Is the Cyanobacterial Bloom Composition Shifting Due to Climate Forcing or Nutrient Changes? Example of a Shallow Eutrophic Reservoir

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Seasonal Cycle
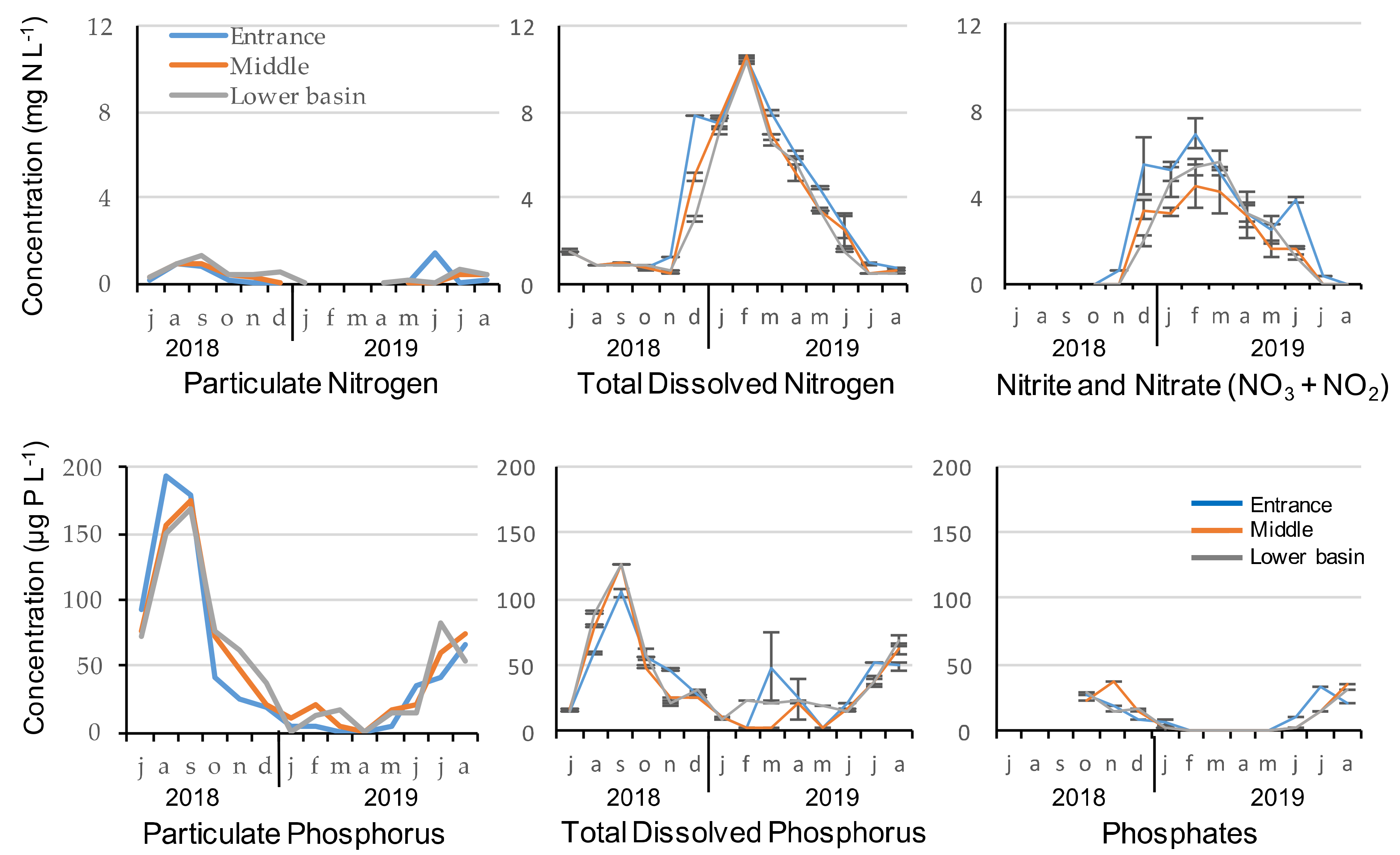
2.1.1. Nutrients’ Concentration
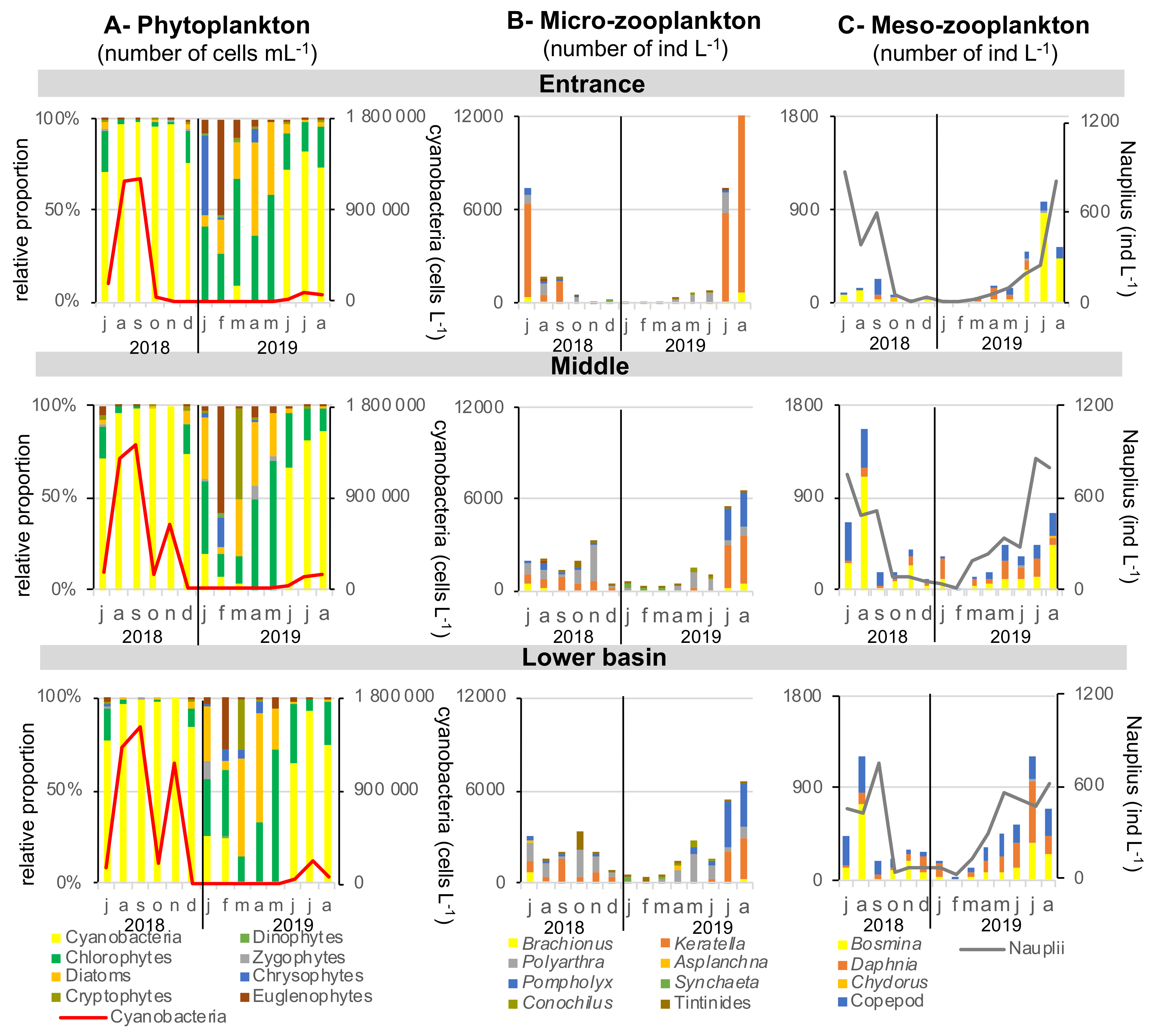
2.1.2. Phytoplankton
2.1.3. Zooplankton
2.2. Inter-Annual Variability of Summer Periods
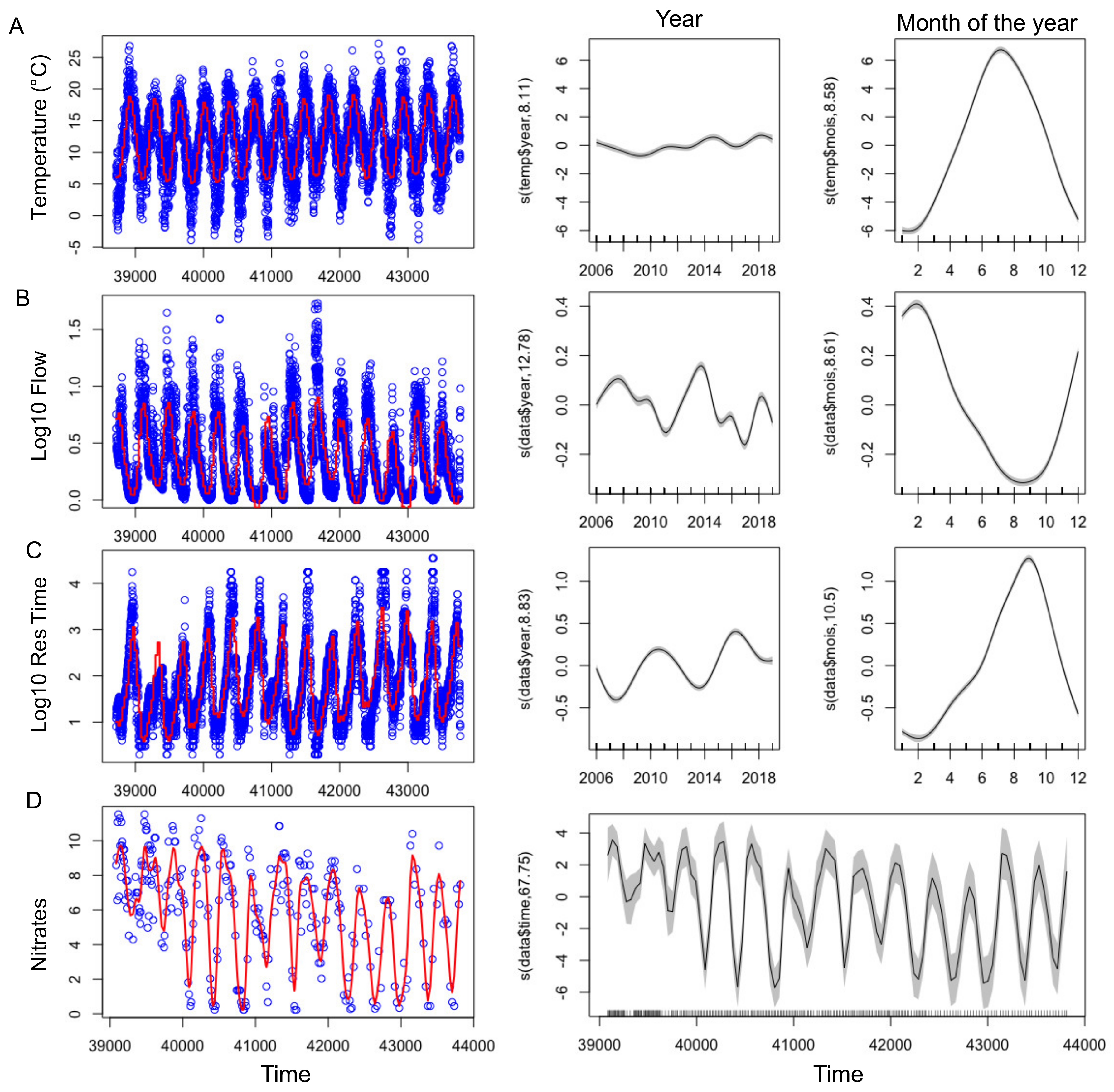
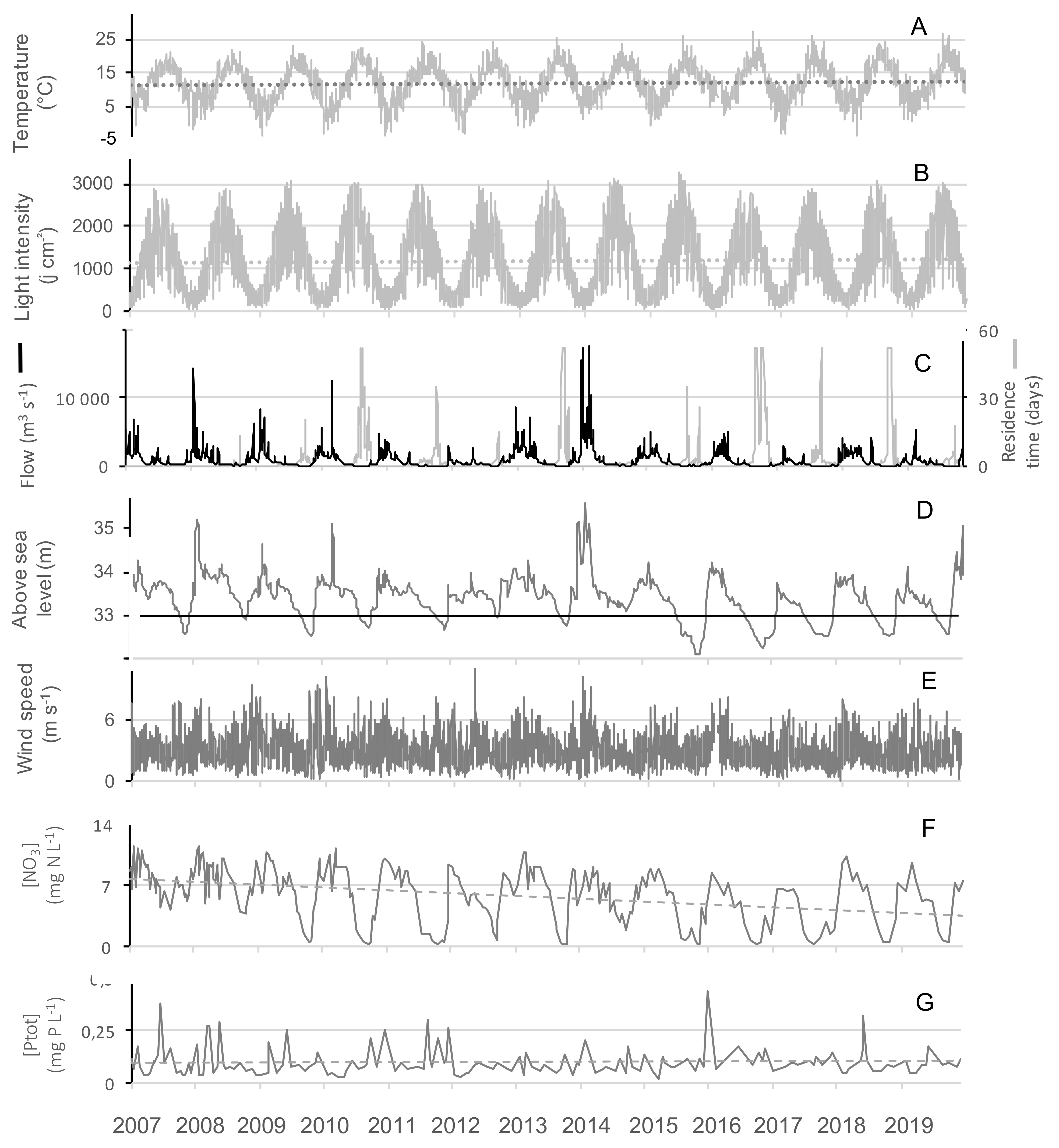
2.2.1. Abiotic Parameters
2.2.2. Nutrients
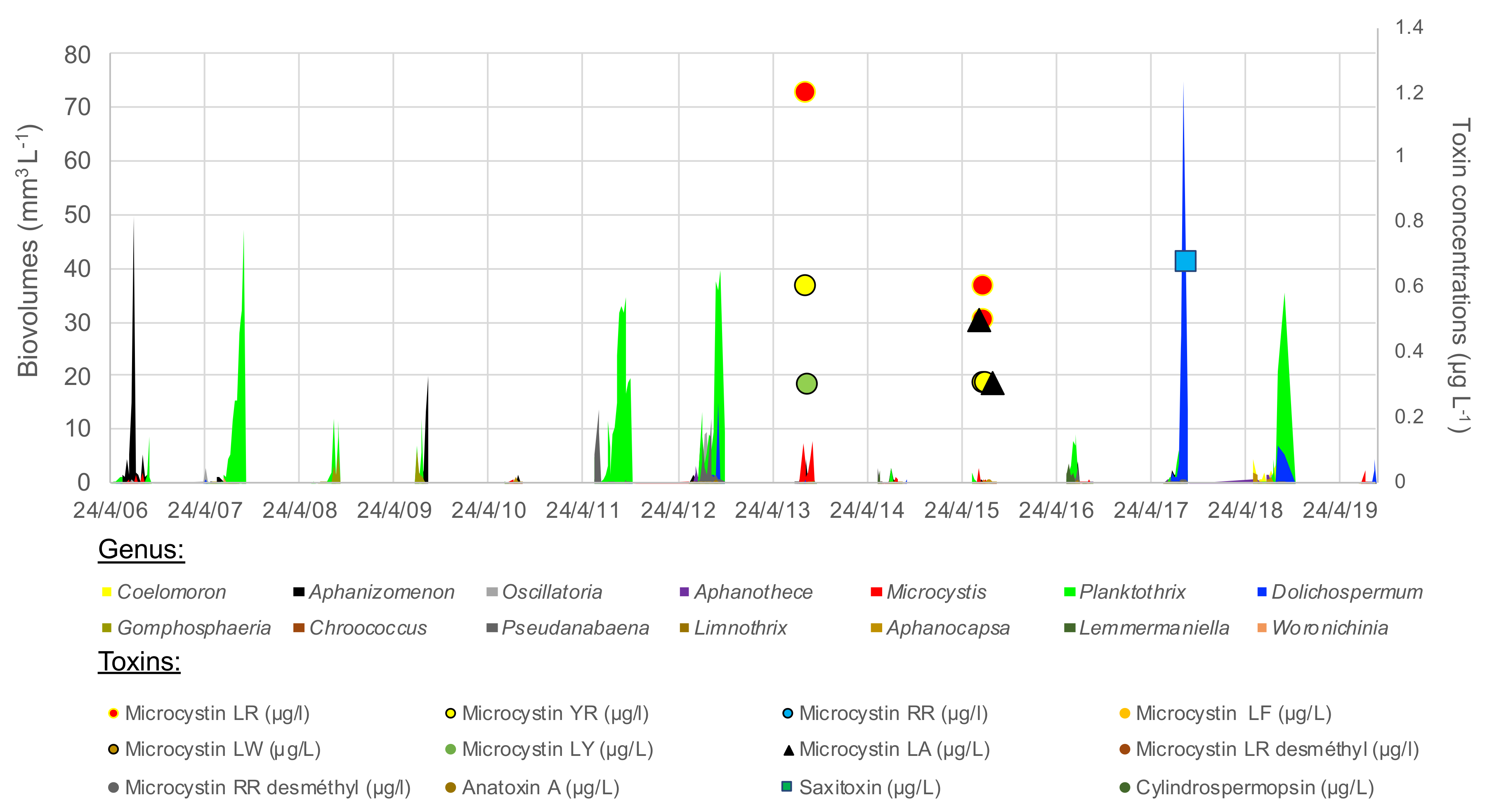
2.2.3. Cyanobacterial Blooms
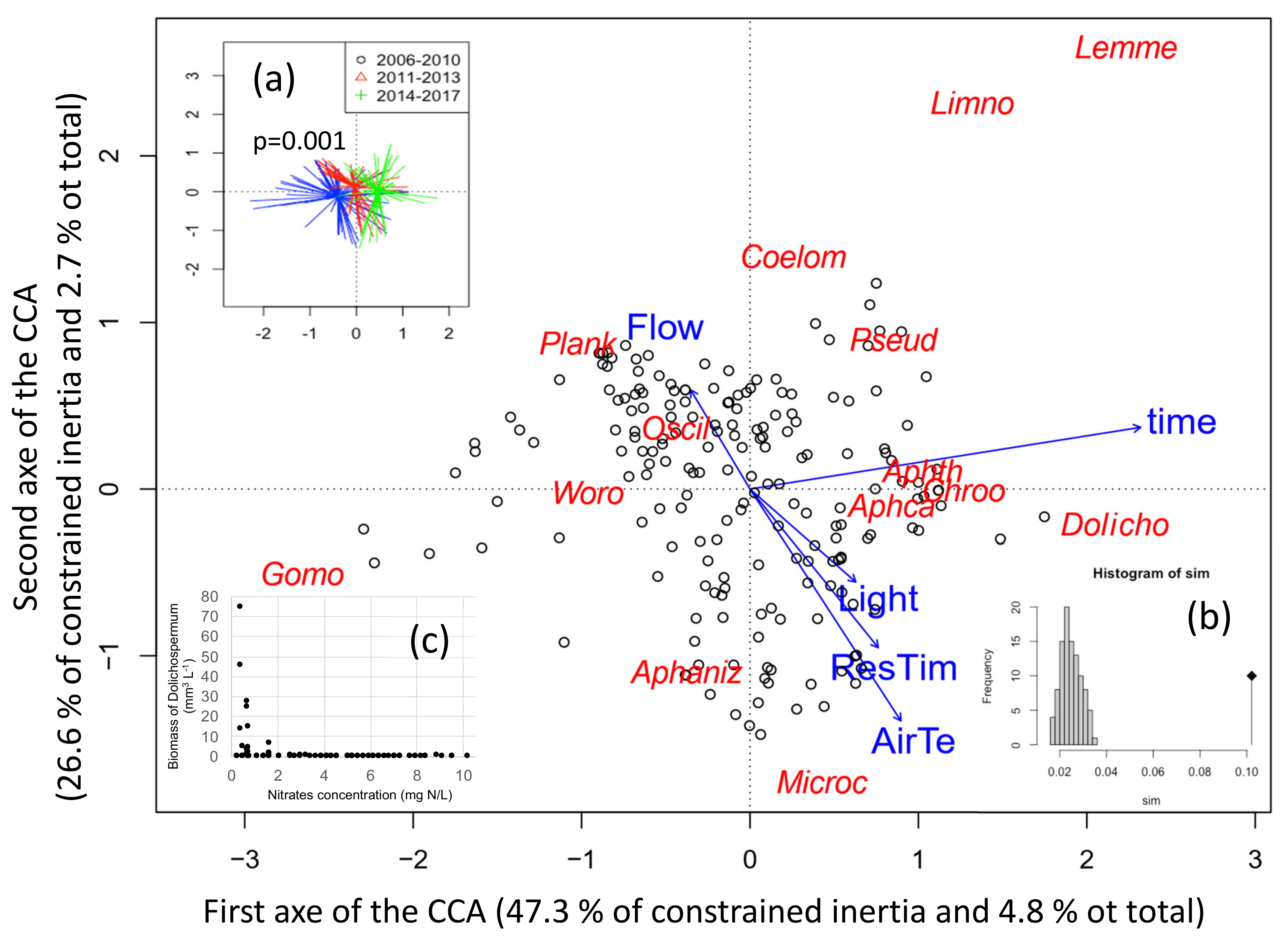
2.2.4. Coupling Blooming Species with Environmental Parameters
3. Discussion
4. Conclusions
5. Materials and Methods
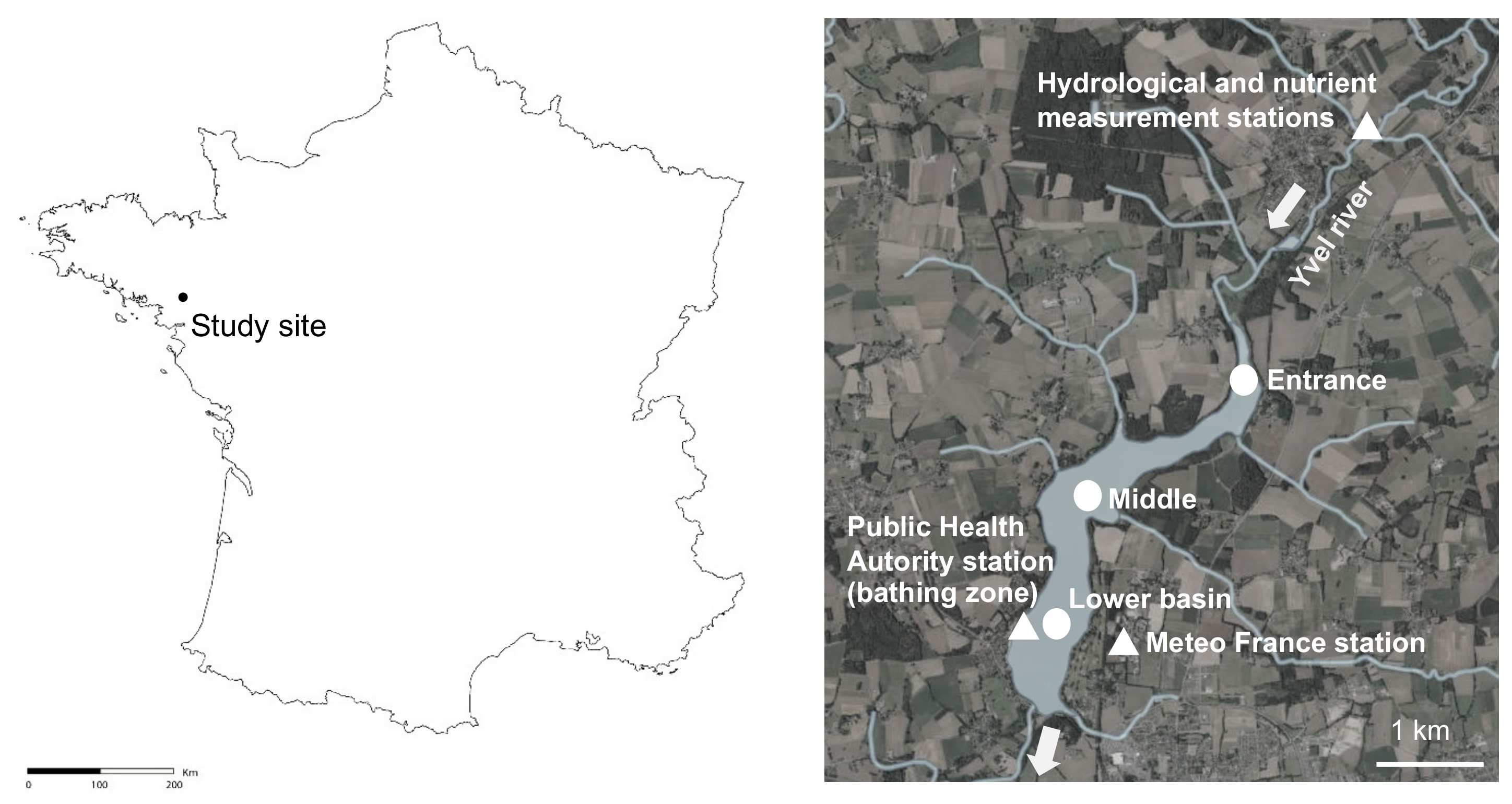
5.1. Study Site
5.2. Seasonal Cycle 2018–2019
5.2.1. Sampling Sites and Sampling
5.2.2. Nutrients
5.2.3. Phytoplankton
5.2.4. Zooplankton
5.3. Long Term Series 2007 to 2019
5.3.1. Phytoplankton and Toxins
5.3.2. Abiotic Parameters
5.3.3. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, V.H.; Tilman, G.D.; Nekola, J.C. Eutrophication: Impacts of Excess Nutrient Inputs on Freshwater, Marine, and Terrestrial Ecosystems. Environ. Pollut. 1999, 100, 179–196. [Google Scholar] [CrossRef]
- Le Moal, M.; Gascuel-Odoux, C.; Menesguen, A.; Souchon, Y.; Etrillard, C.; Levain, A.; Moatar, F.; Pannard, A.; Souchu, P.; Lefebvre, A.; et al. Eutrophication: A New Wine in an Old Bottle? Sci. Total Environ. 2019, 651, 1–11. [Google Scholar] [CrossRef] [Green Version]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The Rise of Harmful Cyanobacteria Blooms: The Potential Roles of Eutrophication and Climate Change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Blooms Bite the Hand That Feeds Them. Science 2013, 342, 433–434. [Google Scholar] [CrossRef]
- Ger, K.A.; Hansson, L.A.; Lurling, M. Understanding Cyanobacteria-Zooplankton Interactions in a More Eutrophic World. Freshw. Biol. 2014, 59, 1783–1798. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial Blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Bockwoldt, K.A.; Nodine, E.R.; Mihuc, T.B.; Shambaugh, A.D.; Stockwell, J.D. Reduced Phytoplankton and Zooplankton Diversity Associated with Increased Cyanobacteria in Lake Champlain, USA. J. Contemp. Water Res. Educ. 2017, 160, 100–118. [Google Scholar] [CrossRef] [Green Version]
- Nurnberg, G.K.; LaZerte, B.D. More than 20 Years of Estimated Internal Phosphorus Loading in Polymictic, Eutrophic Lake Winnipeg, Manitoba. J. Great Lakes Res. 2016, 42, 18–27. [Google Scholar] [CrossRef]
