Degradation of the Indospicine Toxin from Indigofera spicata by a Mixed Population of Rumen Bacteria

 and
and 
Abstract
:1. Introduction
2. Results
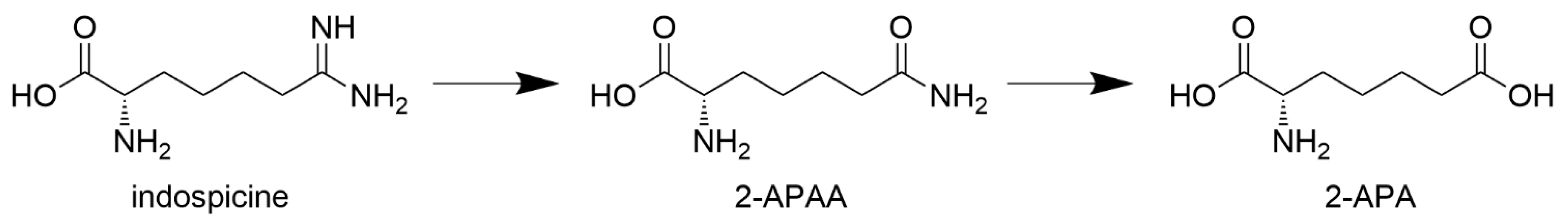
2.1. Fermenter: Concentrations of Indospicine and Breakdown Products
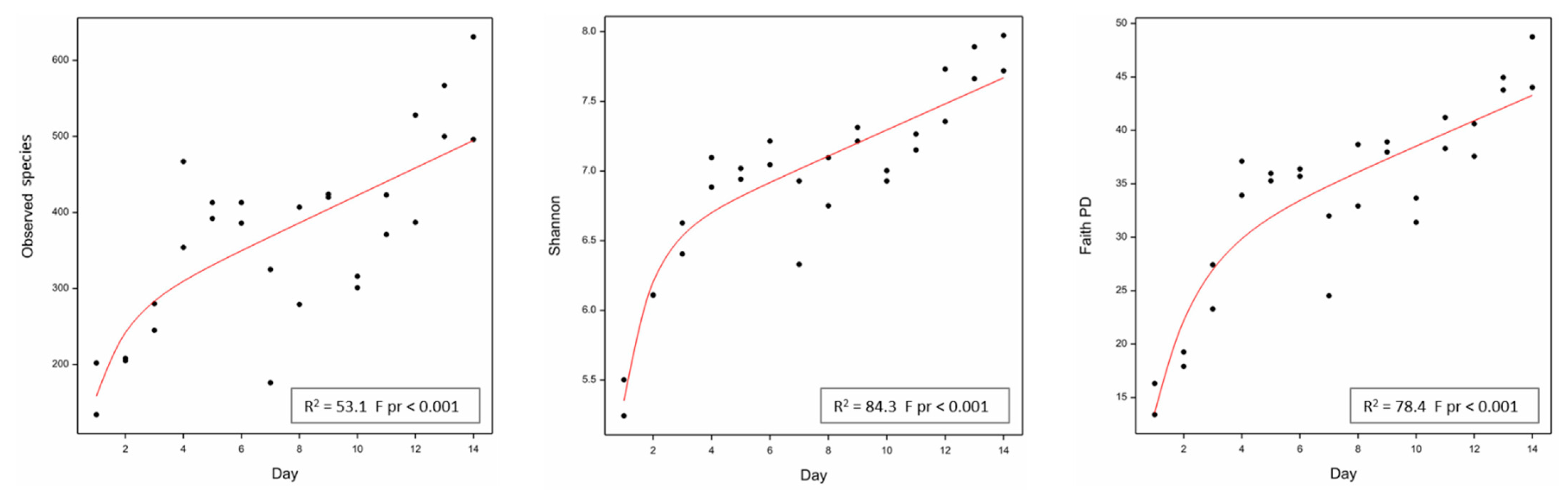
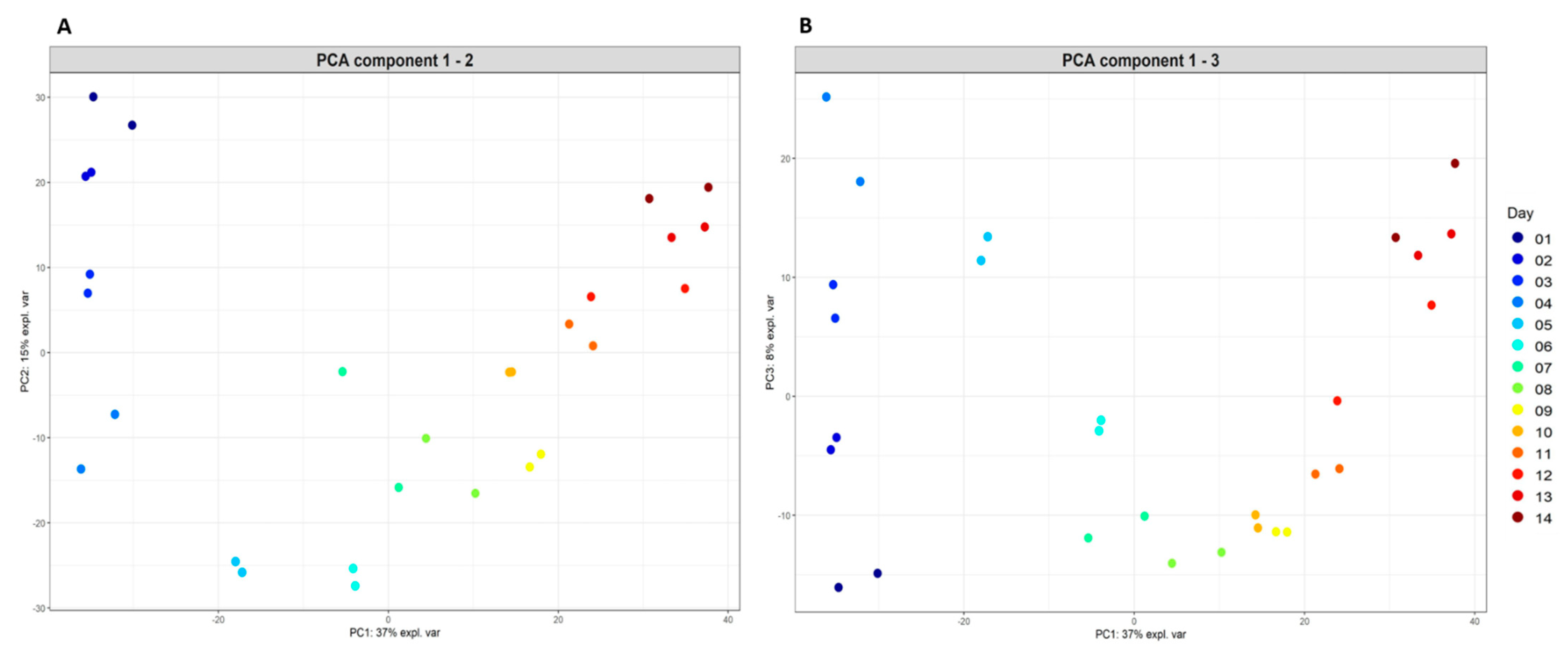
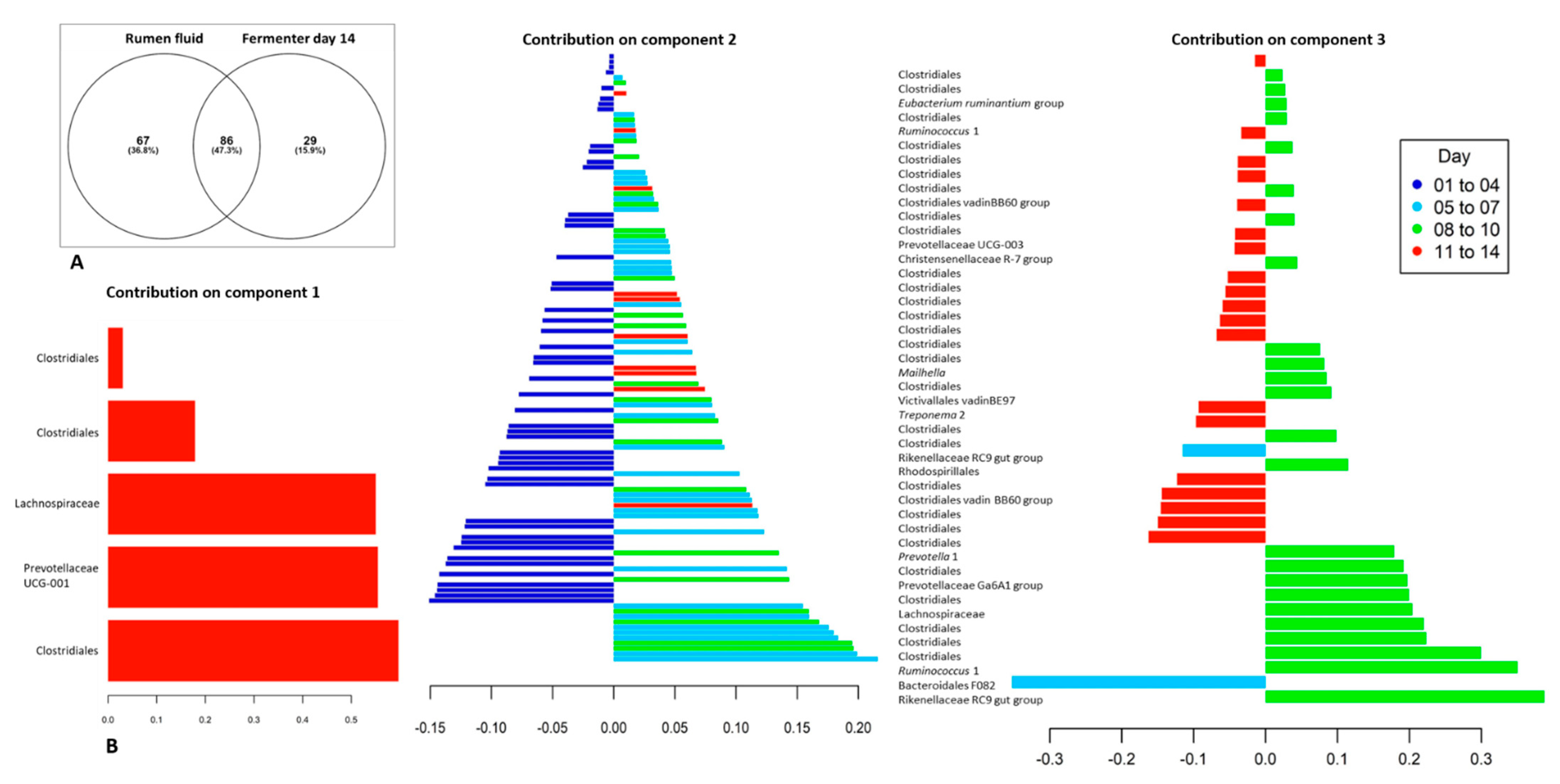
2.2. Fermenter: Microbial Community Diversity
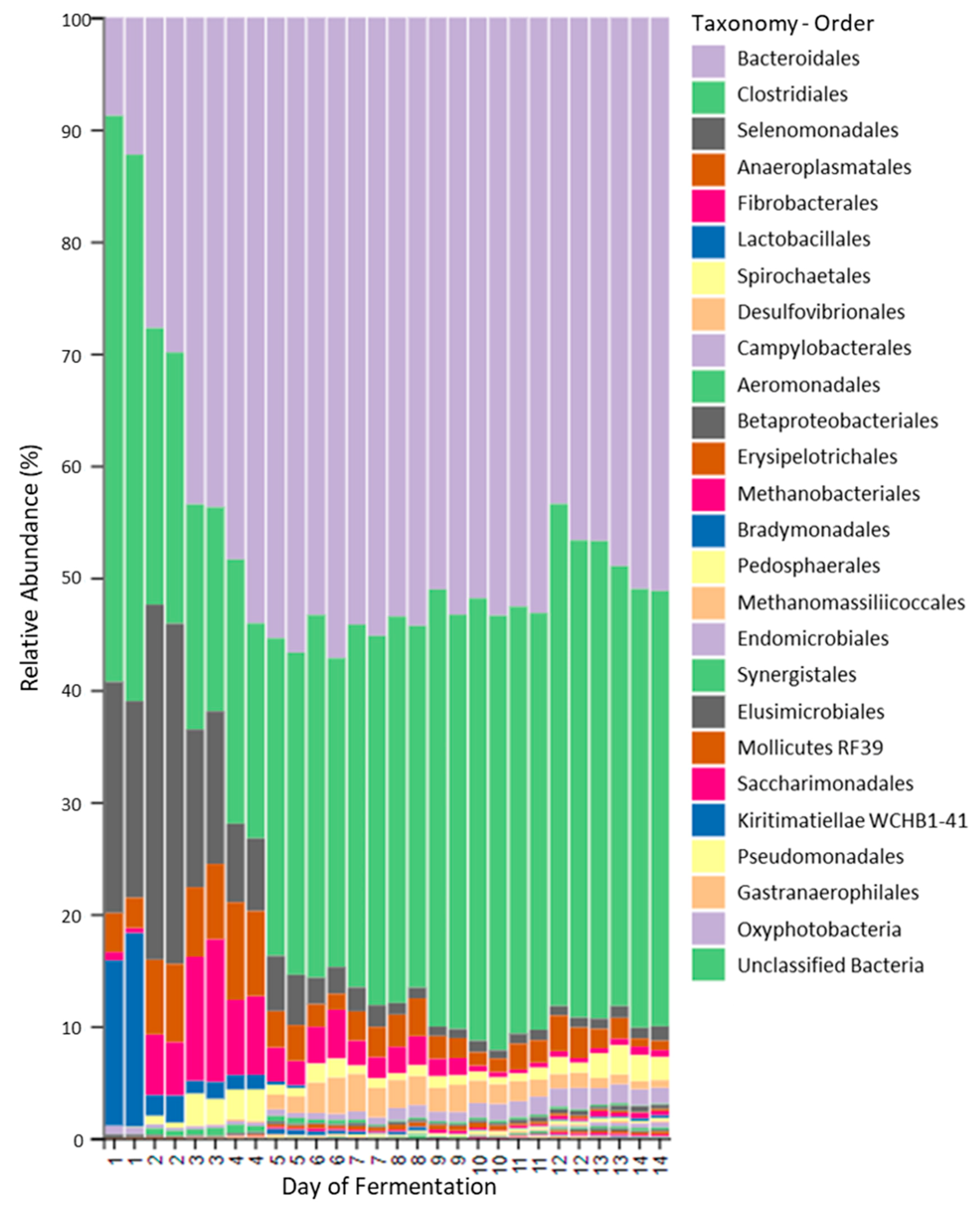
2.3. Fermenter: Microbial Populations
3. Discussion
4. Materials and Methods
4.1. Nutritional Analysis of Indigofera spicata Plant Material and Preparation of Indigofera Extract
4.2. Ethics Approval
4.3. Fermentation Experiment
4.4. Indospicine Degradation Assay
4.5. Chemical Analysis of Indospicine and Related Metabolites in Plant Material, Fermenter and Degradation Assay Samples
4.6. DNA Extractions and Amplicon Generation from Fermenter Samples
4.7. Amplicon Sequence Analysis and Statistics
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fletcher, M.; Al, R.; Cawdell-Smith, A. The occurrence and toxicity of indospicine to grazing animals. Agriculture 2015, 5, 427–440. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.C. Nitro compounds in Indigofera species. Agron. J. 1981, 73, 434–436. [Google Scholar] [CrossRef]
- Emmel, M.W.; Ritchey, G.E. The toxicity of Indigofera endecaphylla Jacq. for Rabbits 1. Agron. J. 1941, 33, 675–677. [Google Scholar] [CrossRef]
- Aylward, J.H.; Court, R.D.; Haydock, K.P.; Strickland, R.W.; Hegarty, M.P. Indigofera species with agronomic potential in the tropics. Rat toxicity studies. Aust. J. Agric. Res. 1987, 38, 177–186. [Google Scholar] [CrossRef]
- Hegarty, M.P.; Pound, A.W. Indospicine, a new hepatotoxic amino-acid from Indigofera spicata. Nature 1968, 217, 354–355. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.T.T.; Materne, C.M.; Silcock, R.G.; D’Arcy, B.R.; Al Jassim, R.; Fletcher, M.T. Seasonal and species variation of the Hepatotoxin indospicine in Australian Indigofera legumes as measured by UPLC-MS/MS. J. Agric. Food. Chem. 2016, 64, 6613–6621. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, M.P.; Pound, A.W. Indospicine, a hepatotoxic amino acid from Indigofera spicata: Isolation, structure, and biological studies. Aust. J. Biol. Sci. 1970, 23, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Tan, E.T.T.; Fletcher, M.T.; Yong, K.W.; D’Arcy, B.R.; Al Jassim, R. Determination of hepatotoxic indospicine in Australian camel meat by ultra-performance liquid chromatography-tandem mass spectrometry. J. Agric. Food. Chem. 2014, 62, 1974–1979. [Google Scholar] [CrossRef]
