Sodium Butyrate More Effectively Mitigates the Negative Effects of High-Concentrate Diet in Dairy Cows than Sodium β-Hydroxybutyrate via Reducing Free Bacterial Cell Wall Components in Rumen Fluid and Plasma


Abstract
:1. Introduction
2. Results
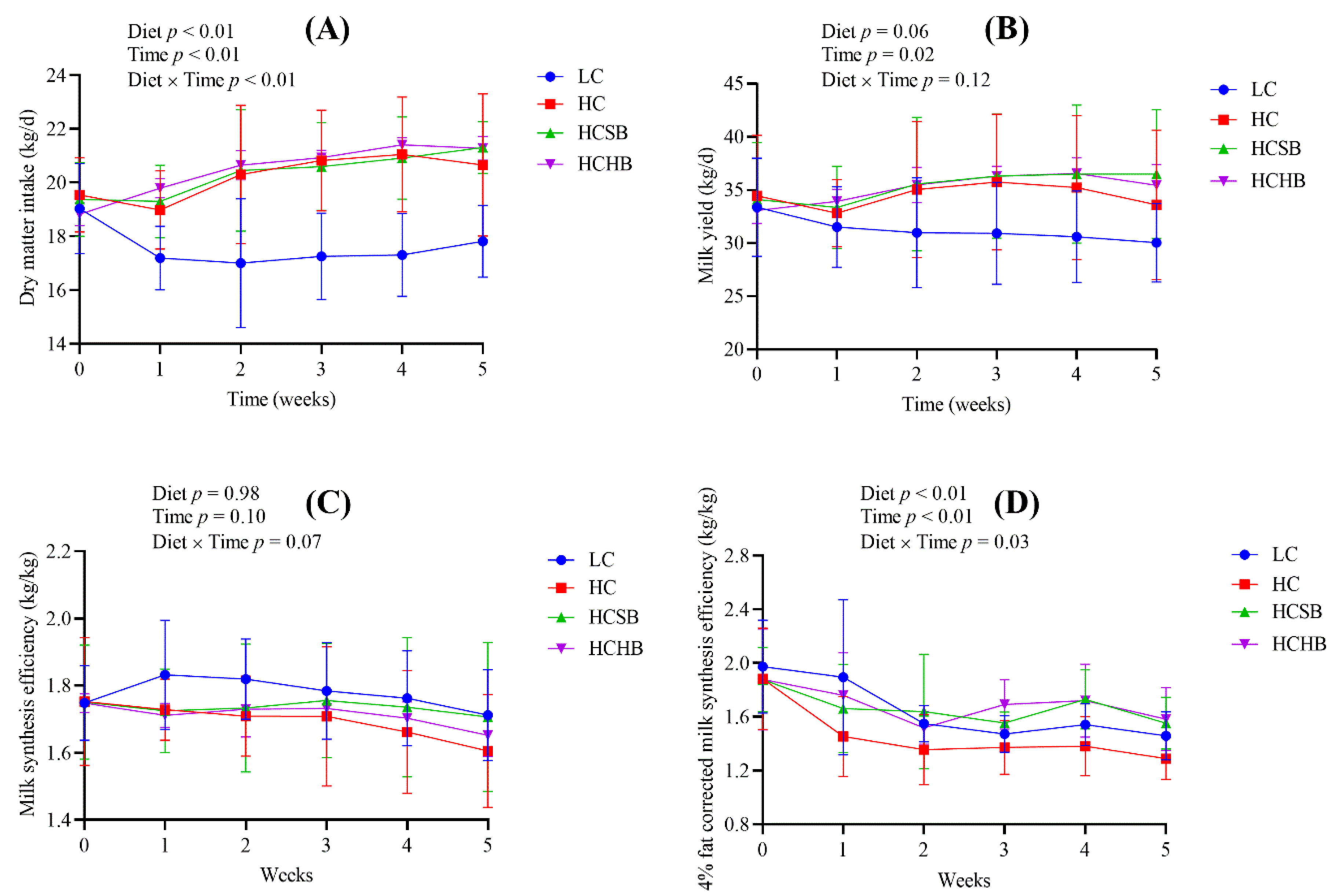
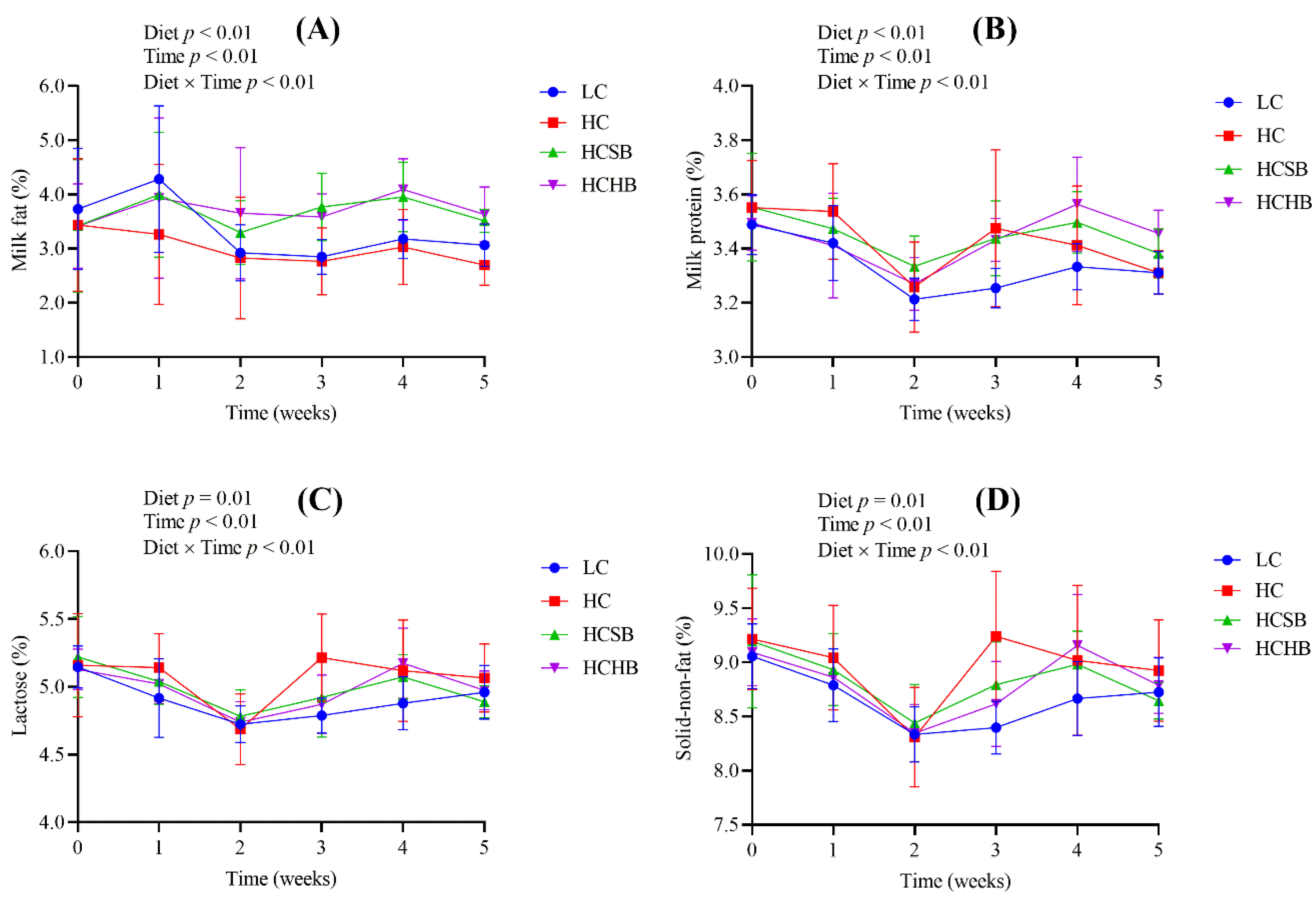
2.1. Production Performance and Milk Components
2.2. Rumen pH and the Concentrations of LPS, PGN and LTA in Rumen Fluid and Plasma
2.3. Concentrations of Proinflammatory Cytokines and Acute Phase Proteins (APPs) in Plasma
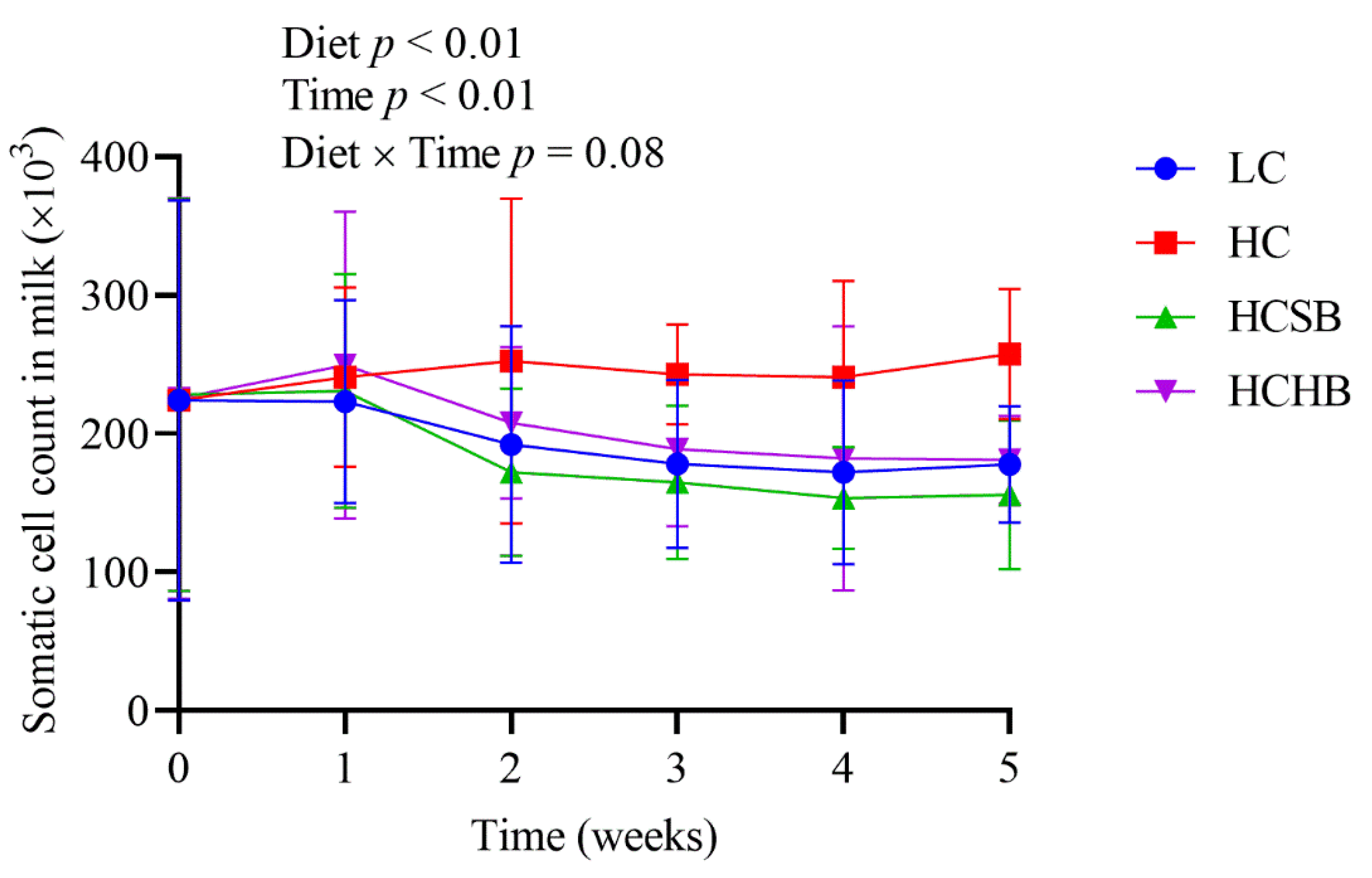
2.4. Blood Leucocyte Indexes and Milk Somatic Cell Count (SCC)
2.5. Blood Glucose and Blood Ketone Bodies
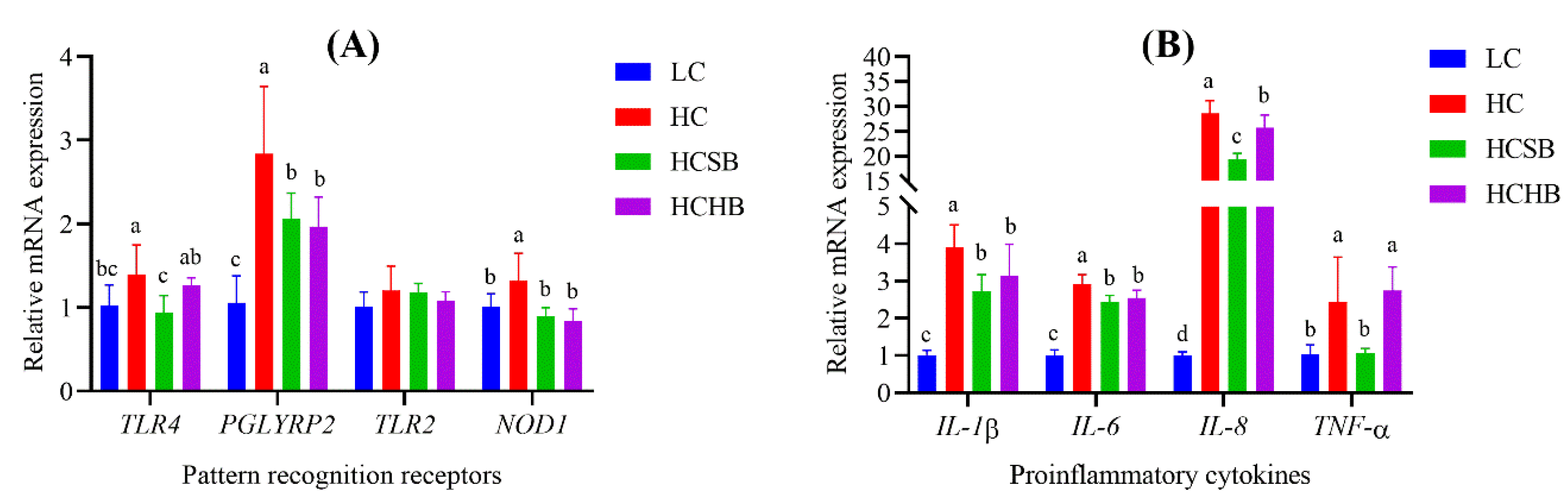
2.6. Gene Expression of PRRs and Proinflammatory Cytokines in Milk Somatic Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethics Statement
5.2. Chemicals and Reagents
5.3. Experimental Design, Dairy Cows, and Diets
5.4. Sample Collection
5.5. Production Performance Measurement
5.6. Measurement of Bacterial Cell Wall Components
5.7. Detection of Proinflammatory Cytokines and APPs
5.8. Measurements of Blood Leucocyte Indexes and Milk SCC
5.9. Detection of Blood Glucose and Blood Ketone Bodies
5.10. Analysis of Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
5.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Tian, K.; Sun, Y.; Wu, Y.; Chen, J.; Zhang, R.; He, T.; Dong, G. Effects of the acid-base treatment of corn on rumen fermentation and microbiota, inflammatory response and growth performance in beef cattle fed high-concentrate diet. Animal 2020, 14, 1876–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, K.; Liu, J.; Sun, Y.; Wu, Y.; Chen, J.; Zhang, R.; He, T.; Dong, G. Effects of dietary supplementation of inulin on rumen fermentation and bacterial microbiota, inflammatory response and growth performance in finishing beef steers fed high or low-concentrate diet. Anim. Feed Sci. Technol. 2019, 258, 114299. [Google Scholar] [CrossRef]
- Eckel, E.F.; Ametaj, B.N. Invited review: Role of bacterial endotoxins in the etiopathogenesis of periparturient diseases of transition dairy cows. J. Dairy Sci. 2016, 99, 5967–5990. [Google Scholar] [CrossRef]
- Porfirio, S.; Carlson, R.W.; Azadi, P. Elucidating peptidoglycan structure: An analytical toolse. Trends Microbiol. 2019, 27, 653–654. [Google Scholar] [CrossRef]
- Mount, J.A.; Karrow, N.A.; Caswell, J.L.; Boermans, H.J.; Leslie, K.E. Assessment of bovine mammary chemokine gene expression in response to lipopolysaccharide, lipotechoic acid plus peptidoglycan, and CpG oligodeoxynucleotide 2135. Can. J. Vet. Res. 2009, 73, 49–57. [Google Scholar]
