Physiological Effects of Deoxynivalenol from Naturally Contaminated Corn on Cerebral Tryptophan Metabolism, Behavioral Response, Gastrointestinal Immune Status and Health in Pigs Following a Pair-Feeding Model


Abstract
:1. Introduction
2. Results
2.1. Growth Performance
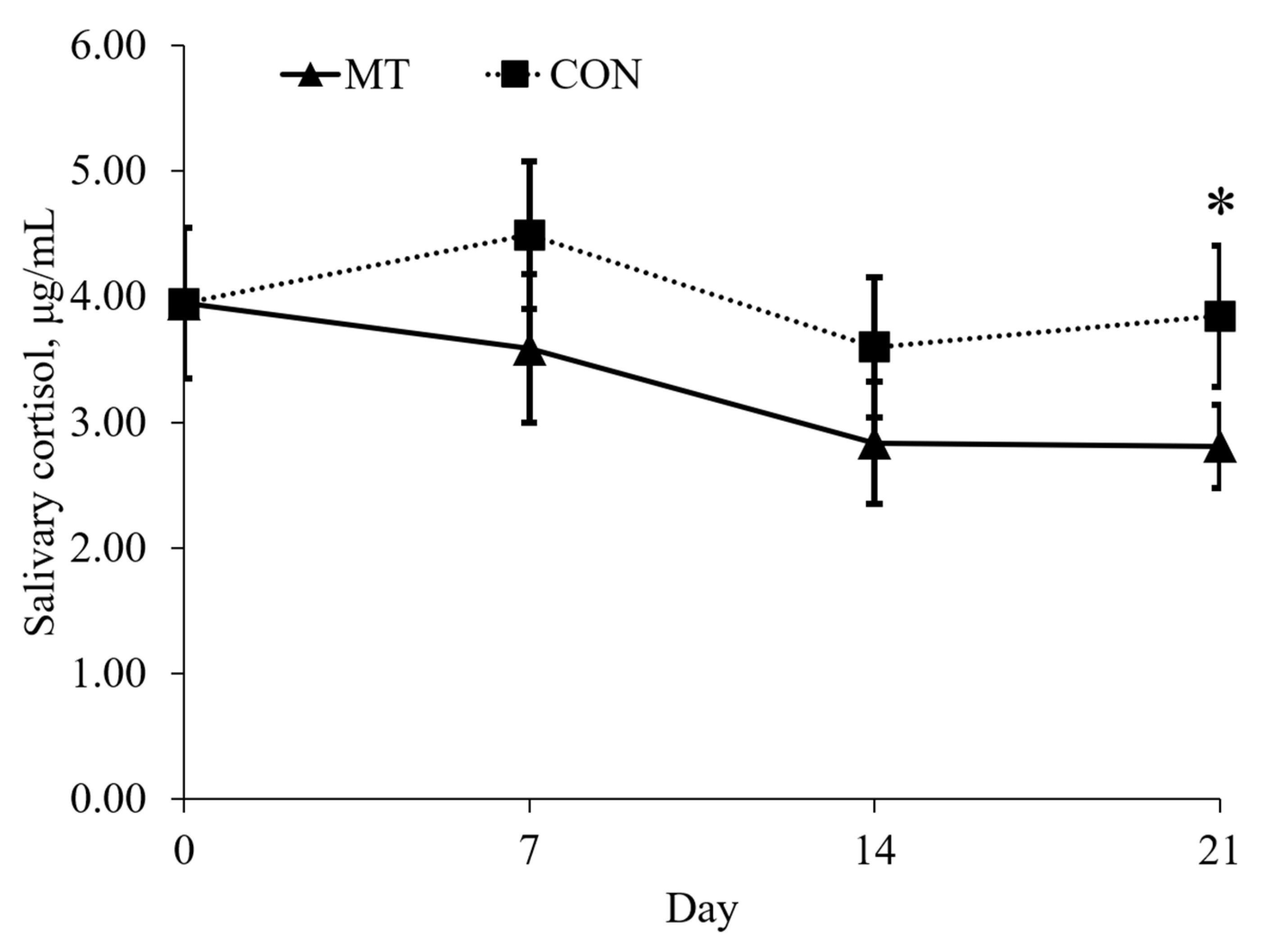
2.2. Physical Activity and Salivary Cortisol
2.3. Immunological and Oxidative Stress Analysis
2.4. Gut Morphology
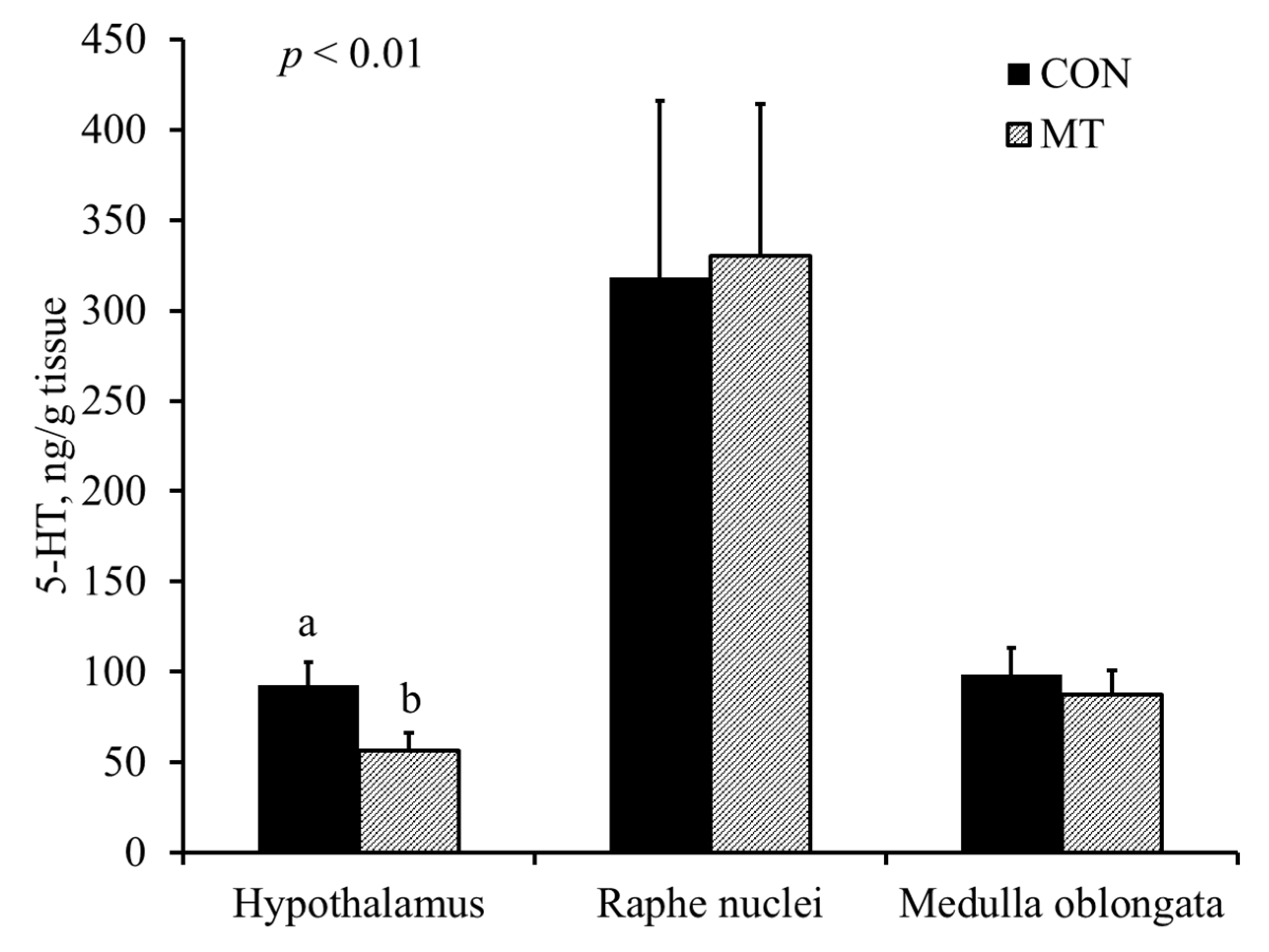
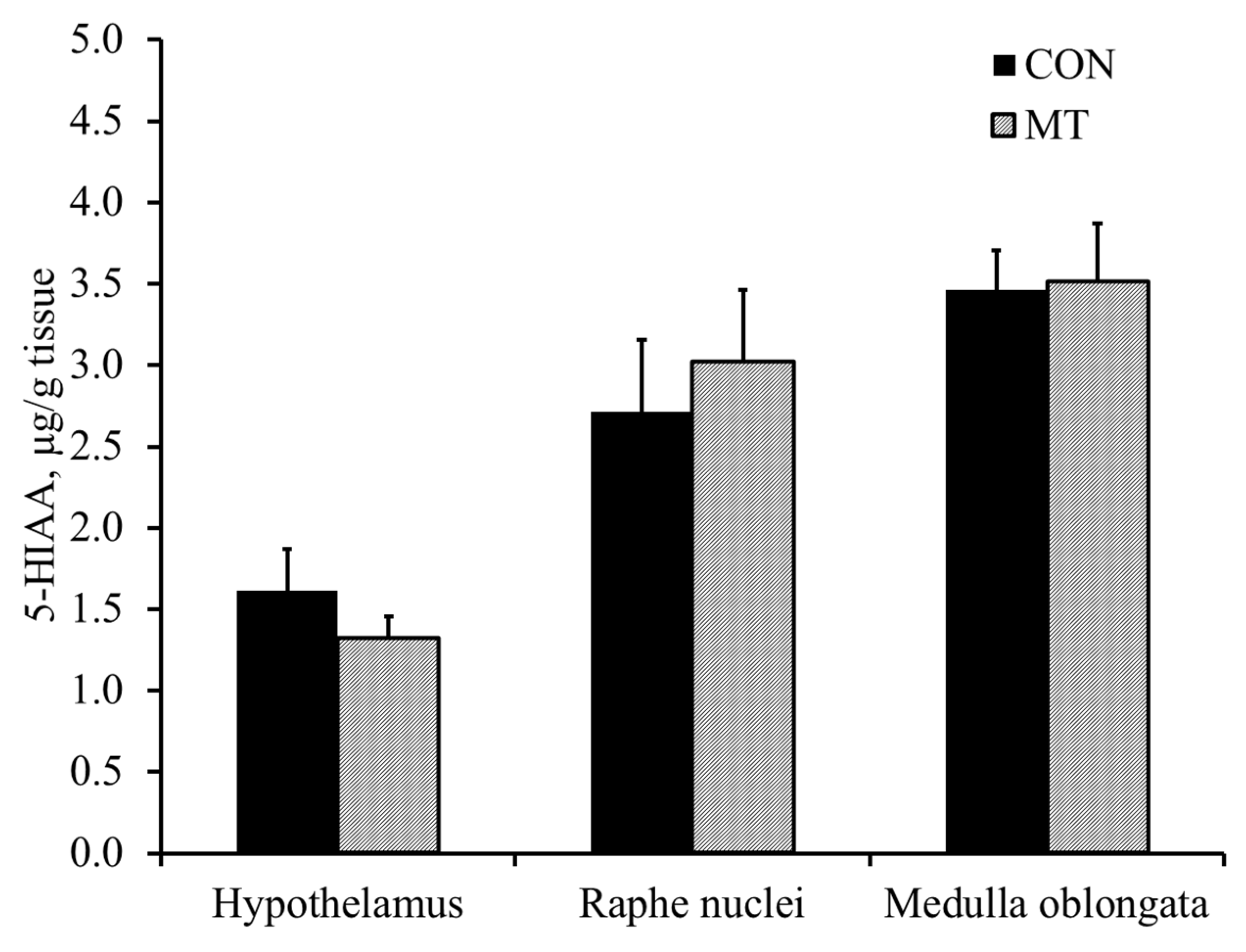
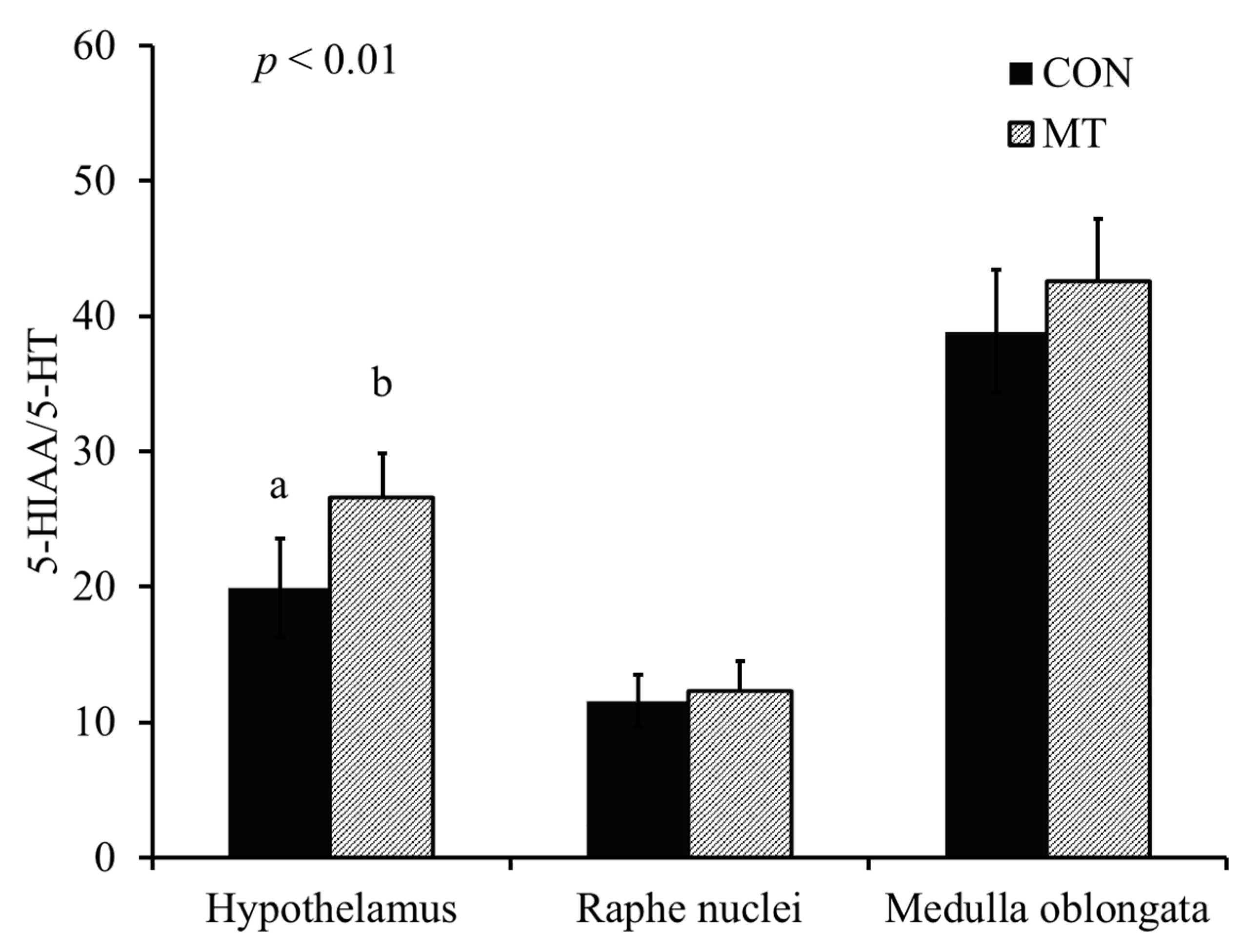
2.5. Hypothalamic Serotonin and 5-Hydroxyindoleacetic Acid
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals and Experimental Diets
5.2. Physical Activity
5.3. Salivary Cortisol
5.4. Immunological and Oxidative Stress Parameters
5.5. Tissue Collection and Analysis
5.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Chaytor, A.C.; Hansen, J.A.; van Heugten, E.; See, M.T.; Kim, S.W. Occurrence and decontamination of mycotoxins in swine feed. Asian-Australas. J. Anim. Sci. 2011, 24, 723–738. [Google Scholar] [CrossRef]
- Holanda, D.M.; Kim, S.W. Mycotoxin occurrence, toxicity, and detoxifying agents in pig production with an emphasis on deoxynivalenol. Toxins 2021, 13, 171. [Google Scholar] [CrossRef]
- Pinton, P.; Braicu, C.; Nougayrede, J.P.; Laffitte, J.; Taranu, I.; Oswald, I.P. Deoxynivalenol impairs porcine intestinal barrier function and decreases the protein expression of claudin-4 through a mitogen-activated protein kinase-dependent mechanism. J. Nutr. 2010, 140, 1956–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and potential effects on humans. J. Toxicol. Environ. Health B Crit. Rev. 2005, 8, 39–69. [Google Scholar] [CrossRef] [PubMed]
- Borutova, R.; Faix, S.; Placha, I.; Gresakova, L.; Cobanova, K.; Leng, L. Effects of deoxynivalenol and zearalenone on oxidative stress and blook phagocytic activity in broilers. Arch. Anim. Nutr. 2008, 62, 303–312. [Google Scholar] [CrossRef]
- Goyarts, T.; Dänicke, S.; Rothkötter, H.-J.; Spilke, J.; Tiemann, U.; Schollenberger, M. On the effects of a chronic deoxynivalenol intoxication on performance, haematological and serum parameters of pigs when diets are offered either for ad libitum or fed restrictively. J. Vet. Med. 2005, 52, 305–314. [Google Scholar] [CrossRef]
