoxin B-1 to aflatoxin Q(1) and epi-aflatoxin Q(1). Food Chem. 2020, 325, 126877. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Renaud, J.B.; Rosini, E.; Pollegioni, L.; Vignali, E.; Haidukowski, M.; Sumarah, M.W.; Logrieco, A.F.; Mule, G. Enzymatic transformation of aflatoxin B-1 by Rh_DypB peroxidase and characterization of the reaction products. Chemosphere 2020, 250, 126296. [Google Scholar] [CrossRef]
- Li, C.H.; Li, W.Y.; Hsu, I.N.; Liao, Y.Y.; Yang, C.Y.; Taylor, M.C.; Liu, Y.F.; Huang, W.H.; Chang, H.H.; Huang, H.L.; et al. Recombinant aflatoxin-degrading F420H2-dependent reductase from Mycobacterium smegmatis protects mammalian cells from aflatoxin toxicity. Toxins 2019, 11, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.W.; Wang, L.Z.; Sun, J.D.; Wang, L.P.; Guo, H.Y.; Ye, Y.L.; Sun, X.L. Microbial detoxification of mycotoxins in food and feed. Crit. Rev. Food Sci. Nutr. 2021, 1, 1–19. [Google Scholar] [CrossRef]
- Brana, M.T.; Sergio, L.; Haidukowski, M.; Logrieco, A.F.; Altomare, C. Degradation of aflatoxin B-1 by a sustainable enzymatic extract from spent mushroom substrate of Pleurotus eryngii. Toxins 2020, 12, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.X.; Wu, X.Y.; Xie, C.F.; Hu, Y.D.; Liu, D.L.; Ma, Y.; Yao, D.S. A rational design for improving the trypsin resistance of aflatoxin-detoxifizyme (ADTZ) based on molecular structure evaluation. Enzym. Microb. Technol. 2016, 86, 84–92. [Google Scholar] [CrossRef]
- Huang, B.; Xiao, H.L.; Zhang, J.; Zhang, L.F.; Yang, H.L.; Zhang, Y.; Jin, J. Dual-label time-resolved fluoroimmunoassay for simultaneous detection of aflatoxin B1 and ochratoxin A. Arch. Toxicol. 2009, 83, 619–624. [Google Scholar] [CrossRef]
- Zhao, L.H.; Guan, S.; Gao, X.; Ma, Q.G.; Lei, Y.P.; Bai, X.M.; Ji, C. Preparation, purification and characteristics of an aflatoxin degradation enzyme from Myxococcus fulvus ANSM068. J. Appl. Microbiol. 2011, 110, 147–155. [Google Scholar] [CrossRef]
- Xu, L.; Ahmed, M.F.E.; Sangare, L.; Zhao, Y.J.; Selvaraj, J.N.; Xing, F.G.; Wang, Y.; Yang, H.P.; Liu, Y. Novel aflatoxin-degrading enzyme from Bacillus shackletonii L7. Toxins 2017, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.L.; Wang, W.; Zhang, S.J. Purification and identification of an aflatoxin B-1 degradation enzyme from Pantoea sp. T6. Toxicon 2019, 157, 35–42. [Google Scholar] [CrossRef]
- Wang, X.L.; Qin, X.; Hao, Z.Z.; Luo, H.Y.; Yao, B.; Su, X.Y. Degradation of four major mycotoxins by eight manganese peroxidases in presence of a dicarboxylic acid. Toxins 2019, 11, 566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yehia, R.S. Aflatoxin detoxification by manganese peroxidase purified from Pleurotus ostreatus. Braz. J. Microbiol. 2014, 45, 127–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.L.; Bai, Y.G.; Huang, H.Q.; Tu, T.; Wang, Y.; Wang, Y.R.; Luo, H.Y.; Yao, B.; Su, X.Y. Degradation of aflatoxin B-1 and zearalenone by Bacterial and fungal laccases in presence of structurally defined chemicals and complex natural mediators. Toxins 2019, 11, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loi, M.; Fanelli, F.; Zucca, P.; Liuzzi, V.C.; Quintieri, L.; Cimmarusti, M.T.; Monaci, L.; Haidukowski, M.; Logrieco, A.F.; Sanjust, E.; et al. Aflatoxin B₁ and M₁ degradation by lac2 from Pleurotus pulmonarius and redox mediators. Toxins 2016, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, P.; Yang, J.; Gu, H. Expression of HpaI in Pichia pastoris and optimization of conditions for the heparinase I production. Carbohydr. Polym. 2014, 106, 223–229. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Sources and Enzymes | Conditions | Catalytic Efficiency and Time |
|---|---|---|
| Mycobacterium smegmatis, F420H2-dependent reductase [25] | 20 mM Tris-HCl, pH 7.5 | 63%, 4 h |
| Myxococcus fulvus, MADE [30] | pH 6 | 96% |
| Armillariella tabescens, ADTZ [28] | 0.02 M PBS, pH 6 | 80% |
| Phanerochaete sordida, peroxidase [33] | 50 mM PBS, pH 6 | 86%, 48 h |
| Pleurotus ostreatus, peroxidase [34] | pH 4.5 | 90%, 48 h |
| Bacillus subtilis, BsCotA laccase [35] | 50 mM Tris–HCl, pH 7.0 | 98%, 10 h |
| Pleurotus pulmonarius, laccase [36] | 1 mM sodium acetate, pH 5 | 90%, 72 h |
| Aspergillus flavus, HRP [29] | 1mM sodium acetate, pH 5 | 42%, 1 h |
| Bacillus shackletonii, BADE [31] | 50 mM PBS, pH 7.0 | 47.5%, 72 h |
| Pseudomonas aeruginosa, PADE [32] | 0.02 M PBS, pH 7 | 72.5%, 12h |
| T. versicolor, TV-AFB1D, in this study | AFB1 standard sample | 75.9%, 12 h |
| T. versicolor, TV-AFB1D, in this study | AFB1-contaminated peanuts | 48.5%, 18 h |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.; Xiao, W.; Lu, S.; Jiang, S.; Zheng, Z.; Zhang, D.; Zhang, M.; Jiang, S.; Jiang, S. Recombinant Expression of Trametes versicolor Aflatoxin B1-Degrading Enzyme (TV-AFB1D) in Engineering Pichia pastoris GS115 and Application in AFB1 Degradation in AFB1-Contaminated Peanuts. Toxins 2021, 13, 349. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050349
Yang P, Xiao W, Lu S, Jiang S, Zheng Z, Zhang D, Zhang M, Jiang S, Jiang S. Recombinant Expression of Trametes versicolor Aflatoxin B1-Degrading Enzyme (TV-AFB1D) in Engineering Pichia pastoris GS115 and Application in AFB1 Degradation in AFB1-Contaminated Peanuts. Toxins. 2021; 13(5):349. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050349
Chicago/Turabian StyleYang, Peizhou, Wei Xiao, Shuhua Lu, Suwei Jiang, Zhi Zheng, Danfeng Zhang, Min Zhang, Shaotong Jiang, and Shuying Jiang. 2021. "Recombinant Expression of Trametes versicolor Aflatoxin B1-Degrading Enzyme (TV-AFB1D) in Engineering Pichia pastoris GS115 and Application in AFB1 Degradation in AFB1-Contaminated Peanuts" Toxins 13, no. 5: 349. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050349