- Tan, E.T.T.; Al Jassim, R.; Cawdell-Smith, A.J.; Ossedryver, S.M.; D’Arcy, B.R.; Fletcher, M.T. Accumulation, persistence, and effects of indospicine residues in Camels fed Indigofera plant. J. Agric. Food Chem. 2016, 64, 6622–6629. [Google Scholar] [CrossRef]
- Fletcher, M.T.; Reichmann, K.G.; Ossedryver, S.M.; McKenzie, R.A.; Carter, P.D.; Blaney, B.J. Accumulation and depletion of indospicine in calves (Bos taurus) fed creeping indigo (Indigofera spicata). Anim. Prod. Sci. 2018, 58, 568–576. [Google Scholar] [CrossRef]
- FitzGerald, L.M.; Fletcher, M.T.; Paul, A.E.; Mansfield, C.S.; O’Hara, A.J. Hepatotoxicosis in dogs consuming a diet of camel meat contaminated with indospicine. Aust. Vet. J. 2011, 89, 95–100. [Google Scholar] [CrossRef]
- Hegarty, M.P.; Kelly, W.R.; McEwan, D.; Williams, O.J.; Cameron, R. Hepatotoxicity to dogs of horse meat contaminated with indospicine. Aust. Vet. J. 1988, 65, 337–340. [Google Scholar] [CrossRef]
- Netzel, G.; Palmer, D.G.; Masters, A.M.; Tai, S.Y.; Allen, J.G.; Fletcher, M.T. Assessing the risk of residues of the toxin indospicine in bovine muscle and liver from north-west Australia. Toxicon Off. J. Int. Soc. Toxinol. 2019, 163, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Tan, E.T.T.; Ng, J.C.; Al Jassim, R.; D’Arcy, B.R.; Netzel, G.; Fletcher, M.T. Emerging food safety risk of hepatotoxic indospicine in feral Australian camel meat. Food Cont. 2020, 113, 107205. [Google Scholar] [CrossRef]
- Tan, E.T.T.; Yong, K.W.; Wong, S.H.; D’Arcy, B.R.; Al Jassim, R.; De Voss, J.J.; Fletcher, M.T. Thermo-alkaline treatment as a practical degradation strategy to reduce indospicine contamination in camel meat. J. Agric. Food Chem. 2016, 64, 8447–8453. [Google Scholar] [CrossRef]