- Wu, Y.; Sun, Y.; Dong, X.; Chen, J.; Wang, Z.; Chen, J.; Dong, G. The synergism of PGN, LTA and LPS in inducing transcriptome changes, inflammatory responses and a decrease in lactation as well as the associated epigenetic mechanisms in bovine mammary epithelial cells. Toxins 2020, 12, 387. [Google Scholar] [CrossRef] [PubMed]
- Kayagaki, N.; Wong, M.T.; Stowe, I.B.; Ramani, S.R.; Gonzalez, L.C.; Akashi-Takamura, S.; Miyake, K.; Zhang, J.; Lee, W.P.; Muszynski, A.; et al. Noncanonical inflammasome activation by intracellular LPS independent of TLR4. Science 2013, 341, 1246–1249. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Sim, J.R.; Yun, C.H.; Han, S.H. Lipoteichoic acids as a major virulence factor causing inflammatory responses via Toll-like receptor 2. Arch. Pharmacal Res. 2016, 39, 1519–1529. [Google Scholar] [CrossRef]
- Girardin, S.E.; Boneca, I.G.; Carneiro, L.A.M.; Antignac, A.; Jehanno, M.; Viala, J.; Tedin, K.; Taha, M.K.; Labigne, A.; Zahringer, U.; et al. Nod1 detects a unique muropeptide from Gram-negative bacterial peptidoglycan. Science 2003, 300, 1584–1587. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Meng, M.; Gao, L.; Tu, Y.; Bai, Y. Sodium butyrate improves high-concentrate-diet-induced impairment of runninal epithelium barrier function in goats. J. Agric. Food Chem. 2018, 66, 8729–8736. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Zebeli, Q.; Patra, A.K.; Greco, G.; Amasheh, S.; Penner, G.B. Symposium review: The importance of the ruminal epithelial barrier for a healthy and productive cow. J. Dairy Sci. 2019, 102, 1866–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aabdin, Z.U.; Bilal, M.S.; Dai, H.; Abaker, J.A.; Liu, X.; Benazir, S.; Yan, J.; Shen, X. NOD1/NF-κB signaling pathway inhibited by sodium butyrate in the mammary gland of lactating goats during sub-acute ruminal acidosis. Microb. Pathog. 2018, 122, 58–62. [Google Scholar] [CrossRef]
- Chang, G.; Liu, X.; Ma, N.; Yan, J.; Dai, H.; Roy, A.C.; Shen, X. Dietary addition of sodium butyrate contributes to attenuated feeding-induced hepatocyte apoptosis in dairy goats. J. Agric. Food Chem. 2018, 66, 9995–10002. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Huang, J.; Chang, G.; Roy, A.C.; Gao, Q.; Cheng, X.; Shen, X. Sodium butyrate attenuated iE-DAP induced inflammatory response in the mammary glands of dairy goats fed high-concentrate diet. J. Sci. Food Agric. 2020, 101, 1218–1227. [Google Scholar] [CrossRef]
- Roy, A.C.; Wang, Y.; Zhang, H.; Roy, S.; Dai, H.; Chang, G.; Shen, X. Sodium butyrate mitigates iE-DAP induced inflammation caused by high-concentrate feeding in liver of dairy goats. J. Agric. Food Chem. 2018, 66, 8999–9009. [Google Scholar] [CrossRef]