- Prelusky, D.B.; Yeung, J.M.; Thompson, B.K.; Trenholm, H.L. Effect of deoxynivalenol on neurotransmitters in discrete regions of swine brain. Arch. Environ. Contam. Toxicol. 1992, 22, 36–40. [Google Scholar] [CrossRef]
- Prelusky, D.B. The effect of low-level deoxynivalenol on neurotransmitter levels measured in pig cerebral spinal fluid. J. Environ. Sci. Health B 1993, 28, 731–761. [Google Scholar] [CrossRef]
- Rotter, B.A.; Prelusky, D.B.; Pestka, J.J. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Swamy, H.V.L.N.; Smith, T.K.; MacDonald, E.J.; Boermans, H.J.; Squires, E.J. Effects of feeding a blend of grains naturally contaminated with Fusarium mycotoxins on swine performance, brain regional neurochemistry, and serum chemistry and the efficacy of a polymeric glucomannan mycotoxin adsorbent. J. Anim. Sci. 2002, 80, 3257–3267. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yin, J.; Li, D.; Zhou, X.; Li, X. Tryptophan enhances ghrelin expression and secretion associated with increased food intake and weight gain in weanling pigs. Domest. Anim. Endocrinol. 2007, 33, 47–61. [Google Scholar] [CrossRef]
- Fitzpatrick, D.W.; Boyd, K.E.; Watts, B.M. Comparison of the trichothecenes deoxynivalenol and T-2 toxin for their effects on brain biogenic monoamines in the rat. Toxicol. Lett. 1988, 40, 241–245. [Google Scholar] [CrossRef]
- Fitzpatrick, D.W.; Boyd, K.E.; Wilson, L.M.; Wilson, J.R. Effect of the trichothecene deoxynivalenol on brain biogenic monoamines concentrations in rats and chickens. J. Environ. Sci. Health B 1988, 23, 159–170. [Google Scholar] [CrossRef]
- Shea, M.M.; Mench, J.A.; Thomas, O.P. The effect of dietary tryptophan on aggressive behavior in developing and mature broiler breeder males. Poult. Sci. 1990, 69, 1664–1669. [Google Scholar] [CrossRef] [PubMed]
- Raleigh, M.J.; McGuire, M.T.; Brammer, G.L.; Pollack, D.B.; Yuwiler, A. Serotonergic mechanisms promote dominance acquisition in adult male vervet monkeys. Brain Res. 1991, 559, 181–190. [Google Scholar] [CrossRef]