- Tan, E.T.T.; Al Jassim, R.; D’Arcy, B.R.; Fletcher, M.T. In vitro biodegradation of hepatotoxic indospicine in Indigofera spicata and its degradation derivatives by camel foregut and cattle rumen fluids. J. Agric. Food Chem. 2017, 65, 7528–7534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netzel, G.; Tan, E.T.T.; Yin, M.; Giles, C.; Yong, K.W.L.; Al Jassim, R.; Fletcher, M.T. Bioaccumulation and distribution of indospicine and its foregut metabolites in camels fed Indigofera spicata. Toxins 2019, 11, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, M.E.; Ranilla, M.J.; Tejido, M.L.; Saro, C.; Carro, M.D. Comparison of fermentation of diets of variable composition and microbial populations in the rumen of sheep and Rusitec fermenters. II. Protozoa population and diversity of bacterial communities1. J. Dairy Sci. 2010, 93, 3699–3712. [Google Scholar] [CrossRef] [Green Version]
- Oss, D.B.; Ribeiro, G.O.; Marcondes, M.I.; Yang, W.; Beauchemin, K.A.; Forster, R.J.; McAllister, T.A. Synergism of cattle and bison inoculum on ruminal fermentation and select bacterial communities in an artificial rumen (Rusitec) fed a barley straw based diet. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.F.O.; Terry, S.A.; Holman, D.B.; Breves, G.; Pereira, L.G.R.; Silva, A.G.M.; Chaves, A.V. Tucumã oil shifted ruminal fermentation, reducing methane production and altering the microbiome but decreased substrate digestibility within a Rusitec fed a mixed hay—concentrate diet. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Hungate, R.E.; Bryant, M.P.; Mah, R.A. The rumen bacteria and protozoa. Ann. Rev. Microbiol. 1964, 18, 131–166. [Google Scholar] [CrossRef] [PubMed]
- Vercoe, P.E.; Makkar, H.; Schlink, A. In Vitro Screening of Plant Resources for Extra-Nutritional Attributes in Ruminants Nuclear and Related Methodologies; Vercoe, P.E., Makkar, H., Schlink, A., Eds.; Springer: Dordrecht, The Netherland; London, UK, 2010. [Google Scholar] [CrossRef]
- Czerkawski, J.W.; Breckenridge, G. Design and development of a long-term rumen simulation technique (Rusitec). Br. J. Nutr. 1977, 38, 371–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newbold, J.R.; Rust, S.R. Effect of asynchronous nitrogen and energy supply on growth of ruminal bacteria in batch culture. J. Anim. Sci. 1992, 70, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.C.; Holman, D.B.; Alexander, T.W.; Durmic, Z.; Vercoe, P.E.; Chaves, A.V. The type of forage substrate preparation included as substrate in a Rusitec system affects the ruminal microbiota and fermentation characteristics. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.J.; Lowry, J.B. Australian goats detoxify the goitrogen 3-hydroxy-4(1H) pyridone (DHP) after rumen infusion from an Indonesian goat. Experientia 1984, 40, 1435–1436. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.J.; Megarrity, R.G. Successful transfer of DHP-degrading bacteria from Hawaiian goats to Australian ruminants to overcome the toxicity of Leucaena. Aust. Vet. J. 1986, 63, 259–262. [Google Scholar] [CrossRef]
- Rincón, M.T.; Allison, M.J.; Michelangeli, F.; De Sanctis, Y.; Domínguez-Bello, M.G. Anaerobic degradation of mimosine-derived hydroxypyridines by cell free extracts of the rumen bacterium Synergistes jonesii. FEMS Microbiol. Ecol. 1998, 27, 127–132. [Google Scholar] [CrossRef]
- Allison, M.J.; Mayberry, W.R.; McSweeney, C.S.; Stahl, D.A. Synergistes jonesii, gen. nov., sp. nov.: A rumen bacterium that degrades toxic pyridinediols. Syst. Appl. Microbiol. 1992, 15, 522–529. [Google Scholar] [CrossRef]
- Klieve, A.V.; Ouwerkerk, D.; Turner, A.; Roberton, R. The production and storage of a fermentor-grown bacterial culture containing Synergistes jonesii, for protecting cattle against mimosine and 3-hydroxy-4(1H)-pyridone toxicity from feeding on Leucaena leucocephala. Aust. J. Agric. Res. 2002, 53, 1–5. [Google Scholar] [CrossRef]
- Gilbert, R.A.; Ouwerkerk, D.; Klieve, A.V. Modulation of the rumen microbiome. Microbiol. Aust. 2015, 36, 18–21. [Google Scholar] [CrossRef] [Green Version]
- Pass, M.A.; Arab, H.; Pollitt, S.; Hegarty, M.P. Effects of the naturally occurring arginine analogues indospicine and canavanine on nitric oxide mediated functions in aortic endothelium and peritoneal macrophages. Nat. Toxins 1996, 4, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Hummel, J.; SÜDekum, K.H.; Streich, W.J.; Clauss, M. Forage fermentation patterns and their implications for herbivore ingesta retention times. Functional. Ecol. 2006, 20, 989–1002. [Google Scholar] [CrossRef]
- Lechner-Doll, M.; Rutagwenda, T.; Schwartz, H.J.; Schultka, W.; Engelhardt, W.V. Seasonal changes of ingesta mean retention time and forestomach fluid volume in indigenous camels, cattle, sheep and goats grazing a thornbush savannah pasture in Kenya. J. Agric. Sci. 1990, 115, 409–420. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef]
