Heat-Labile Toxin from Enterotoxigenic Escherichia coli Causes Systemic Impairment in Zebrafish Model

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
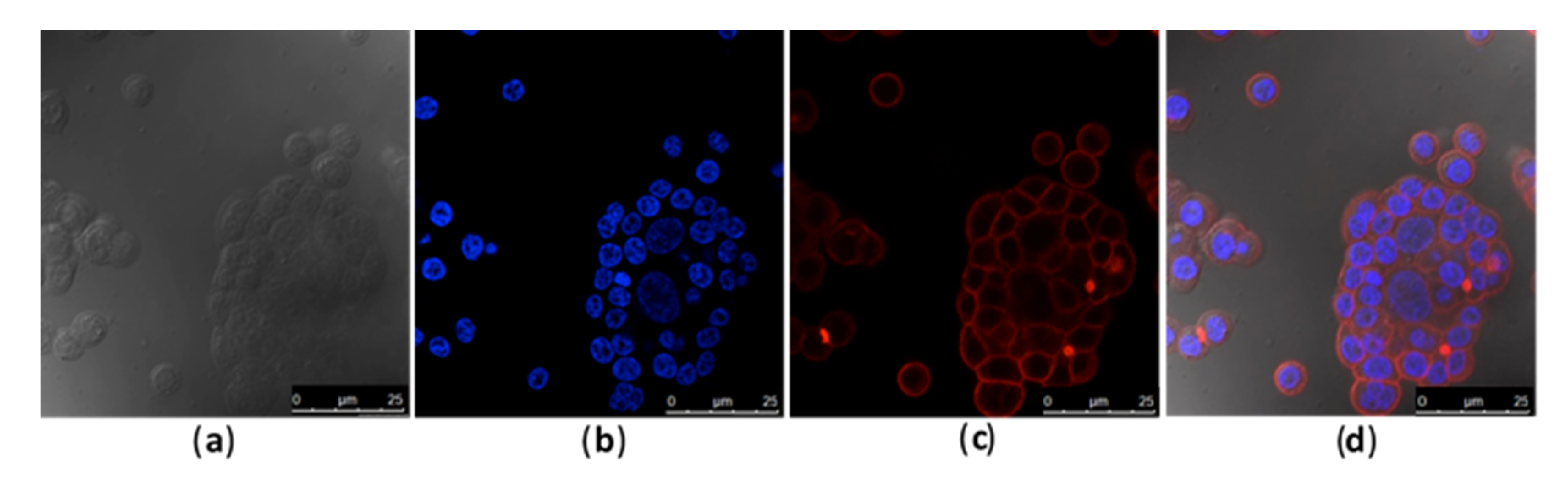
2.1. Toxin and Caco-2 Cell Interaction
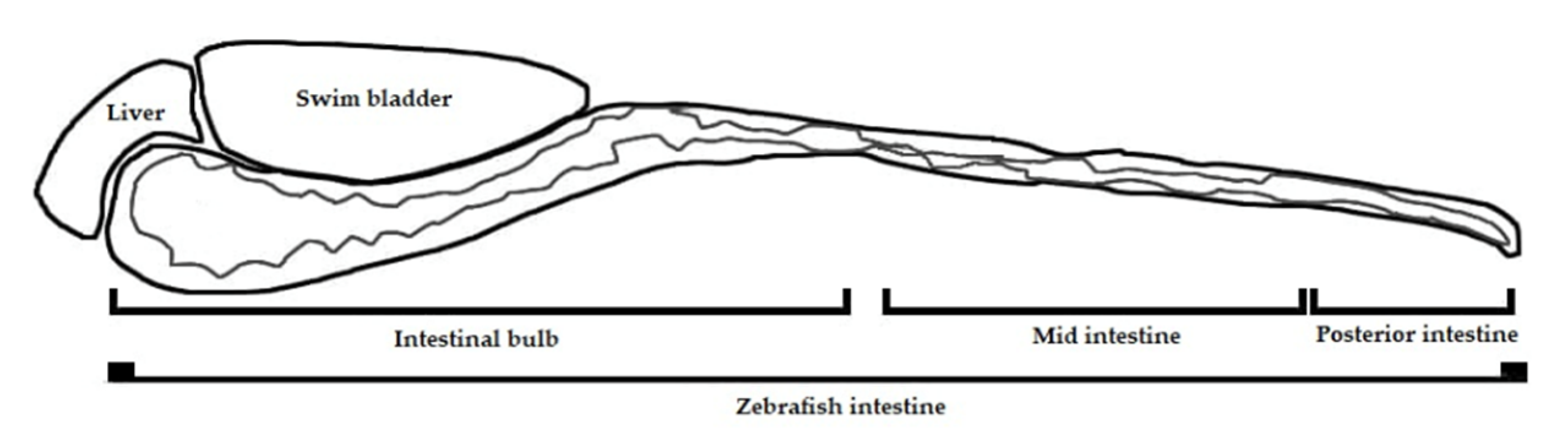
2.2. Systemic Distribution of LT-I Toxin in Zebrafish
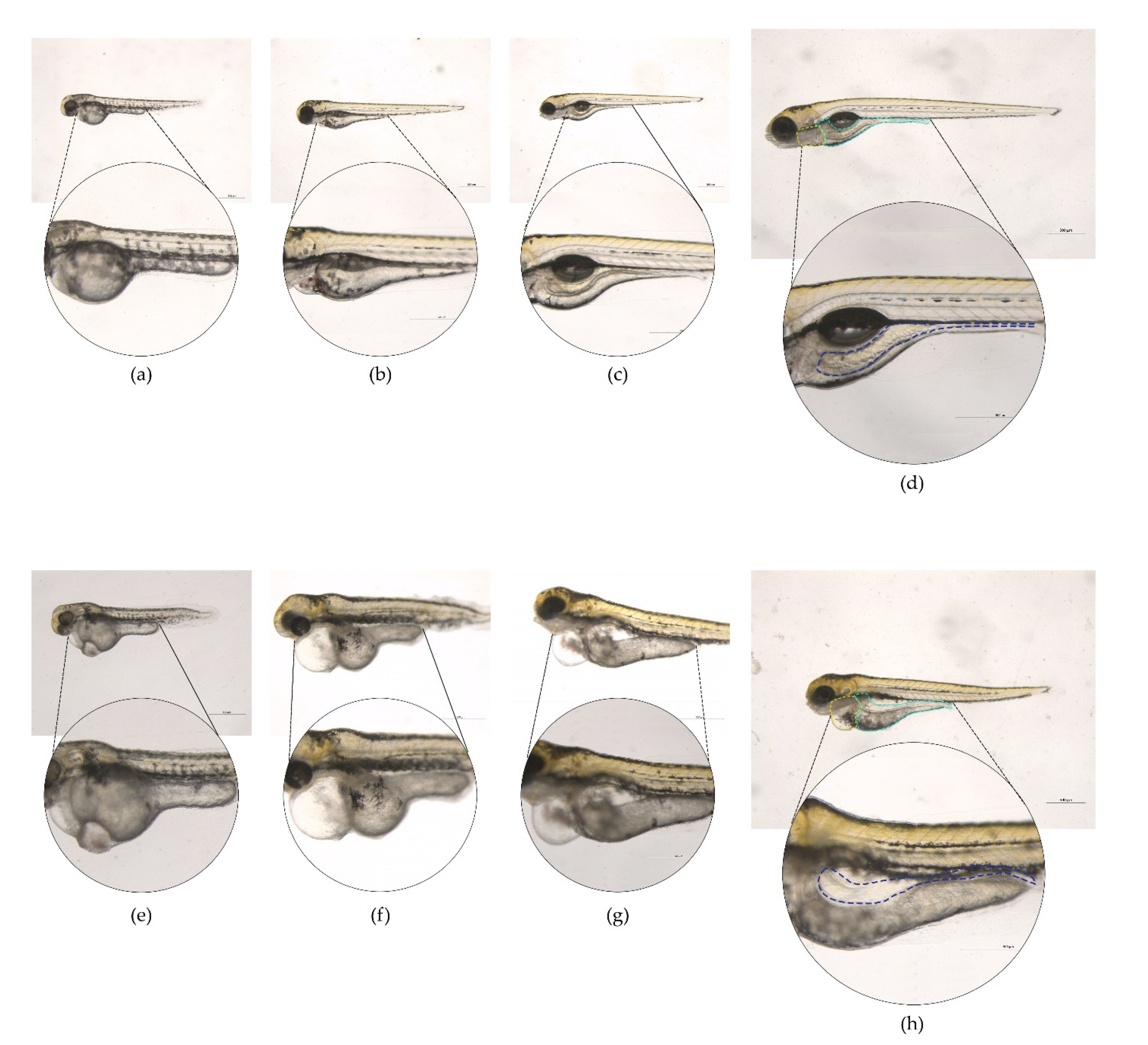
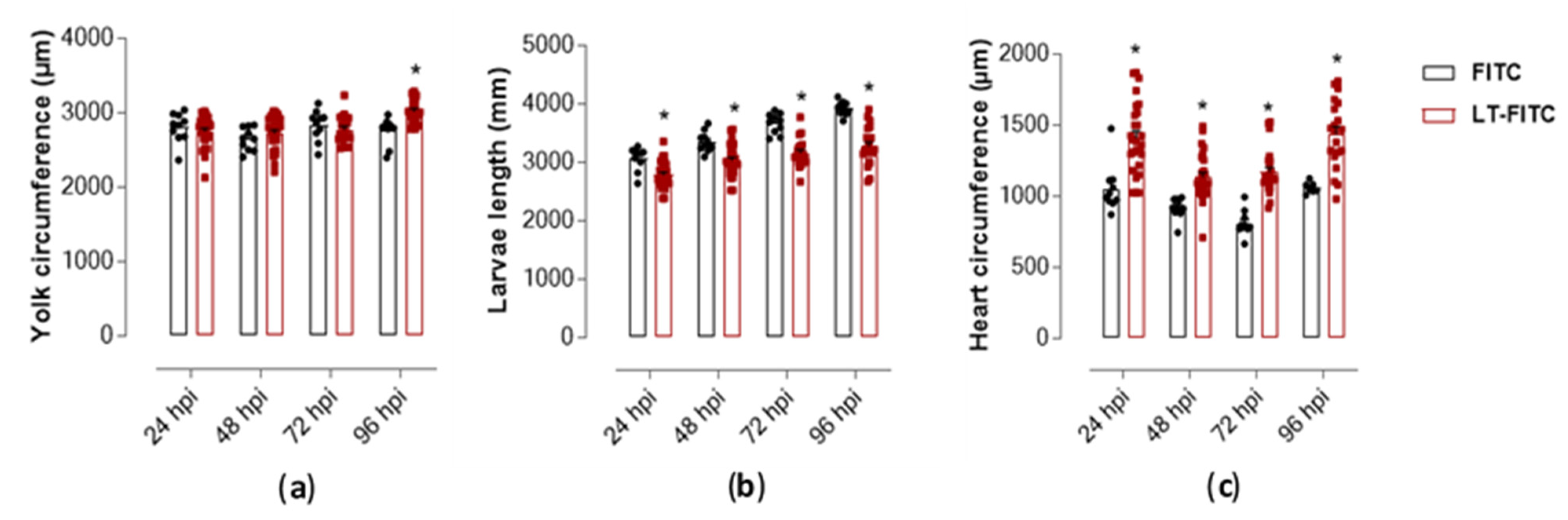
2.3. Phenotypic Assessment in Zebrafish Larvae Exposed to LT-I Toxin
3. Discussion
4. Materials and Methods
4.1. LT Toxin
4.2. Caco-2 Cell
4.3. Conjugation of LT with FITC Fluorophore
4.4. Fluorescence
4.5. Zebrafish Husbandry
4.6. Zebrafish Embryo Toxicity Assay (Fet Test OECD 236)
4.7. Microinjection into Zebrafish Larvae
4.8. Phenotypic Analysis
4.9. Zebrafish Euthanasia
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schaechter, M.; Medoff, G.; Eisenstein, B.I. Mechanisms of Microbial Disease, 2nd ed.; Williams and Wilkins: Baltimore, MD, USA, 1993; pp. 162–175. [Google Scholar]
- Schmitt, C.K.; Meysick, K.C.; O’Brien, A.D. Bacterial toxins: Friends or foes. Emerg. Infect. Dis. 1999, 5, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Spangler, B.D. Structure and Function of Cholera Toxin and the Related Escherichia coli Heat-Labile Enterotoxin. Microbiol. Rev. 1992, 56, 622–647. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, J.M.; Kuhlmann, M. Enterotoxigenic Escherichia coli Infections. Curr. Infect. Dis. Rep. 2019, 21, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mirhoseini, A.; Amani, J.; Nazarianb, S. Review on pathogenicity mechanism of enterotoxigenic Escherichia coli and vaccines against it. Microb. Pathog. 2018, 117, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Xia, P.; Nandre, R.; Zhang, W.; Zhu, G. Review of Newly Identified Functions Associated with the Heat-Labile Toxin of Enterotoxigenic Escherichia coli. Front. Cell. Infect. Microbiol. 2019, 9, 292. [Google Scholar] [CrossRef] [Green Version]
- Villamil, J.S.; Navarro-Garcia, F. Role of virulence factors on host inflammatory response induced by Diarrheagenic Escherichia coli pathotypes. Future Microbiol. 2015, 10, 1009–1033. [Google Scholar] [CrossRef]