- Roy, A.C.; Chang, G.; Ma, N.; Wang, Y.; Roy, S.; Liu, J.; Aabdin, Z.-U.; Shen, X. Sodium butyrate suppresses NOD1-mediated inflammatory molecules expressed in bovine hepatocytes during iE-DAP and LPS treatment. J. Cell. Physiol. 2019, 234, 19602–19620. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Liu, X.; Yan, J.; ul Aabdin, Z.; Bilal, M.S.; Shen, X. Sodium butyrate ameliorates high-concentrate diet-induced inflammation in the rumen epithelium of dairy goats. J. Agric. Food Chem. 2017, 65, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, Y.; Sun, Y.; Dong, X.; Wang, Z.; Zhang, Z.; Xiao, Y.; Dong, G. Bacterial endotoxin decreased histone H3 acetylation of bovine mammary epithelial cells and the adverse effect was suppressed by sodium butyrate. BMC Vet. Res. 2019, 15, 267. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Zhang, Y.; Ge, Y.; Li, W.; Cao, Y.; Qu, Y.; Liu, S.; Guo, Y.; Fu, S.; Liu, J. Sodium butyrate promotes milk fat synthesis in bovine mammary epithelial cells via GPR41 and its downstream signalling pathways. Life Sci. 2020, 259, 118375. [Google Scholar] [CrossRef] [PubMed]
- Izumi, K.; Fukumori, R.; Oikawa, S.; Oba, M. Short communication: Effects of butyrate supplementation on the productivity of lactating dairy cows fed diets differing in starch content. J. Dairy Sci. 2019, 102, 11051–11056. [Google Scholar] [CrossRef] [PubMed]
- Naeem, A.; Drackley, J.K.; Stamey, J.; Loor, J.J. Role of metabolic and cellular proliferation genes in ruminal development in response to enhanced plane of nutrition in neonatal Holstein calves. J. Dairy Sci. 2012, 95, 1807–1820. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.H.; Huang, Q.Y.; Tominaga, T.; Liu, C.H.; Suzuki, K. An 8-week ketogenic diet alternated interleukin-6, ketolytic and lipolytic gene expression, and enhanced exercise capacity in mice. Nutrients 2018, 10, 1696. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Li, X.B.; Fu, S.X.; Wu, C.C.; Wang, X.X.; Yu, G.J.; Long, M.; Wang, Z.; Liu, G.W. Alterations of fatty acid beta-oxidation capability in the liver of ketotic cows. J. Dairy Sci. 2012, 95, 1759–1766. [Google Scholar] [CrossRef]
- Shimazu, T.; Hirschey, M.D.; Newman, J.; He, W.; Shirakawa, K.; Le Moan, N.; Grueter, C.A.; Lim, H.; Saunders, L.R.; Stevens, R.D.; et al. Suppression of oxidative stress by beta-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science 2013, 339, 211–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, J.C.; Verdin, E. Ketone bodies as signaling metabolites. Trends Endocrinol. Metab. 2014, 25, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Bauman, D.E.; Griinari, J.M. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr. 2003, 23, 203–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Zhang, S.; Hui, Q.; Lei, L.; Du, X.; Gao, W.; Zhang, R.; Liu, G.; Li, X.; Li, X. Beta-hydroxybutyrate facilitates fatty acids synthesis mediated by sterol regulatory element-binding protein1 in bovine mammary epithelial cells. Cell. Physiol. Biochem. 