- Russell, J.B.; Rychlik, J.L. Factors that alter rumen microbial ecology. Science 2001, 292, 1119–1122. [Google Scholar] [CrossRef]
- McLoughlin, S.; Spillane, C.; Claffey, N.; Smith, P.E.; O’Rourke, T.; Diskin, M.G.; Waters, S.M. Rumen microbiome composition is altered in sheep divergent in feed efficiency. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Wallace, R.J.; Sasson, G.; Garnsworthy, P.C.; Tapio, I.; Gregson, E.; Bani, P.; Huhtanen, P.; Bayat, A.R.; Strozzi, F.; Biscarini, F.; et al. A heritable subset of the core rumen microbiome dictates dairy cow productivity and emissions. Sci. Adv. 2019, 5, eaav8391. [Google Scholar] [CrossRef] [Green Version]
- Deusch, S.; Camarinha-Silva, A.; Conrad, J.; Beifuss, U.; Rodehutscord, M.; Seifert, J. A Structural and functional elucidation of the rumen microbiome influenced by various diets and microenvironments. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Holman, D.B.; Gzyl, K.E. A meta-analysis of the bovine gastrointestinal tract microbiota. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef]
- Graf, J. The Family Rikenellaceae. In The Prokaryotes: Other Major Lineages of Bacteria and the Archaea; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 857–859. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D. Challenges in Extrapolating in vitro Findings to in vivo Evaluation of Plant Resources. In In Vitro Screening of Plant Resources for Extra-Nutritional Attributes in Ruminants: Nuclear and Related Methodologies; Vercoe, P.E., Makkar, H.P.S., Schlink, A.C., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 233–242. [Google Scholar] [CrossRef]
- Zehavi, T.; Probst, M.; Mizrahi, I. Insights into culturomics of the rumen microbiome. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Seshadri, R.; Leahy, S.C.; Attwood, G.T.; Teh, K.H.; Lambie, S.C.; Cookson, A.L.; Eloe-Fadrosh, E.A.; Pavlopoulos, G.A.; Hadjithomas, M.; Varghese, N.J.; et al. Cultivation and sequencing of rumen microbiome members from the Hungate1000 Collection. Nat. Biotech. 2018, 36, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Creevey, C.J.; Kelly, W.J.; Henderson, G.; Leahy, S.C. Determining the culturability of the rumen bacterial microbiome. Microb. Biotech. 2014, 7, 467–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, N.D.; Newbold, C.J.; Wallace, R.J. Nitrogen metabolism in the rumen. In Nitrogen and Phosphorus Nutrition of Cattle: Reducing the Environmental Impact of Cattle Operations; Pfeffer, E., Hristov, A.N., Eds.; CABI Publishing: Cambridge, MA, USA, 2005; pp. 71–115. [Google Scholar] [CrossRef]