- Lanata, C.F.; Fischer-Walker, C.L.; Olascoaga, A.C.; Torres, C.X.; Aryee, M.J.; Black, R.E. Global Causes of Diarrheal Disease Mortality in Children, 5 Years of Age: A Systematic Review. PLoS ONE 2013, 8, e72788. [Google Scholar] [CrossRef] [Green Version]
- Horstman, A.L.; Kuehn, M.J. Enterotoxigenic Escherichia coli Secretes Active Heat-labile Enterotoxin via Outer Membrane Vesicles. J. Biol. Chem. 2000, 275, 12489–12496. [Google Scholar] [CrossRef] [Green Version]
- Horstman, A.L.; Kuehn, M.J. Bacterial Surface Association of Heat-labile Enterotoxin through Lipopolysaccharide after Secretion via the General Secretory Pathway. J. Biol. Chem. 2002, 277, 32538–32545. [Google Scholar] [CrossRef] [Green Version]
- Moss, J.; Richardson, S.H. Activation of adenylate cyclase by heat-labile Escherichia coli enterotoxin. Evidence for ADP-ribosyltransferase activity similar to that of choleragen. J. Clin. Investig. 1978, 62, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.H.; Rich, D.P.; Marshall, J.; Gregory, R.J.; Welsh, M.J.; Smith, A.E. Phosphorylation of the R domain by cAMP-dependent protein kinase regulates the CFTR chloride channel. Cell 1991, 66, 1027–1036. [Google Scholar] [CrossRef]
- Sheppard, D.N.; Welsh, M.J. Structure and Function of the CFTR Chloride Channel. Physiol. Rev. 1999, 79, S23–S45. [Google Scholar] [CrossRef]
- Hu, J.C.; Mathias-Santos, C.; Greene, C.J.; King-Lyons, N.D.; Rodrigues, J.F.; Hajishengallis, G.; Ferreira, L.C.S.; Connell, T.D. Intradermal Administration of the Type II Heat-Labile Enterotoxins LT-IIb and LT-IIc of Enterotoxigenic Escherichia coli Enhances Humoral and CD8+ T Cell Immunity to a Co-Administered Antigen. PLoS ONE 2014, 9, e113978. [Google Scholar] [CrossRef]
- Connell, T.D. Cholera toxin, LT-I, LT-IIa and LT-IIb: The critical role of ganglioside binding in immunomodulation by Type I and Type II heat-labile enterotoxins. Expert Rev. Vaccines 2007, 6, 821–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajishengallis, G.; Arce, S.; Gockel, C.; Connell, T.; Russell, M. Immunomodulation with Enterotoxins for the Generation of Secretory Immunity or Tolerance: Applications for Oral Infections. J. Dent. Res. 2005, 84, 1104–1116. [Google Scholar] [CrossRef] [PubMed]