2015, 37, 2115–2124. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Li, C.; Li, L.; Kuang, M.; Shafiq, M.; Wang, Y.; Yang, M.; Wang, G. Effect of acetate, beta-hydroxybutyrate and their interaction on lipogenic gene expression, triglyceride contents and lipid droplet formation in dairy cow mammary epithelial cells. Vitr. Cell. Dev. Biol. Anim. 2021, 57, 66–75. [Google Scholar] [CrossRef]
- Youm, Y.-H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The ketone metabolite beta-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Puchalska, P.; Crawford, P.A. Multi-dimensional roles of ketone bodies in fuel metabolism, signaling, and therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.B.; Sun, L.L.; Cong, R.H.; Tao, S.Y.; DuanMu, Y.Q.; Tian, J.; Ni, Y.D.; Zhao, R.Q. Changes in milk performance and hepatic metabolism in mid-lactating dairy goats after being fed a high concentrate diet for 10 weeks. Animal 2017, 11, 418–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.L.; Zhang, J.; Zhang, D.M.; Li, J.F.; Lin, Y.H. Effect of a high-concentrate diet on milk components and mammary health in Holstein dairy cows. Genet. Mol. Res. 2017, 16, 16019204. [Google Scholar] [CrossRef]
- Li, L.; Cao, Y.; Xie, Z.L.; Zhang, Y.S. A high-concentrate diet induced milk fat decline via glucagon-mediated activation of AMP-activated protein kinase in dairy cows. Sci. Rep. 2017, 7, 44217. [Google Scholar] [CrossRef]
- Tian, P.; Luo, Y.; Li, X.; Tian, J.; Tao, S.; Hua, C.; Geng, Y.; Ni, Y.; Zhao, R. Negative effects of long-term feeding of high-grain diets to lactating goats on milk fat production and composition by regulating gene expression and DNA methylation in the mammary gland. J. Anim. Sci. Biotechnol. 2017, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.E.; Farag, M.A.; Holvoet, P.; Hanafi, R.S.; Gad, M.Z. A comparative metabolomics approach reveals early biomarkers for metabolic response to acute myocardial infarction. Sci. Rep. 2016, 6, 36359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girma, D.D.; Ma, L.; Wang, F.; Jiang, Q.R.; Callaway, T.R.; Drackley, J.K.; Bu, D.P. Effects of close-up dietary energy level and supplementing rumen-protected lysine on energy metabolites and milk production in transition cows. J. Dairy Sci. 2019, 102, 7059–7072. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.; Zhang, H.; Wang, Y.; Ma, N.; Chandra, R.A.; Ye, G.; Zhuang, S.; Zhu, W.; Shen, X. Microbial community shifts elicit inflammation in the caecal mucosa via the GPR41/43 signalling pathway during subacute ruminal acidosis. BMC Vet. Res. 2019, 15, 298. [Google Scholar] [CrossRef]