- Carmichael, W.W. The Cyanotoxins. Adv. Bot. Res. 1997, 211–256. [Google Scholar] [CrossRef]
- Wiegand, C.; Pflugmacher, S. Ecotoxicological Effects of Selected Cyanobacterial Secondary Metabolites a Short Review. Toxicol. Appl. Pharmacol. 2005, 203, 201–218. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Codd, G.A. Cyanotoxins. In Ecology of Cyanobacteria II; Springer: Berlin/Heidelberg, Germany, 2012; pp. 651–675. [Google Scholar]
- World Health Organization Guidelines for Drinking Water Quality, Addendum to Volume 2: Health Criteria and Other Supporting Information; WHO Publications: Geneva, Switzerland, 1998.
- Ibelings, B.W.; Backer, L.C.; Kardinaal, W.E.A.; Chorus, I. Current Approaches to Cyanotoxin Risk Assessment and Risk Management around the Globe. Harmful Algae 2014, 40, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Dokulil, M.T.; Teubner, K. Cyanobacterial Dominance in Lakes. Hydrobiologia 2000, 438, 1–12. [Google Scholar] [CrossRef]
- Taranu, Z.E.; Gregory-Eaves, I.; Leavitt, P.R.; Bunting, L.; Buchaca, T.; Catalan, J.; Domaizon, I.; Guilizzoni, P.; Lami, A.; McGowan, S.; et al. Acceleration of Cyanobacterial Dominance in North Temperate-Subarctic Lakes during the Anthropocene. Ecol. Lett. 2015, 18, 375–384. [Google Scholar] [CrossRef]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer Heatwaves Promote Blooms of Harmful Cyanobacteria: Heatwaves Promote Harmful Cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef] [Green Version]
- Kosten, S.; Huszar, V.L.M.; Becares, E.; Costa, L.S.; van Donk, E.; Hansson, L.A.; Jeppesenk, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; et al. Warmer Climates Boost Cyanobacterial Dominance in Shallow Lakes. Glob. Chang. Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Fastner, J.; Abella, S.; Litt, A.; Morabito, G.; Vörös, L.; Pálffy, K.; Straile, D.; Kümmerlin, R.; Matthews, D.; Phillips, M.G.; et al. Combating Cyanobacterial Proliferation by Avoiding or Treating Inflows with High P Load—Experiences from Eight Case Studies. Aquat. Ecol. 2016, 50, 367–383. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Sondergaard, M.; Jensen, J.P.; Havens, K.E.; Anneville, O.; Carvalho, L.; Coveney, M.F.; Deneke, R.; Dokulil, M.T.; Foy, B.; et al. Lake Responses to Reduced Nutrient Loading—An Analysis of Contemporary Long-Term Data from 35 Case Studies. Freshw. Biol. 2005, 50, 1747–1771. [Google Scholar] [CrossRef]