- Arce, S.; Nawar, H.F.; Muehlinghaus, G.; Russell, M.W.; Connell, T.D. In Vitro Induction of Immunoglobulin A (IgA)- and IgM-Secreting Plasma Blasts by Cholera Toxin Depends on T-Cell Help and Is Mediated by CD154 Up-Regulation and Inhibition of Gamma Interferon Synthesis. Infect. Immun. 2007, 75, 1413–1423. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.G.; Silver, R.P.; Chase, D.G.; Gorbach, S.L. Plasmid-controlled colonization factor associated with virulence in Escherichia coli enterotoxigenic for humans. Infect. Immun. 1975, 12, 656–667. [Google Scholar] [CrossRef] [Green Version]
- Dean, A.G.; Ching, Y.-C.; Williams, R.G.; Harden, L.B. Test for Escherichia coli Enterotoxin Using Infant Mice: Application in a Study of Diarrhea in Children in Honolulu. J. Infect. Dis. 1972, 125, 407–411. [Google Scholar] [CrossRef]
- Smith, H.; Hall, W.S. Observations by the ligated intestinal segment and oral inoculation methods on Escherichia coli infections in pigs, calves, lambs and rabbits. J. Pathol. Bacteriol. 1963, 93, 449–529. [Google Scholar] [CrossRef]
- De, S.N.; Bhattacharya, K.; Sarkar, J.K. A study of the pathogenicity of strains of bacterium coli from acute and chronic enteritis. J. Pathol. Bacteriol. 1956, 71, 201–209. [Google Scholar] [CrossRef]
- Guidry, J.J.; Cárdenas, L.; Cheng, E.; Clements, J.D. Role of receptor binding in toxicity, immunogenicity, and adjuvanticity of Escherichia coli heat-labile enterotoxin. Infect. Immun. 1997, 65, 4943–4950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varas, M.; Ortíz-Severín, J.; Marcoleta, A.E.; Díaz-Pascual, F.; Allende, M.L.; Santiviago, C.A.; Chávez, F.P. Salmonella Typhimurium induces cloacitis-like symptomsin zebrafish larvae. Microb. Pathog. 2017, 107, 317–320. [Google Scholar] [CrossRef]
- Jim, K.K.; Engelen-Lee, J.; van der Sar, A.M.; Bitter, W.; Brouwer, M.C.; van der Ende, A.; Veening, J.-W.; van de Beek, D.; Vandenbroucke-Grauls, C.M.J.E. Infection of zebrafish embryos with live fluorescent Streptococcus pneumoniae as a real-time pneumococcal meningitis model. J. Neuroinflamm. 2016, 13, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saralahti, A.; Rämet, M. Zebrafish and Streptococcal Infections. Scand. J. Immunol. 2015, 82, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Sachidanandan, C. Zebrafish: A Multifaceted Tool for Chemical Biologists. Chem. Rev. 2013, 113, 7952–7980. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Kari, G.; Rodeck, U.; Dicker, A.P. Zebrafish: An Emerging Model System for Human Disease and Drug Discovery. Clin. Pharmacol. Ther. 2007, 82, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Langheinrich, U. Zebrafish: A new model on the pharmaceutical catwalk. BioEssays 2003, 25, 904–912. [Google Scholar] [CrossRef]
- Melo, B.D.S.; Fernandes, B.H.V.; Lopes-Ferreira, M.V.A.; Henrique, C.; Piazza, R.M.F.; Luz, D. Zebrafish embryo sensitivity test as in vivo platform to anti-Shiga toxin compound screening. Braz. J. Microbiol. 2020, 51, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Kavaliauskis, A.; Arnemo, M.; Speth, M.; Lagos, L.; Rishovd, A.-L.; Estepa, A.; Griffiths, G.; Gjøen, T. Protective effect of a recombinant VHSV-G vaccine using poly(I:C) loaded nanoparticles as an adjuvant in zebrafish (Danio rerio) infection model. Dev. Comp. Immunol. 2016, 61, 248–257. [Google Scholar] [CrossRef]
- Kavaliauskis, A.; Arnemo, M.; Kim, S.-H.; Ulanova, L.; Speth, M.; Novoa, B.; Dios, S.; Evensen, Ø.; Griffiths, G.W.; Gjøen, T. Use of Poly(I:C) Stabilized with Chitosan As a Vaccine-Adjuvant Against Viral Hemorrhagic Septicemia Virus Infection in Zebrafish. Zebrafish 2015, 12, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, H.; Gao, Y.; Zhang, Y.; Wu, H.; Zhang, Y. Efficacy of chitosan oligosaccharide as aquatic adjuvant administrated with a formalin-inactivated Vibrio anguillarum vaccine. Fish Shellfish Immunol. 2015, 47, 855–860. [Google Scholar] [CrossRef]
- Bailone, R.L.; Fukushima, H.C.S.; Fernandes, B.H.V.; De Aguiar, L.K.; Corrêa, T.; Janke, H.; Setti, P.G.; Roça, R.D.O.; Borra, R.C. Zebrafish as an alternative animal model in human and animal vaccination research. Lab. Anim. Res. 2020, 36, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Pack, M.T. Modeling intestinal disorders using zebrafish. In The Zebrafish Disease Models and Chemical Screens, 4th ed.; Zon, L.I., Westerfield, H.M., Detrich, W., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 148, pp. 241–270. [Google Scholar] [CrossRef]
- Flores, E.M.; Nguyen, A.T.; Odem, M.A.; Eisenhoffer, G.T.; Krachler, A.M. The zebrafish as a model for gastrointestinal tract–microbe interactions. Cell. Microbiol. 2020, 22, e13152. [Google Scholar] [CrossRef] [Green Version]