- Zhou, J.; Dong, G.; Ao, C.; Zhang, S.; Qiu, M.; Wang, X.; Wu, Y.; Erdene, K.; Jin, L.; Lei, C.; et al. Feeding a high-concentrate corn straw diet increased the release of endotoxin in the rumen and pro-inflammatory cytokines in the mammary gland of dairy cows. BMC Vet. Res. 2014, 10, 172. [Google Scholar] [CrossRef] [Green Version]
- Memon, M.A.; Wang, Y.; Xu, T.; Ma, N.; Zhang, H.; Roy, A.-C.; ul Aabdin, Z.; Shen, X. Lipopolysaccharide induces oxidative stress by triggering MAPK and Nrf2 signalling pathways in mammary glands of dairy cows fed a high-concentrate diet. Microb. Pathog. 2019, 128, 268–275. [Google Scholar] [CrossRef]
- ul Aabdin, Z.; Cheng, X.; Dai, H.; Wang, Y.; Sahito, B.; Roy, A.C.; Memon, M.A.; Shen, X. High-concentrate feeding to dairy cows induces apoptosis via the NOD1/Caspase-8 pathway in mammary epithelial cells. Genes 2020, 11, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, W.; Ma, N.; Wang, L.; Dai, H.; Bilal, M.S.; Roy, A.C.; Shen, X. Overfeeding with a high-concentrate diet activates the NOD1-NF-kappa B signalling pathway in the mammary gland of mid-lactating dairy cows. Microb. Pathog. 2019, 128, 390–395. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, G.; Li, X.; Guan, Y.; Wang, Y.; Yuan, X.; Sun, G.; Wang, Z.; Li, X. Inflammatory mechanism of rumenitis in dairy cows with subacute ruminal acidosis. BMC Vet. Res. 2018, 14, 135. [Google Scholar] [CrossRef]
- Lodemann, U.; Martens, H. Effects of diet and osmotic pressure on Na+ transport and tissue conductance of sheep isolated rumen epithelium. Exp. Physiol. 2006, 91, 539–550. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Girton, A.W.; Popescu, N.I.; Keshari, R.S.; Burgett, T.; Lupu, F.; Coggeshall, K.M. Serum amyloid P and IgG exhibit differential capabilities in the activation of the innate immune system in response to bacillus anthracis peptidoglycan. Infect. Immun. 2018, 86, e00076-18. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Aabdin, Z.-U.; Wang, Y.; Ma, N.; Dai, H.; Shi, X.; Shen, X. Glutamine pretreatment protects bovine mammary epithelial cells from inflammation and oxidative stress induced by gamma-d-glutamyl-meso-diaminopimelic acid (iE-DAP). J. Dairy Sci. 2020, 104, 2123–2139. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the LPS gene product. J. Immunol. 1999, 162, 3749–3752. [Google Scholar]
- Saha, S.; Qi, J.; Wang, S.; Wang, M.; Li, X.; Kim, Y.-G.; Nunez, G.; Gupta, D.; Dziarski, R. PGLYRP-2 and Nod2 are both required for peptidoglycan-induced arthritis and local inflammation. Cell Host Microbe 2009, 5, 137–150. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, A.A.; McCoard, S.A.; Roy, N.C.; Barnett, M.P.G.; Mackenzie, D.D.S.; McNabb, W.C. Gene expression in bovine mammary somatic cells isolated from milk. J. Anim. Feed Sci. 2004, 13, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Canovas, A.; Rincon, G.; Bevilacqua, C.; Islas-Trejo, A.; Brenaut, P.; Hovey, R.C.; Boutinaud, M.; Morgenthaler, C.; VanKlompenberg, M.K.; Martin, P.; et al. Comparison of five different RNA sources to examine the lactating bovine mammary gland transcriptome using RNA-Sequencing. Sci. Rep. 2014, 4, 5297. [Google Scholar] [CrossRef] [Green Version]