- Hartinger, T.; Gresner, N.; Sudekum, K.H. Does intra-ruminal nitrogen recycling waste valuable resources? A review of major players and their manipulation. J. Anim. Sci. Biotechnol. 2018, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firkins, J.L. Rumen protein breakdown and ammonia assimilation. In Improving Rumen Function; McSweeney, C.S., Mackie, R.I., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK; Philadelphia, PA, USA, 2020; Volume 83, pp. 383–420. [Google Scholar] [CrossRef]
- McSweeny, C.S.; Allison, M.J.; Mackie, R.I. Amino acid utilization by the ruminal bacterium Synergistes jonesii strain 78-1. Arch. Microbiol. 1993, 159, 131–135. [Google Scholar] [CrossRef]
- Cunin, R.; Glansdorff, N.; Pierard, A.; Stalon, V. Biosynthesis and metabolism of arginine in bacteria. Microbiol. Mol. Biol. Rev. 1986, 50, 314. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesinger, T.; Haas, D.; Hegarty, M.P. Indospicine as an arginine antagonist in Escherichia coli and Pseudomonas aeruginosa. Biochim. Biophys. Acta 1972, 262, 214–219. [Google Scholar] [CrossRef]
- Madsen, N.P.; Hegarty, M.P. Inhibition of rat liver homogenate arginase activity in vitro by the hepatotoxic amino acid indospicine. Biochem. Pharmacol. 1970, 19, 2391–2393. [Google Scholar] [CrossRef]
- Hungate, R.E. A roll-tube method for the cultivation of strict anaerobes. In Methods in Microbiology; Norris, J.R., Ribbons, D.W., Eds.; Academic Press: London, UK, 1969; pp. 117–132. [Google Scholar]
- McSweeney, C.S.; Denman, S.E.; Mackie, R.I. Rumen bacteria. In Methods in Gut Microbial Ecology for Ruminants; Makkar, H.S., McSweeney, C., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 23–37. [Google Scholar] [CrossRef]
- Klieve, A.V.; Hudman, J.F.; Bauchop, T. Inducible bacteriophages from ruminal bacteria. Appl. Environ. Microbiol. 1989, 55, 1630–1634. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.M.; Klieve, A.V.; Plumb, J.J.; Aisthorpe, R.; Blackall, L.L. An in vitro cultured rumen inoculum improves nitrogen digestion in mulga-fed sheep. Aust. J. Agric. Res. 1997, 48, 403–409. [Google Scholar] [CrossRef]