- Saslowsky, D.E.; Cho, J.A.; Chinnapen, H.; Massol, R.H.; Chinnapen, D.J.-F.; Wagner, J.S.; De Luca, H.E.; Kam, W.; Paw, B.H.; Lencer, W.I. Intoxication of zebrafish and mammalian cells by cholera toxin depends on the flotillin/reggie proteins but not Derlin-1 or -2. J. Clin. Investig. 2010, 120, 4399–4409. [Google Scholar] [CrossRef] [Green Version]
- Donta, S.T.; Moon, H.W.; Whipp, S.C. Detection of Heat-Labile Escherichia coli Enterotoxin with the Use of Adrenal Cells in Tissue Culture. Science 1974, 183, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Guerrant, R.L.; Brunton, L.L.; Schnaitman, T.C.; Rebhun, L.I.; Gilman, A.G. Cyclic Adenosine Monophosphate and Alteration of Chinese Hamster Ovary Cell Morphology: A Rapid, Sensitive In Vitro Assay for the Enterotoxins of Vibrio cholerae and Escherichia coli. Infect. Immun. 1974, 10, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, C.Y.; Silveira, C.R.F.; Andrade, F.B.; Nepomuceno, R.; Silva, A.; Munhoz, D.D.; Yamamoto, B.B.; Luz, D.; Abreu, P.A.E.; Horton, D.S.P.Q.; et al. Single Chain Variable Fragments Produced in Escherichia coli against Heat-Labile and Heat-Stable Toxins from Enterotoxigenic E. coli. PLoS ONE 2015, 10, e0131484. [Google Scholar] [CrossRef] [Green Version]
- Dorsey, F.C.; Fischer, J.F.; Fleckenstein, J.M. Directed delivery of heat-labile enterotoxin by enterotoxigenic Escherichia coli. Cell. Microbiol. 2006, 8, 1516–1527. [Google Scholar] [CrossRef]
- Bowman, C.C.; Clements, J.D. Differential Biological and Adjuvant Activities of Cholera Toxin and Escherichia coli Heat-Labile Enterotoxin Hybrids. Infect. Immun. 2001, 69, 1528–1535. [Google Scholar] [CrossRef] [Green Version]
- Lopes, L.M.; Maroof, A.; Dougan, G.; Chain, B.M. Inhibition of T-cell Response by Escherichia coli Heat-Labile Enterotoxin-Treated Epithelial Cells. Infect. Immun. 2000, 68, 6891–6895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lea, T. Caco-2 Cell Line. In The Impact of Food Bioactives on Heath: In Vivo and Ex Vivo Models, 1st ed.; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer Open: New York, NY, USA, 2015; pp. 103–111. [Google Scholar] [CrossRef] [Green Version]