- Wickramasinghe, S.; Rincon, G.; Islas-Trejo, A.; Medrano, J.F. Transcriptional profiling of bovine milk using RNA sequencing. BMC Genom. 2012, 13, 45. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. BBA Mol. Cell Res. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Wang, Y.; Loor, J.J.; Bucktrout, R.; Shu, X.; Jia, H.; Dong, J.; Zuo, R.; Liu, G.; Li, X.; et al. High expression of cell death-inducing DFFA-like effector a (CIDEA) promotes milk fat content in dairy cows with clinical ketosis. J. Dairy Sci. 2019, 102, 1682–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, G.; Yan, J.; Ma, N.; Liu, X.; Dai, H.; Bilal, M.S.; Shen, X. Dietary sodium butyrate supplementation reduces high-concentrate diet feeding-induced apoptosis in mammary cells in dairy goats. J. Agric. Food Chem. 2018, 66, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Hillreiner, M.; Flinspach, C.; Pfaffl, M.W.; Kliem, H. Effect of the ketone body beta-hydroxybutyrate on the innate defense capability of primary bovine mammary epithelial cells. PLoS ONE 2016, 11, e0157774. [Google Scholar] [CrossRef]
- Zhang, L.Y. Feed Analysis and Quality Test Technology, 3rd ed.; China Agricultural University Press: Beijing, China, 2007; pp. 10–147. [Google Scholar]
- Shen, J.S.; Chai, Z.; Song, L.J.; Liu, J.X.; Wu, Y.M. Insertion depth of oral stomach tubes may affect the fermentation parameters of ruminal fluid collected in dairy cows. J. Dairy Sci. 2012, 95, 5978–5984. [Google Scholar] [CrossRef] [Green Version]
- Jagtap, P.; Singh, R.; Deepika, K.; Sritharan, V.; Gupta, S. A flowthrough assay for rapid bedside stratification of bloodstream bacterial infection in critically III patients: A pilot study. J. Clin. Microbiol. 2018, 56, e00408. [Google Scholar] [CrossRef] [Green Version]
- Kubicek-Sutherland, J.Z.; Vu, D.M.; Noormohamed, A.; Mendez, H.M.; Stromberg, L.R.; Pedersen, C.A.; Hengartner, A.C.; Klosterman, K.E.; Bridgewater, H.A.; Otieno, V.; et al. Direct detection of bacteremia by exploiting host-pathogen interactions of lipoteichoic acid and lipopolysaccharide. Sci. Rep. 2019, 9, 6203. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Salter, A.M.; Parr, T.; Garnsworthy, P.C. Extraction and quantitative analysis of stearoyl-coenzyme A desaturase mRNA from dairy cow milk somatic cells. J. Dairy Sci. 2007, 90, 4128–4136. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Diet | p-Value | |||
|---|---|---|---|---|---|
| LC | HC | HCSB | HCHB | ||