- Li, Y.Z.; Kerr, B.J.; Kidd, M.T.; Gonyou, H.W. Use of supplementary tryptophan to modify the behavior of pigs. J. Anim. Sci. 2006, 84, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.B.; Voilque, G.; Kim, J.D.; Odle, J.; Kim, S.W. Effects of increasing tryptophan intake on growth and physiological changes in nursery pigs. J. Anim. Sci. 2012, 90, 2264–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meunier-Salaun, M.C.; Monnier, M.; Colleaux, Y.; Seve, B.; Henry, Y. Impact of dietary tryptophan and behavioral type on behavior, plasma cortisol, and brain metabolites of young pigs. J. Anim. Sci. 1991, 69, 3689–3698. [Google Scholar] [CrossRef] [PubMed]
- Boyd, K.E.; Fitzpatrick, D.W.; Wilson, J.R.; Wilson, L.M. Effect of T-2 toxin on brain biogenic monoamines in rats and chickens. Can. J. Vet. Res. 1988, 52, 181–185. [Google Scholar]
- Shen, Y.B.; Voilque, G.; Odle, J.; Kim, S.W. Dietary L-tryptophan supplementation with reduced large neutral amino acids enhances feed efficiency and decreases stress hormone secretion in nursery pigs under social-mixing stress. J. Nutr. 2012, 142, 1540–1546. [Google Scholar] [CrossRef] [Green Version]
- Prelusky, D.B. The effect of deoxynivalenol on serotoninergic neurotransmitter levels in pig blood. J. Environ. Sci. Health B 1994, 29, 1203–1218. [Google Scholar] [CrossRef]
- Hornung, J.P. The human raphe nuclei and the serotonergic system. J. Chem. Neuroanat. 2003, 26, 331–343. [Google Scholar] [CrossRef]
- Stahl, S.M. Mechanism of action of serotonin selective reuptake inhibitors. Serotonin receptors and pathways mediate therapeutic effects and side effects. J. Affect. Disord. 1998, 51, 215–235. [Google Scholar] [CrossRef]
- Gregory, R.E.; Ettinger, D.S. 5-HT3 receptor antagonists for the prevention of chemotherapy-induced nausea and vomiting. A comparison of their pharmacology and clinical efficacy. Drugs 1998, 55, 173–189. [Google Scholar] [CrossRef]
- Markus, C.R.; Panhuysen, G.; Tuiten, A.; Koppeschaar, H.; Fekkes, D.; Peters, M.L. Does carbohydrate-rich, protein-poor food prevent a deterioration of mood and cognitive performance of stress-prone subjects when subjected to a stressful task? Appetite 1998, 31, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, S.J.; Ruis, M.; Dekker, R.; van Diepen, H.; Korte, M.; Mroz, Z. Surplus dietary tryptophan reduces plasma cortisol and noradrenaline concentrations and enhances recovery after social stress in pigs. Physiol. Behav. 2005, 85, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, S.J.; Guzik, A.C.; van der Meulen, J.; Dekker, R.; Kogut, J.; Kerr, B.J.; Southern, L.L. Effects of supplemental L-tryptophan on serotonin, cortisol, intestinal integrity, and behavior in weanling piglets. J. Anim. Sci. 2006, 84, 963–971. [Google Scholar] [CrossRef]