- Jahn, K.A.; Biazik, J.M.; Braet, F. GM1 Expression in Caco-2 Cells: Characterisation of a Fundamental Passage-dependent Transformation of a Cell Line. J. Pharm. Sci. 2011, 100, 3751–3762. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Cieplak, W.; Messer, R.J.; Konkel, M.E.; Grant, C.C.R. Role of a potential endoplasmic reticulum retention sequence (RDEL) and the Golgi complex in the cytotonic activity of Escherichia coli heat-labile enterotoxin. Mol. Microbiol. 1995, 16, 789–800. [Google Scholar] [CrossRef]
- Lencer, W.I.; Constable, C.; Moe, S.; Jobling, M.G.; Webb, H.M.; Ruston, S.; Madara, J.L.; Hirst, T.R.; Holmesll, R.K. Targeting of cholera toxin and Escherichia coli heat labile toxin in polarized epithelia: Role of COOH-terminal KDEL. J. Cell Biol. 1995, 131, 951–962. [Google Scholar] [CrossRef]
- Lencer, W.I.; Moe, S.; Rufo, P.A.; Madara, J.L. Transcytosis of cholera toxin subunits across model human intestinal epithelia. Proc. Natl. Acad. Sci. USA 1995, 92, 10094–10098. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.G.; Evans, D.J.; Pierce, N.F. Differences in the Response of Rabbit Small Intestine to Heat-Labile and Heat-Stable Enterotoxins of Escherichia coli. Infect. Immun. 1973, 7, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.P.; Randolph, M.M.; Fleckenstein, J.M. Importance of Heat-Labile Enterotoxin in Colonization of the Adult Mouse Small Intestine by Human Enterotoxigenic Escherichia coli Strains. Infect. Immun. 2006, 74, 869–875. [Google Scholar] [CrossRef] [Green Version]
- Byrd, W.; Cassels, F.J. Mucosal immunization of BALB/c mice using enterotoxigenic Escherichia coli colonization factors CFA/I and CS6 administered with and without a mutant heat-labile enterotoxin. Vaccine 2003, 21, 1884–1893. [Google Scholar] [CrossRef]
- Byrd, W.; Mog, S.R.; Cassels, F.J. Pathogenicity and Immune Response Measured in Mice following Intranasal Challenge with Enterotoxigenic Escherichia coli Strains H10407 and B7A. Infect. Immun. 2003, 71, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Bolick, D.T.; Medeiros, P.H.Q.S.; Ledwaba, S.E.; Lima, A.A.M.; Nataro, J.P.; Barry, E.M.; Guerrant, R.L. Critical Role of Zinc in a New Murine Model of Enterotoxigenic Escherichia coli Diarrhea. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Luiz, W.B.; Rodrigues, J.F.; Crabb, J.H.; Savarino, S.J.; Ferreira, L.C.S. Maternal Vaccination with a Fimbrial Tip Adhesin and Passive Protection of Neonatal Mice against Lethal Human Enterotoxigenic Escherichia coli Challenge. Infect. Immun. 2015, 83, 4555–4564. [Google Scholar] [CrossRef] [Green Version]