- Lang, C.S.; Wong, S.H.; Chow, S.; Challinor, V.L.; Yong, K.W.; Fletcher, M.T.; Arthur, D.M.; Ng, J.C.; De Voss, J.J. Synthesis of l-indospicine, [5,5,6-(2)H3]-l-indospicine and l-norindospicine. Org. Biomol. Chem. 2016, 14, 6826–6832. [Google Scholar] [CrossRef]
- Yu, Z.; Forster, R.J. Nucleic acid extraction, oligonucleotide probes and PCR methods. In Methods in Gut Microbial Ecology for Ruminants; Makkar, H.P.S., McSweeney, C.S., Eds.; International Atomic Energy Agency; Springer Academic Press: Dordrecht, The Netherlands, 2005; pp. 81–104. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformation 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Meth. 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Meth. 2016, 13, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glockner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohart, F.; Gautier, B.; Singh, A.; Le Cao, K.A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]
- Jolliffe, I.T.; Cadima, J. Principal component analysis: A review and recent developments. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef]
- Shen, H.; Huang, J.Z. Sparse principal component analysis via regularized low rank matrix approximation. J. Multivar. Anal. 2008, 99, 1015–1034. [Google Scholar] [CrossRef] [Green Version]
- Le Cao, K.A.; Costello, M.E.; Lakis, V.A.; Bartolo, F.; Chua, X.Y.; Brazeilles, R.; Rondeau, P. MixMC: A multivariate statistical framework to gain insight into microbial communities. PLoS ONE 2016, 11, e0160169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Cao, K.A.; Boitard, S.; Besse, P. Sparse PLS discriminant analysis: Biologically relevant feature selection and graphical displays for multiclass problems. BMC Bioinform. 2011, 12, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutritional Analysis | Concentration (g/100 g or % w/w) |
|---|---|
| Dry Matter | 92.4% |
| Ash | 14.50 |
| Crude Fibre | 23.60% |
| Dietary Fibre (total) | 50.60 |
| Fat | 4.70 |
| Nitrogen | 3.4% |
| Protein (N % × 6.25) | 21.25% |
| Sodium | 0.025 |
| Phosphorus | 0.27 |
| Calcium | 2.40 |
| Potassium | 1.99 |
| Sulphur | 0.308 |