- Young, S.N.; Leyton, M. The role of serotonin in human mood and social interaction. Insight from altered tryptophan levels. Pharmacol. Biochem. Behav. 2002, 71, 857–865. [Google Scholar] [CrossRef]
- Young, L.G.; McGirr, L.; Valli, V.E.; Lumsden, J.H.; Lun, A. Vomitoxin in corn fed to young pigs. J. Anim. Sci. 1983, 57, 655–664. [Google Scholar] [CrossRef]
- Williams, K.C.; Blaney, B.J.; Magee, M.H. Responses of pigs fed wheat naturally infected with fusarium graminearum and containing the mycotoxins 4-deoxynivalenol and zearalenone. Aust. J. Agric. Res. 1988, 39, 1095–1105. [Google Scholar] [CrossRef]
- Weaver, A.C.; See, M.T.; Kim, S.W. Protective effect of two yeast based feed additives on pigs chronically exposed to deoxynivalenol and zearalenone. Toxins 2014, 6, 3336–3353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holanda, D.M.; Yiannikouris, A.; Kim, S.W. Investigation of the efficacy of a postbiotic yeast cell wall-based blend on newly-weaned pigs under a dietary challenge of multiple mycotoxins with emphasis on deoxynivalenol. Toxins 2020, 12, 504. [Google Scholar] [CrossRef]
- Rotter, M.A.; Thompson, B.K.; Lessard, M.; Trenholm, H.L.; Tryphonas, H. Influence of low-level exposure to Fusarium mycotoxins on selected immunological and hematological parameters in young swine. Fund. Appl. Toxicol. 1994, 23, 117–124. [Google Scholar] [CrossRef]
- Kim, S.W.; Holanda, D.M.; Gao, X.; Park, I.K.; Yiannikouris, A. Efficacy of a yeast cell wall extract to mitigate the effect of naturally co-occurring mycotoxins contaminating feed ingredients fed to young pigs: Impact on gut health, microbiome, and growth. Toxins 2019, 11, 633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, A.C.; See, M.T.; Hansen, J.A.; Kendall, D.C.; Kim, Y.B.; de Souza, A.L.P.; Middleton, T.F.; Kim, S.W. Effects of feed additives for pigs under chronic exposure to aflatoxin and deoxynivalenol on their growth, organ health, and immune status. Toxins 2013, 5, 1261–1281. [Google Scholar] [CrossRef] [Green Version]