- Shatwell, T.; Köhler, J. Decreased Nitrogen Loading Controls Summer Cyanobacterial Blooms without Promoting Nitrogen-fixing Taxa: Long-term Response of a Shallow Lake. Limnol. Oceanogr. 2019, 64, S166–S178. [Google Scholar] [CrossRef] [Green Version]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.; Stainton, M.; Parker, B.; Paterson, M.; Beaty, K.; Lyng, M.; Kasian, S. Eutrophication of Lakes Cannot Be Controlled by Reducing Nitrogen Input: Results of a 37-Year Whole-Ecosystem Experiment. Proc. Natl. Acad. Sci. 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Lu, X.H.; Chen, Y.W. The Effects of Temperature and Nutrient Ratios on Microcystis Blooms in Lake Taihu, China: An 11-Year Investigation. Harmful Algae 2011, 10, 337–343. [Google Scholar] [CrossRef]
- Deng, J.; Salmaso, N.; Jeppesen, E.; Qin, B.; Zhang, Y. The Relative Importance of Weather and Nutrients Determining Phytoplankton Assemblages Differs between Seasons in Large Lake Taihu, China. Aquat. Sci. 2019, 81, 48. [Google Scholar] [CrossRef]
- Vasconcelos, V.; Morais, J.; Vale, M. Microcystins and Cyanobacteria Trends in a 14 Year Monitoring of a Temperate Eutrophic Reservoir (Aguieira, Portugal). J. Environ. Monit. 2011, 13, 668–672. [Google Scholar] [CrossRef] [PubMed]
- Rücker, J.; Wiedner, C.; Zippel, P. Factors controlling the dominance of Planktothrix agardhii and Limnothrix redekei in eutrophic shallow lakes. In Shallow Lakes’ 95; Springer: Dordrecht, The Netherlands, 1997; pp. 107–115. [Google Scholar]
- Kurmayer, R.; Deng, L.; Entfellner, E. Role of Toxic and Bioactive Secondary Metabolites in Colonization and Bloom Formation by Filamentous Cyanobacteria Planktothrix. Harmful Algae 2016, 54, 69–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casquin, A.; Gu, S.; Dupas, R.; Petitjean, P.; Gruau, G.; Durand, P. River Network Alteration of C-N-P Dynamics in a Mesoscale Agricultural Catchment. Sci. Total Environ. 2020, 749. [Google Scholar] [CrossRef]
- Sondergaard, M.; Jensen, J.P.; Jeppesen, E. Seasonal Response of Nutrients to Reduced Phosphorus Loading in 12 Danish Lakes. Freshw. Biol. 2005, 50, 1605–1615. [Google Scholar] [CrossRef]
- Dolman, A.M.; Mischke, U.; Wiedner, C. Lake-Type-Specific Seasonal Patterns of Nutrient Limitation in German Lakes, with Target Nitrogen and Phosphorus Concentrations for Good Ecological Status. Freshw. Biol. 2016, 61, 444–456. [Google Scholar] [CrossRef]