- Klipstein, F.A.; Engert, R.F.; Clements, J.D. Protection in rats immunized with Escherichia coli heat-stable enterotoxin. Infect. Immun. 1981, 34, 637–639. [Google Scholar] [CrossRef] [Green Version]
- Spira, W.M.; Sack, R.B.; Froehlich, J.L. Simple adult rabbit model for Vibrio cholerae and enterotoxigenic Escherichia coli diarrhea. Infect. Immun. 1981, 32, 739–747. [Google Scholar] [CrossRef] [Green Version]
- Nag, D.; Mitchell, K.; Breen, P.; Withey, J.H. Quantifying Vibrio cholerae Colonization and Diarrhea in the Adult Zebrafish Model. J. Vis. Exp. 2018, 137, e57767. [Google Scholar] [CrossRef]
- Manneh-Roussel, J.; Haycocks, J.R.J.; Magán, A.; Perez-Soto, N.; Voelz, K.; Camilli, A.; Krachler, A.-M.; Grainger, D.C. cAMP Receptor Protein Controls Vibrio cholerae Gene Expression in Response to Host Colonization. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Runft, D.L.; Mitchell, K.C.; Abuaita, B.H.; Allen, J.P.; Bajer, S.; Ginsburg, K.; Neely, M.N.; Withey, J.H. Zebrafish as a Natural Host Model for Vibrio cholerae Colonization and Transmission. Appl. Environ. Microbiol. 2013, 80, 1710–1717. [Google Scholar] [CrossRef] [Green Version]
- Dubreuil, J.D. Antibacterial and Antidiarrheal Activities of Plant Products against Enterotoxinogenic Escherichia coli. Toxins 2013, 5, 2009–2041. [Google Scholar] [CrossRef]
- OECD. OECD Guidelines for the Testing of Chemicals, Section 2 Test No. 203: Fish, Acute Toxicity Test; OECD Publishing: Paris, France, 2013. [Google Scholar] [CrossRef]
- Ali, S.; Champagne, D.L.; Spaink, H.P.; Richardson, M.K. Zebrafish embryos and larvae: A new generation of disease models and drug screens. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 115–133. [Google Scholar] [CrossRef]
- Grunwald, D.J.; Eisen, J.S. Headwaters of the zebrafish—Emergence of a new model vertebrate. Nat. Rev. Genet. 2002, 3, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R.; Lahav, M.; Guo, S.; Rosenthal, A. Drinks like a fish: Zebra fish (Danio rerio) as a behavior genetic model to study alcohol effects. Pharmacol. Biochem. Behav. 2000, 67, 773–782. [Google Scholar] [CrossRef]