- Swamy, H.V.L.N.; Smith, T.K.; MacDonald, E.J.; Karrow, N.A.; Woodward, B.; Boermans, H.J. Effects of feeding a blend of grains naturally contaminated with Fusarium mycotoxins on growth and immunological measurements of starter pigs, and the efficacy of a polymeric glucomannan mycotoxin adsorbent. J. Anim. Sci. 2003, 81, 2792–2803. [Google Scholar] [CrossRef] [PubMed]
- Chaytor, A.C.; See, M.T.; Hansen, J.A.; de Souza, A.L.P.; Middleton, T.F.; Kim, S.W. Effects of chronic exposure of diets with low levels of aflatoxin and deoxynivalenol on growth and immune status of pigs. J. Anim. Sci. 2011, 89, 124–135. [Google Scholar] [CrossRef]
- Weaver, A.C.; Campbell, J.M.; Crenshaw, J.D.; Polo, J.; Kim, S.W. Efficacy of dietary spray dried plasma protein to mitigate the negative effects on performance of pigs fed diets with mycotoxin contaminated corn. J. Anim. Sci. 2014, 92, 3878–3886. [Google Scholar] [CrossRef]
- Lessard, M.; Savard, C.; Deschene, K.; Lauzon, K.; Pinilla, V.A.; Gagnon, C.A.; Lapointe, J.; Guay, F.; Chorfi, Y. Impact of deoxynivalenol (DON) contaminated feed on intestinal integrity and immune response in swine. Food Chem. Toxicol. 2015, 80, 7–16. [Google Scholar] [CrossRef]
- Cheat, S.; Pinton, P.; Cossalter, A.-M.; Cognie, J.; Vilarino, M.; Callu, P.; Raymond-Letron, I.; Oswald, I.P.; Kolf-Clauw, M. The mycotoxins deoxynivalenol and nivalenol show in vivo synergism on jejunum enterocytes apoptosis. Food Chem. Toxicol. 2016, 87, 45–54. [Google Scholar] [CrossRef]
- Avantaggiato, G.; Havenaar, R.; Visconti, A. Evaluation of the intestinal absorption of deoxynivalenol and nivalenol by an in vitro gastrointestinal model, and the binding efficacy of activated carbon and other adsorbent materials. Food Chem. Toxicol. 2004, 42, 817–824. [Google Scholar] [CrossRef]
- Weaver, A.C.; King, W.D.; Verax, M.; Fox, U.; Kudupoje, M.B.; Mathis, G.; Lumpkins, B.; Yiannikouris, A. Impact of chronic levels of naturally multi-contaminated feed with Fusarium mycotoxins on broiler chickens and evaluation of the mitigation properties of different titers of yeast cell wall extract. Toxins 2020, 12, 636. [Google Scholar] [CrossRef] [PubMed]