| rumen pH value | 6.70 ± 0.18 ab | 6.24 ± 0.17 c | 6.72 ± 0.27 a | 6.53 ± 0.16 b | <0.01 |
| rumen LPS (EU/mL) | 25,565 ± 3433 c | 68,777 ± 5978 a | 55,173 ± 6135 b | 65,310 ± 7391 a | <0.01 |
| rumen PGN (μg/mL) | 42.9 ± 3.5 b | 122.4 ± 17.6 a | 106.7 ± 14.4 a | 121.1 ± 50.7 a | <0.01 |
| rumen LTA (μg/mL) | 2.42 ± 0.50 c | 4.16 ± 0.81 a | 3.10 ± 0.56 b | 4.03 ± 0.58 a | <0.01 |
| plasma LPS (EU/mL) | 0.07 ± 0.01 c | 0.23 ± 0.04 a | 0.20 ± 0.02 b | 0.23 ± 0.04 a | <0.01 |
| plasma PGN (pg/mL) | 173.8 ± 149.0 b | 485.6 ± 207.5 a | 200.1 ± 77.8 b | 423.6 ± 197.9 a | <0.01 |
| plasma LTA (ng/mL) | 11.5 ± 1.1 c | 16.9 ± 2.7 a | 14.1 ± 0.9 b | 16.3 ± 2.81 a | <0.01 |
| Item | Diet | p-Value | |||
|---|---|---|---|---|---|
| LC | HC | HCSB | HCHB | ||
| Proinflammatory cytokines | |||||
| IL-1β (pg/mL) | 122.9 ± 11.7 c | 169.4 ± 14.9 a | 140.4 ± 14.3 b | 162.3 ± 6.8 a | <0.01 |
| IL-6 (pg/mL) | 161.1 ± 23.6 b | 203.5 ± 24.0 a | 169.9 ± 14.1 b | 178.9 ± 27.3 b | <0.01 |
| IL-8 (pg/mL) | 272.7 ± 53.7 c | 346.1 ± 15.7 a | 297.1 ± 25.4 bc | 321.8 ± 30.1 ab | <0.01 |
| TNF-α (pg/mL) | 1313 ± 111 b | 1499 ± 66 a | 1346 ± 73 b | 1368 ± 118 b | <0.01 |
| Acute phase proteins (APPs) | |||||
| LBP (μg/mL) | 143.0 ± 25.2 b | 240.8 ± 23.3 a | 227.4 ± 20.5 a | 234.2 ± 20.4 a | <0.01 |
| SAA (μg/mL) | 12.3 ± 1.6 b | 14.8 ± 2.0 a | 13.1 ± 0.8 ab | 14.5 ± 1.9 a | 0.02 |
| CRP (mg/L) | 10.4 ± 1.1 c | 12.5 ± 1.3 a | 10.9 ± 0.8 bc | 11.9 ± 0.9 ab | <0.01 |
| Hp (μg/mL) | 1152 ± 326 c | 1688 ± 169 a | 1315 ± 143 b | 1244 ± 91 b | <0.01 |
| Index | Unit | Diet | p-Value | |||
|---|---|---|---|---|---|---|
| LC | HC | HCSB | HCHB | |||
| Number of white blood cells | 109/L | 17.5 ± 4.9 b | 27.3 ± 5.2 a | 17.2 ± 8.0 b | 15.7 ± 5.3 b | <0.01 |
| Number of lymphocytes | 109/L | 9.9 ± 3.6 b | 15.7 ± 4.1 a | 8.2 ± 6.1 b | 7.3 ± 4.5 b | <0.01 |
| Number of monocytes | 109/L | 0.88 ± 0.29 b | 1.64 ± 0.76 a | 0.97 ± 0.43 b | 1.04 ± 0.43 ab | <0.01 |
| Number of neutrophils | 109/L | 6.7 ± 1.6 b | 9.9 ± 3.2 a | 7.8 ± 2.0 a | 7.3 ± 1.5 a | 0.01 |
| Ratio of lymphocytes | % | 55.2 ± 8.2 a | 57.5 ± 10.2 a | 42.8 ± 14.8 b | 43.3 ± 14.7 b | 0.01 |
| Ratio of monocytes | % | 5.3 ± 1.5 b | 6.1 ± 3.0 ab | 7.6 ± 2.3 a | 6.9 ± 1.8 ab | 0.13 |
| Ratio of neutrophils | % | 39.5 ± 7.1 ab | 36.4 ± 9.7 b | 49.6 ± 13.1 a | 49.8 ± 13.9 a | 0.02 |
| Item | Diet | p-Value | |||
|---|---|---|---|---|---|
| LC | HC | HCSB | HCHB | ||
| Blood glucose (mmol/L) | 2.33 ± 0.41 | 2.52 ± 0.62 | 2.69 ± 0.50 | 2.74 ± 0.46 | 0.27 |
| Blood ketone bodies (mmol/L) | 0.71 ± 0.07 b | 0.62 ± 0.08 c | 0.71 ± 0.07 b | 0.79 ± 0.07 a | <0.01 |
| Blood ketone bodies (mg/dL) | 7.42 ± 0.72 b | 6.66 ± 0.85 c | 7.48 ± 0.58 b | 8.37 ± 0.65 a | <0.01 |
| Item | Diet 1 | |||
|---|---|---|---|---|