- Sondergaard, M.; Jensen, J.P.; Jeppesen, E.; Møller, P.H. Seasonal Dynamics in the Concentrations and Retention of Phosphorus in Shallow Danish Lakes after Reduced Loading. Aquat. Ecosyst. Health Manag. 2002, 5, 19–29. [Google Scholar] [CrossRef]
- Guildford, S.J.; Hecky, R.E. Total Nitrogen, Total Phosphorus, and Nutrient Limitation in Lakes and Oceans: Is There a Common Relationship? Limnol. Oceanogr. 2000, 45, 1213–1223. [Google Scholar] [CrossRef] [Green Version]
- Abrantes, N.; Antunes, S.; Pereira, M.; Gonçalves, F. Seasonal Succession of Cladocerans and Phytoplankton and Their Interactions in a Shallow Eutrophic Lake (Lake Vela, Portugal). Acta Oecologica 2006, 29, 54–64. [Google Scholar] [CrossRef]
- Cazzanelli, M.; Warming, T.P.; Christoffersen, K.S. Emergent and Floating-Leaved Macrophytes as Refuge for Zooplankton in a Eutrophic Temperate Lake without Submerged Vegetation. Hydrobiologia 2008, 605, 113–122. [Google Scholar] [CrossRef]
- Zingel, P.; Haberman, J. A comparison of zooplankton densities and biomass in Lakes Peipsi and Võrtsjärv (Estonia): Rotifers and crustaceans versus ciliates. In European Large Lakes Ecosystem Changes and Their Ecological and Socioeconomic Impacts; Springer: Dordrecht, The Netherlands, 2007; pp. 153–159. [Google Scholar]
- Moody, E.K.; Wilkinson, G.M. Functional Shifts in Lake Zooplankton Communities with Hypereutrophication. Freshw. Biol. 2019, 64, 608–616. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; CRC Press: Boca Raton, FL, USA, 1999; ISBN 1-4822-9506-7. [Google Scholar]
- Barnett, A.J.; Finlay, K.; Beisner, B.E. Functional Diversity of Crustacean Zooplankton Communities: Towards a Trait-Based Classification. Freshw. Biol. 2007, 52, 796–813. [Google Scholar] [CrossRef]
- Obertegger, U.; Flaim, G. Community Assembly of Rotifers Based on Morphological Traits. Hydrobiologia 2015, 753, 31–45. [Google Scholar] [CrossRef]
- Burian, A.; Kainz, M.J.; Schagerl, M.; Yasindi, A. Species-Specific Separation of Lake Plankton Reveals Divergent Food Assimilation Patterns in Rotifers. Freshw. Biol. 2014, 59, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Jensen, J.P.; Sondergaard, M.; Lauridsen, T.; Landkildehus, F. Trophic Structure, Species Richness and Biodiversity in Danish Lakes: Changes along a Phosphorus Gradient. Freshw. Biol. 2000, 45, 201–218. [Google Scholar] [CrossRef]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006; ISBN 1-139-45489-7. [Google Scholar]
- Suda, S. Taxonomic Revision of Water-Bloom-Forming Species of Oscillatorioid Cyanobacteria. Int. J. Syst. Evol. Microbiol. 2002, 52, 1577–1595. [Google Scholar] [CrossRef]