- Girish, C.K.; Smith, T.K. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on small intestinal morphology of turkeys. Poult. Sci. 2008, 87, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, T.B.; Slate, A.B.; Johansson, A.S. Sampling feeds for mycotoxin analysis. In The Mycotoxin Blue Book, Diaz, D.E., Ed.; Nottingham University Press: Bath, UK, 2005; pp. 1–23. [Google Scholar]
- Shen, Y.B.; Coffey, M.T.; Kim, S.W. Effects of short term supplementation of L-tryptophan and reducing large neutral amino acid along with L-tryptophan supplementation on growth and stress response in pigs. Anim. Feed Sci. Technol. 2015, 207, 245–252. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, S.; Kim, S.W. Effects of supplemental xylanase on gut health and growth performance of nursery pigs fed diets with corn distillers’ dried grains with solubles. J. Anim. Sci. 2020, 98, skaa185. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.E.; Tyus, J.; Kim, S.W. Synbiotic effects of enzyme and probiotics on intestinal health and growth of newly-weaned pigs challenged with enterotoxigenic F18+ coli. Front. Vet. Sci. 2020, 7, 573. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Flowers, W.L.; Saraiva, A.; Yeum, K.-J.; Kim, S.W. Effect of social ranks and gestation housing systems on oxidative stress status, reproductive performance, and immune status of sows. J. Anim. Sci. 2013, 91, 5848–5858. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Kim, S.W. Oxidative stress status and reproductive performance of sows during gestation and lactation under different thermal environments. Asian-Australas. J. Anim. Sci. 2020, 33, 722–731. [Google Scholar] [CrossRef]
- Shen, Y.B.; Weaver, A.C.; Kim, S.W. Effect of feed grade L-methionine on growth performance and gut health in nursery pigs compared with conventional DL-methionine. J. Anim. Sci. 2014, 92, 5530–5539. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | ||||
|---|---|---|---|---|
| Item | CON | MT | SEM | p-Value |
| Initial body weight, kg | 25.6 | 25.5 | 0.4 | 0.938 |
| Average daily gain, g/d | ||||
| day 0 to 7 | 338 | 523 | 38 | 0.011 |
| day 7 to 14 | 745 | 751 | 32 | 0.904 |
| day 14 to 21 | 605 | 596 | 26 | 0.808 |
| day 0 to 21 | 563 | 623 | 14 | 0.017 |
| Average daily feed intake 1, g/d | ||||
| day 0 to 7 | 1266 | 1296 | 19 | 0.299 |