| LC | HC | HCSB | HCHB | |
| Ingredient (% DM) | ||||
| Leymus chinensis | 14.81 | 6.04 | 5.98 | 5.98 |
| Corn silage | 22.02 | 13.96 | 13.82 | 13.82 |
| Alfalfa hay | 12.77 | 10.97 | 10.86 | 10.86 |
| Fresh distiller’s grains | 5.87 | 3.64 | 3.60 | 3.60 |
| Sugar beet meal | 4.36 | 3.88 | 3.84 | 3.84 |
| Cottonseed | 0.00 | 1.75 | 1.73 | 1.73 |
| Corn | 21.80 | 32.36 | 32.04 | 32.04 |
| Wheat bran | 1.53 | 3.46 | 3.43 | 3.43 |
| Palm oil | 1.09 | 1.56 | 1.54 | 1.54 |
| Soybean meal | 5.43 | 7.77 | 7.69 | 7.69 |
| Cottonseed meal | 2.77 | 3.97 | 3.93 | 3.93 |
| Distillers dried grain with solubles (DDGS) | 3.93 | 5.62 | 5.56 | 5.56 |
| Calcium hydrogen phosphate | 1.10 | 1.58 | 1.56 | 1.56 |
| Calcium carbonate (light powder) | 0.66 | 0.95 | 0.94 | 0.94 |
| Magnesium oxide | 0.22 | 0.32 | 0.32 | 0.32 |
| Salt | 0.44 | 0.48 | 0.48 | 0.48 |
| Vitamin and mineral premix 2 | 0.88 | 1.26 | 1.25 | 1.25 |
| Urea premix | 0.22 | 0.32 | 0.32 | 0.32 |
| Yeast premix | 0.09 | 0.13 | 0.13 | 0.13 |
| Sodium butyrate | 0.00 | 0.00 | 1.00 | 0.00 |
| Sodium β-hydroxybutyrate | 0.00 | 0.00 | 0.00 | 1.00 |
| Concentrate:forage | 4:6 | 6:4 | 6:4 | 6:4 |
| Nutrient composition 3 | ||||
| Net energy for lactation (NEL, MJ/Kg) | 6.18 | 6.56 | 6.49 | 6.49 |
| Crude protein (% DM) | 15.4 | 16.4 | 16.3 | 16.2 |
| Ether extract (% DM) | 5.2 | 6.1 | 5.4 | 5.8 |
| Neutral detergent fiber (% DM) | 49.3 | 43.8 | 44.4 | 43.7 |
| Acid detergent fiber (% DM) | 22.4 | 18.9 | 18.8 | 18.3 |
| Calcium (% DM) | 1.02 | 1.03 | 1.04 | 1.06 |
| Total phosphorus (% DM) | 0.52 | 0.52 | 0.54 | 0.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Sun, Y.; Zhang, R.; He, T.; Huang, G.; Tian, K.; Liu, J.; Chen, J.; Dong, G. Sodium Butyrate More Effectively Mitigates the Negative Effects of High-Concentrate Diet in Dairy Cows than Sodium β-Hydroxybutyrate via Reducing Free Bacterial Cell Wall Components in Rumen Fluid and Plasma. Toxins 2021, 13, 352. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050352
Wu Y, Sun Y, Zhang R, He T, Huang G, Tian K, Liu J, Chen J, Dong G. Sodium Butyrate More Effectively Mitigates the Negative Effects of High-Concentrate Diet in Dairy Cows than Sodium β-Hydroxybutyrate via Reducing Free Bacterial Cell Wall Components in Rumen Fluid and Plasma. Toxins. 2021; 13(5):352. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050352
Chicago/Turabian StyleWu, Yongjiang, Yawang Sun, Ruiming Zhang, Tianle He, Guohao Huang, Ke Tian, Junhui Liu, Juncai Chen, and Guozhong Dong. 2021. "Sodium Butyrate More Effectively Mitigates the Negative Effects of High-Concentrate Diet in Dairy Cows than Sodium β-Hydroxybutyrate via Reducing Free Bacterial Cell Wall Components in Rumen Fluid and Plasma" Toxins 13, no. 5: 352. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050352