| Fermentation Days 8 to 10 | Fermentation Days 11 to 14 | ||
|---|---|---|---|
| Highly abundant core bacteria * | Relative abundance (%) ** | Highly abundant core bacteria | Relative abundance (%) |
| Rikenellaceae RC9 gut group | 20.84 | Rikenellaceae RC9 gut group | 15.65 |
| Prevotella 1 | 11.44 | Bacteroidales F082 (uncultured rumen bacterium) | 12.39 |
| Bacteroidales F082 (uncultured rumen bacterium) | 10.80 | Prevotella 1 | 10.90 |
| Ruminococcus 1 | 5.68 | Butyrivibrio 2 | 6.51 |
| Prevotellaceae Ga6A1 group | 4.75 | Ruminococcus 1 | 5.70 |
| Lachnospiraceae | 4.36 | Lachnoclostridium | 3.76 |
| Butyrivibrio 2 | 4.27 | Lachnospiraceae | 3.07 |
| Lachnoclostridium | 3.14 | Clostridiales | 2.66 |
| Clostridiales | 2.99 | Prevotellaceae Ga6A1 group | 2.17 |
| Desulfovibrio | 2.26 | Eubacterium coprostanoligenes group | 2.17 |
| Anaeroplasma | 2.15 | Christensenellaceae R-7 group | 1.89 |
| Ruminococcaceae NK4A214 group | 1.82 | Anaeroplasma | 1.87 |
| Fibrobacter | 1.60 | Bacteroidales RF16 group | 1.85 |
| Lachnospiraceae | 1.51 | Ruminococcaceae NK4A214 group | 1.81 |
| Eubacterium coprostanoligenes group | 1.19 | Treponema 2 | 1.76 |
| Campylobacter | 1.10 | Campylobacter | 1.58 |
| Christensenellaceae R-7 group | 1.10 | Prevotellaceae UCG-003 | 1.50 |
| Lachnospiraceae UCG-009 | 0.99 | Eubacterium ruminantium group | 1.45 |
| Eubacterium ruminantium group | 0.91 | Ruminococcaceae UCG-014 | 1.14 |
| Bacteroidales RF16 group | 0.87 | Desulfovibrio | 1.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilbert, R.A.; Netzel, G.; Chandra, K.; Ouwerkerk, D.; Fletcher, M.T. Degradation of the Indospicine Toxin from Indigofera spicata by a Mixed Population of Rumen Bacteria. Toxins 2021, 13, 389. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060389
Gilbert RA, Netzel G, Chandra K, Ouwerkerk D, Fletcher MT. Degradation of the Indospicine Toxin from Indigofera spicata by a Mixed Population of Rumen Bacteria. Toxins. 2021; 13(6):389. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060389
Chicago/Turabian StyleGilbert, Rosalind A., Gabriele Netzel, Kerri Chandra, Diane Ouwerkerk, and Mary T. Fletcher. 2021. "Degradation of the Indospicine Toxin from Indigofera spicata by a Mixed Population of Rumen Bacteria" Toxins 13, no. 6: 389. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060389