| day 7 to 14 | 1614 | 1646 | 23 | 0.351 |
| day 14 to 21 | 1508 | 1546 | 20 | 0.209 |
| day 0 to 21 | 1463 | 1496 | 17 | 0.208 |
| Gain to feed ratio | ||||
| day 0 to 7 | 0.27 | 0.40 | 0.03 | 0.014 |
| day 7 to 14 | 0.46 | 0.46 | 0.01 | 0.882 |
| day 14 to 21 | 0.40 | 0.39 | 0.02 | 0.595 |
| day 0 to 21 | 0.39 | 0.42 | 0.01 | 0.062 |
| Treatment | ||||
|---|---|---|---|---|
| Item | CON | MT | SEM | p-Value |
| day 7 | ||||
| IgA 1, µg/mL | 2341 | 2588 | 138 | 0.247 |
| IgG 2, mg/mL | 118 | 78 | 16 | 0.126 |
| IgM 3, µg/mL | 877 | 872 | 42 | 0.946 |
| TNF-α 4, pg/mL | 94.7 | 90.9 | 4.7 | 0.581 |
| MDA 5, µM | 13.1 | 13.1 | 1.7 | 0.991 |
| day 21 | ||||
| IgA, µg/mL | 2555 | 2510 | 318 | 0.923 |
| IgG, mg/mL | 96 | 106 | 23 | 0.755 |
| IgM, µg/mL | 1250 | 1374 | 86 | 0.340 |
| TNF-α, pg/mL | 79.7 | 93.5 | 11.8 | 0.436 |
| MDA, µM | 12.2 | 10.7 | 1.0 | 0.304 |
| Treatment | ||||
|---|---|---|---|---|
| Item | CON | MT | SEM | p-Value |
| IgA 1, mg/g protein | ||||
| Stomach | 27.51 | 26.66 | 3.00 | 0.846 |
| Duodenum | 39.64 | 43.25 | 5.14 | 0.635 |
| Jejunum | 20.71 | 21.84 | 1.98 | 0.700 |
| Ileum | 34.60 | 33.37 | 1.77 | 0.640 |
| IgG 2, mg/g protein | ||||
| Stomach | 53.87 | 48.42 | 11.55 | 0.749 |
| Duodenum | 59.13 | 78.38 | 12.26 | 0.304 |
| Jejunum | 36.62 | 37.58 | 4.92 | 0.894 |
| Ileum | 59.19 | 69.47 | 7.23 | 0.349 |
| IgM 3, mg/g protein | ||||
| Stomach | 15.56 | 14.55 | 1.45 | 0.637 |
| Duodenum | 9.34 | 7.92 | 1.63 | 0.557 |
| Jejunum | 9.11 | 11.05 | 1.12 | 0.260 |
| Ileum | 7.40 | 7.33 | 0.53 | 0.926 |
| TNF-α 4, pg/g protein | ||||
| Stomach | 709 | 685 | 45 | 0.720 |
| Duodenum | 1227 | 986 | 93 | 0.112 |
| Jejunum | 716 | 1038 | 37 | 0.004 |
| Ileum | 2737 | 2946 | 333 | 0.671 |
| Liver | 863 | 756 | 113 | 0.524 |
| Lung | 2424 | 1786 | 244 | 0.107 |
| Lymph node | 1908 | 1709 | 185 | 0.477 |
| Treatment | ||||
|---|---|---|---|---|
| Item | CON | MT | SEM | p-Value |
| Stomach | ||||
| Depth of gastric pits, µm | 241 | 175 | 12 | 0.006 |
| Duodenum | ||||
| Villus height, µm | 626 | 576 | 37 | 0.365 |
| Crypt depth, µm | 550 | 555 | 17 | 0.845 |
| Villus to crypt ratio | 1.15 | 1.04 | 0.08 | 0.330 |
| Jejunum | ||||
| Villus height, µm | 553 | 459 | 20 | 0.013 |
| Crypt depth, µm | 355 | 423 | 17 | 0.026 |