- Pitois, F.; Thoraval, I.; Baures, E.; Thomas, O. Geographical Patterns in Cyanobacteria Distribution: Climate Influence at Regional Scale. Toxins 2014, 6, 509–522. [Google Scholar] [CrossRef]
- Sivonen, K.; Jones, G. Cyanobacterial Toxins. Toxic Cyanobacteria Water A Guide Public Health Conseq. Monit. Manag. 1999, 1, 43–112. [Google Scholar]
- Paerl, H.W.; Huisman, J. Climate—Blooms like It Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [Green Version]
- Dittmann, E.; Neilan, B.A.; Erhard, M.; vonDohren, H.; Borner, T. Insertional Mutagenesis of a Peptide Synthetase Gene That Is Responsible for Hepatotoxin Production in the Cyanobacterium Microcystis Aeruginosa PCC 7806. Mol. Microbiol. 1997, 26, 779–787. [Google Scholar] [CrossRef]
- Kurmayer, R.; Christiansen, G.; Fastner, J.; Borner, T. Abundance of Active and Inactive Microcystin Genotypes in Populations of the Toxic Cyanobacterium Planktothrix Spp. Environ. Microbiol. 2004, 6, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Yepremian, C.; Gugger, M.F.; Briand, E.; Catherine, A.; Berger, C.; Quiblier, C.; Bernard, C. Microcystin Ecotypes in a Perennial Planktothrix Agardhii Bloom. Water Res. 2007, 41, 4446–4456. [Google Scholar] [CrossRef]
- Briand, E.; Gugger, M.; Francois, J.C.; Bernard, C.; Humbert, J.F.; Quiblier, C. Temporal Variations in the Dynamics of Potentially Microcystin-Producing Strains in a Bloom-Forming Planktothrix Agardhii (Cyanobacterium) Population. Appl. Environ. Microbiol. 2008, 74, 3839–3848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiansen, G.; Molitor, C.; Philmus, B.; Kurmayer, R. Nontoxic Strains of Cyanobacteria Are the Result of Major Gene Deletion Events Induced by a Transposable Element. Mol. Biol. Evol. 2008, 25, 1695–1704. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Blom, J.F.; Deng, L.; Pernthaler, J. Integrating Phylogeny, Geographic Niche Partitioning and Secondary Metabolite Synthesis in Bloom-Forming Planktothrix. ISME J. 2015, 9, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Briand, E.; Yéprémian, C.; Humbert, J.; Quiblier, C. Competition between Microcystin-and Non-microcystin-producing Planktothrix Agardhii (Cyanobacteria) Strains under Different Environmental Conditions. Environ. Microbiol. 2008, 10, 3337–3348. [Google Scholar] [CrossRef] [PubMed]