| Villus to crypt ratio | 1.58 | 1.09 | 0.05 | < 0.001 |
| Ileum | ||||
| Villus height, µm | 525 | 421 | 34 | 0.070 |
| Crypt depth, µm | 345 | 383 | 19 | 0.193 |
| Villus to crypt ratio | 1.53 | 1.11 | 0.06 | 0.002 |
| Treatment 1 | ||
|---|---|---|
| Item | CON | MT |
| Ingredient, % | ||
| Ground yellow corn | 74.25 | 59.25 |
| Ground yellow corn with deoxynivalenol 2 | 0.00 | 15.00 |
| Soybean meal, dehulled | 22.00 | 22.00 |
| L-Lys HCl | 0.10 | 0.10 |
| L-Trp | 0.40 | 0.40 |
| Salt | 0.30 | 0.30 |
| Vitamin premix 3 | 0.03 | 0.03 |
| Trace mineral premix 4 | 0.15 | 0.15 |
| Dicalcium phosphate | 0.90 | 0.90 |
| Ground limestone | 0.70 | 0.70 |
| Poultry fat | 1.00 | 1.00 |
| Calculated composition | ||
| DM, % | 89.6 | 89.6 |
| ME, Mcal/kg | 3.39 | 3.39 |
| CP, % | 17.06 | 17.06 |
| SID 5 Lys, % | 0.83 | 0.83 |
| SID Cys + Met, % | 0.51 | 0.51 |
| SID Trp, % | 0.56 | 0.56 |
| SID Thr, % | 0.53 | 0.53 |
| Ca, % | 0.55 | 0.55 |
| Available P, % | 0.23 | 0.23 |
| Total P, % | 0.53 | 0.53 |
| Deoxynivalenol, mg/kg | 0.00 | 3.00 |
| earalenone, mg/kg | 0.00 | 0.13 |
| Analyzed composition | ||
| DM, % | 88.7 | 89.1 |
| CP, % | 16.3 | 16.4 |
| Deoxynivalenol, mg/kg | 0.00 | 3.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.-B.; Weaver, A.C.; Kim, S.W. Physiological Effects of Deoxynivalenol from Naturally Contaminated Corn on Cerebral Tryptophan Metabolism, Behavioral Response, Gastrointestinal Immune Status and Health in Pigs Following a Pair-Feeding Model. Toxins 2021, 13, 393. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060393
Shen Y-B, Weaver AC, Kim SW. Physiological Effects of Deoxynivalenol from Naturally Contaminated Corn on Cerebral Tryptophan Metabolism, Behavioral Response, Gastrointestinal Immune Status and Health in Pigs Following a Pair-Feeding Model. Toxins. 2021; 13(6):393. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060393
Chicago/Turabian StyleShen, Yan-Bin, Alexandra C. Weaver, and Sung Woo Kim. 2021. "Physiological Effects of Deoxynivalenol from Naturally Contaminated Corn on Cerebral Tryptophan Metabolism, Behavioral Response, Gastrointestinal Immune Status and Health in Pigs Following a Pair-Feeding Model" Toxins 13, no. 6: 393. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060393