- Briand, E.; Bormans, M.; Quiblier, C.; Salencon, M.J.; Humbert, J.F. Evidence of the Cost of the Production of Microcystins by Microcystis Aeruginosa under Differing Light and Nitrate Environmental Conditions. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- De la Cruz, A.A.; Hiskia, A.; Kaloudis, T.; Chernoff, N.; Hill, D.; Antoniou, M.G.; He, X.; Loftin, K.; O’Shea, K.; Zhao, C.; et al. A Review on Cylindrospermopsin: The Global Occurrence, Detection, Toxicity and Degradation of a Potent Cyanotoxin. Environ. Sci. Process. Impacts 2013, 15, 1979. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Mur, L.R.; Walsby, A.E. Diurnal Changes in Buoyancy and Vertical-Distribution in Populations of Microcystins in 2 Shallow Lakes. J. Plankton Res. 1991, 13, 419–436. [Google Scholar] [CrossRef]
- Huisman, J.; Sharples, J.; Stroom, J.M.; Visser, P.M.; Kardinaal, W.E.A.; Verspagen, J.M.H.; Sommeijer, B. Changes in Turbulent Mixing Shift Competition for Light between Phytoplankton Species. Ecology 2004, 85, 2960–2970. [Google Scholar] [CrossRef] [Green Version]
- Li, X.C.; Dreher, T.W.; Li, R.H. An Overview of Diversity, Occurrence, Genetics and Toxin Production of Bloom-Forming Dolichospermum (Anabaena) Species. Harmful Algae 2016, 54, 54–68. [Google Scholar] [CrossRef] [Green Version]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A Review of the Global Ecology, Genomics, and Biogeography of the Toxic Cyanobacterium, Microcystis Spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.K.; Litchman, E. Effects of Temperature and Nitrogen Availability on the Growth of Invasive and Native Cyanobacteria. Hydrobiologia 2016, 763, 357–369. [Google Scholar] [CrossRef]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The Interaction between Climate Warming and Eutrophication to Promote Cyanobacteria Is Dependent on Trophic State and Varies among Taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef] [Green Version]
- Gobler, C.J.; Burkholder, J.M.; Davis, T.W.; Harke, M.J.; Johengen, T.; Stow, C.A.; Van de Waal, D.B. The Dual Role of Nitrogen Supply in Controlling the Growth and Toxicity of Cyanobacterial Blooms. Harmful Algae 2016, 54, 87–97. [Google Scholar] [CrossRef]
- Chaffin, J.D.; Davis, T.W.; Smith, D.J.; Baer, M.M.; Dick, G.J. Interactions between Nitrogen Form, Loading Rate, and Light Intensity on Microcystis and Planktothrix Growth and Microcystin Production. Harmful Algae 2018, 73, 84–97. [Google Scholar] [CrossRef]
- Dupas, R.; Minaudo, C.; Gruau, G.; Ruiz, L.; Gascuel-Odoux, C. Multidecadal Trajectory of Riverine Nitrogen and Phosphorus Dynamics in Rural Catchments. Water Resour. Res. 2018, 54, 5327–5340. [Google Scholar] [CrossRef]
- Lewis, W.M.; Wurtsbaugh, W.A. Control of Lacustrine Phytoplankton by Nutrients: Erosion of the Phosphorus Paradigm. Int. Rev. Hydrobiol. 2008, 93, 446–465. [Google Scholar] [CrossRef]
- Moss, B.; Jeppesen, E.; Sondergaard, M.; Lauridsen, T.L.; Liu, Z.W. Nitrogen, Macrophytes, Shallow Lakes and Nutrient Limitation: Resolution of a Current Controversy? Hydrobiologia 2013, 710, 3–21. [Google Scholar] [CrossRef]
- Scott, J.T.; Grantz, E.M. N-2 Fixation Exceeds Internal Nitrogen Loading as a Phytoplankton Nutrient Source in Perpetually Nitrogen-Limited Reservoirs. Freshw. Sci. 2013, 32, 849–861. [Google Scholar] [CrossRef]
- Kolzau, S.; Dolman, A.M.; Voss, M.; Wiedner, C. The Response of Nitrogen Fixing Cyanobacteria to a Reduction in Nitrogen Loading. Int. Rev. Hydrobiol. 2018, 103, 5–14. [Google Scholar] [CrossRef]
- Paerl, H.W.; Scott, J.T.; McCarthy, M.J.; Newell, S.E.; Gardner, W.S.; Havens, K.E.; Hoffman, D.K.; Wilhelm, S.W.; Wurtsbaugh, W.A. It Takes Two to Tango: When and Where Dual Nutrient (N & P) Reductions Are Needed to Protect Lakes and Downstream Ecosystems. Environ. Sci. Technol. 2016, 50, 10805–10813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantzouki, E.; Visser, P.M.; Bormans, M.; Ibelings, B.W. Understanding the Key Ecological Traits of Cyanobacteria as a Basis for Their Management and Control in Changing Lakes. Aquat. Ecol. 2016, 50, 333–350. [Google Scholar] [CrossRef]
- Grasshoff, P. Methods of Seawater Analysis. Verlag Chemie. FRG 1983, 419, 61–72. [Google Scholar]
- US EPA United States Environmental Protection Agency. Method 365.1: Determination of Phosphorus by Semi-Automated Colorimetry; O’Dell, J.W., Ed.; Office of Research and Development: Cincinnati, OH, USA, 1993. [Google Scholar]
- US EPA United States Environmental Protection Agency. Method 353.2. Determination of Nitrate-Nitrite Nitrogen by Automated Colorimetry; O’Dell, J.W., Ed.; Office of Research and Development: Cincinnati, OH, USA, 1993. [Google Scholar]
- Brient, L.; Lengronne, M.; Bertrand, E.; Rolland, D.; Sipel, A.; Steinmann, D.; Baudin, I.; Legeas, M.; Le Rouzic, B.; Bormans, M. A Phycocyanin Probe as a Tool for Monitoring Cyanobacteria in Freshwater Bodies. J. Environ. Monit. 2008, 10, 248–255. [Google Scholar] [CrossRef]
- US EPA United States Environmental Protection Agency. Standard Operating Procedure for Zooplankton Analysis (Method LG403). Great Lakes Natl. Off. 2016. Available online: https://www.epa.gov/sites/production/files/2017-01/documents/sop-for-zooplankton-analysis-201607-22pp.pdf (accessed on 1 May 2018).
- Pourriot, R.; Francez, A.-J. Introduction Pratique à La Systématique Des Organismes Des Eaux Continentales Françaises.-8: Rotifères. Publ. De La Société Linnéenne De Lyon 1986, 55, 148–176. [Google Scholar] [CrossRef]
- Amoros, C. Introduction Pratique à La Systématique Des Organismes Des Eaux Continentales Françaises-5. Crustacés Cladocères. Publ. De La Société Linnéenne De Lyon 1984, 53, 72–107. [Google Scholar]
- Méthode Interne ME 230; Laboratoire Départemental d’analyses Du Morbihan: Brittany, France, 2016.
- Wolf, H.U.; Frank, C. Toxicity Assessment of Cyanobacterial Toxin Mixtures. Environ. Toxicol. 2002, 17, 395–399. [Google Scholar] [CrossRef]
- Fastner, J.; Humpage, A. Chapter 2.1.; Hepatotoxic cyclic peptides—Microcystins and Nodularins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; CRC Press: Boca Raton, FL, USA, 2021; pp. 21–52. [Google Scholar]
- US EPA Method LG 401, Standard Operating Procedure for Phytoplankton Analysis. Environ. Monit. Syst. Lab. 2003.
- Rimet, F.; Druart, J.C. A Trait Database for Phytoplankton of Temperate Lakes. Ann. De Limnol. Int. J. Limnol. 2018, 54. [Google Scholar] [CrossRef] [Green Version]
- HydroFrance. Available online: Http://Www.Hydro.Eaufrance.Fr/ (accessed on 10 January 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; Software Version R Version 3.6.1 (2019-07-05); R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N/P Ratios of the Particulate Matter | ||||
|---|---|---|---|---|
| Entrance | Middle | Lower Basin | ||
| 2018 | July | 5.45 | 10.16 | 10.34 |
| August | 10.67 | 12.99 | 14.62 | |
| September | 10.00 | 12.43 | 16.41 | |
| October | 7.61 | 11.62 | 11.18 | |
| November | 1.15 | 12.23 | 13.83 | |
| 2019 | July | 0.28 | 15.45 | 17.80 |
| August | 6.78 | 13.16 | 19.92 | |
| Mean ± SD | 5.99 ± 4.03 | 12.58 ± 1.62 | 14.87 ± 3.46 | |
| Number of Days from June to September | River Inflow in the Reservoir: | Date at Which Downstream Flow is Closed to 0 | Date of Autumn Discharge Beginning | ||||
|---|---|---|---|---|---|---|---|
| With Water Temperature > 20 °C | With Light > 2800 J cm−2 | With Daily Wind > 4m s−1 | Annual River Inflow (m3) | Summer River Inflow (m3) | |||
| 2007 | 2 days | 1 days | 25 days | 104,466,499 | 14,288,573 | outflow all year round | |
| 2008 | 4 days | 6 days | 29 days | 82,705,190 | 6,988,378 | outflow all year round | |
| 2009 | 10 days | 5 days | 11 days | 72,962,554 | 3,228,854 | 26/08/2009 | 17/11/2009 |
| 2010 | 19 days | 15 days | 27 days | 74,001,686 | 900,547 | 27/07/2010 | 07/10/2010 |
| 2011 | 11 days | 9 days | 33 days | 53,371,440 | 879,984 | 24/09/2011 | 15/12/2011 |
| 2012 | 14 days | 3 days | 22 days | 30,433,277 | 3,377,462 | 14/09/2012 | 05/10/2012 |
| 2013 | 25 days | 10 days | 18 days | 105,320,477 | 3,085,603 | 12/09/2013 | 06/11/2013 |
| 2014 | 18 days | 16 days | 7 days | 155,305,037 | 2,709,418 | outflow all year | |
| 2015 | 12 days | 18 days | 18 days | 68,434,762 | 1,434,931 | 05/07/2015 | 23/11/2015 |
| 2016 | 20 days | 1 days | 7 days | 62,350,906 | 2,336,688 | 25/07/2016 | 26/01/2017 |
| 2017 | 23 days | 3 days | 11 days | 19,428,682 | 1,200,269 | 03/07/2017 | 15/12/2017 |
| 2018 | 30 days | 8 days | 10 days | 71,447,098 | 7,710,250 | 23/07/2018 | 07/12/2018 |
| 2019 | 25 days | 13 days | 29 days | 38,914,906 | 2,677,622 | 12/07/2019 | 26/10/2019 |
| Dependant Variable (Number of Data Points) | GAMs Model | Smoothing Functions | Estimated Degrees of Freedom | Fisher Test | p-Value | ||
|---|---|---|---|---|---|---|---|
| Adj. R2 | DE (%) | REML | |||||
| Temperature (n = 5019) | 0.7 | 70.1% | 12598 | S (year) | 8.105 | 12.04 | 4.92 × 10−16 |
| S (month) | 8.584 | 1292.95 | <2 × 10−16 | ||||
| Log10 Res Time (n = 5052) | 0.719 | 72% | 3360.3 | S (year) | 8.829 | 129.8 | <2 × 10−16 |
| S (month, k = 12) | 10.497 | 1073.5 | <2 × 10−16 | ||||
| Nitrates (n = 283) | 0.817 | 86.1% | 612.37 | S (time) | 67.75 | 15.15 | <2 × 10−16 |
| Flow (n = 5027) | 0.651 | 65.2% | −970.35 | S (year) | 12.78 | 71.01 | <2 × 10−16 |
| S (month) | 8.61 | 8.61 | <2 × 10−16 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Moal, M.; Pannard, A.; Brient, L.; Richard, B.; Chorin, M.; Mineaud, E.; Wiegand, C. Is the Cyanobacterial Bloom Composition Shifting Due to Climate Forcing or Nutrient Changes? Example of a Shallow Eutrophic Reservoir. Toxins 2021, 13, 351. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050351
Le Moal M, Pannard A, Brient L, Richard B, Chorin M, Mineaud E, Wiegand C. Is the Cyanobacterial Bloom Composition Shifting Due to Climate Forcing or Nutrient Changes? Example of a Shallow Eutrophic Reservoir. Toxins. 2021; 13(5):351. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050351
Chicago/Turabian StyleLe Moal, Morgane, Alexandrine Pannard, Luc Brient, Benjamin Richard, Marion Chorin, Emilien Mineaud, and Claudia Wiegand. 2021. "Is the Cyanobacterial Bloom Composition Shifting Due to Climate Forcing or Nutrient Changes? Example of a Shallow Eutrophic Reservoir" Toxins 13, no. 5: 351. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050351