- Tanguay, R.L.; Reimers, M.J. Analysis of Ethanol Developmental Toxicity in Zebrafish. Adv. Struct. Saf. Stud. 2008, 447, 63–74. [Google Scholar] [CrossRef]
- Dahm, R.; Geisler, R. Learning from Small Fry: The Zebrafish as a Genetic Model Organism for Aquaculture Fish Species. Mar. Biotechnol. 2006, 8, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Hajishengallis, G. Heat-Labile Enterotoxins as Adjuvants or Anti-Inflammatory Agents. Immunol. Investig. 2010, 39, 449–467. [Google Scholar] [CrossRef] [Green Version]
- Van Ginkel, F.W.; Jackson, R.J.; Yoshino, N.; Hagiwara, Y.; Metzger, D.J.; Connell, T.D.; Vu, H.L.; Martin, M.; Fujihashi, K.; McGhee, J.R. Enterotoxin-Based Mucosal Adjuvants Alter Antigen Trafficking and Induce Inflammatory Responses in the Nasal Tract. Infect. Immun. 2005, 73, 6892–6902. [Google Scholar] [CrossRef] [Green Version]
- Van Ginkel, F.W.; Jackson, R.J.; Yuki, Y.; McGhee, J.R. Cutting Edge: The Mucosal Adjuvant Cholera Toxin Redirects Vaccine Proteins into Olfactory Tissues. J. Immunol. 2000, 165, 4778–4782. [Google Scholar] [CrossRef] [Green Version]
- Zoeteweij, J.P.; Epperson, D.E.; Porter, J.D.; Zhang, C.X.; Frolova, O.Y.; Constantinides, A.P.; Fuhrmann, S.R.; El-Amine, M.; Tian, J.-H.; Ellingsworth, L.R.; et al. GM1 Binding-Deficient Exotoxin Is a Potent Noninflammatory Broad Spectrum Intradermal Immunoadjuvant. J. Immunol. 2006, 177, 1197–1207. [Google Scholar] [CrossRef] [Green Version]
- Rigoni, M.; Paoli, M.; Milanesi, E.; Caccin, P.; Rasola, A.; Bernardi, P.; Montecucco, C. Snake Phospholipase A2 Neurotoxins Enter Neurons, Bind Specifically to Mitochondria, and Open Their Transition Pores. J. Biol. Chem. 2008, 283, 34013–34020. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henrique, C.; Falcão, M.A.P.; De Araújo Pimenta, L.; Maleski, A.L.A.; Lima, C.; Mitsunari, T.; Sampaio, S.C.; Lopes-Ferreira, M.; Piazza, R.M.F. Heat-Labile Toxin from Enterotoxigenic Escherichia coli Causes Systemic Impairment in Zebrafish Model. Toxins 2021, 13, 419. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060419
Henrique C, Falcão MAP, De Araújo Pimenta L, Maleski ALA, Lima C, Mitsunari T, Sampaio SC, Lopes-Ferreira M, Piazza RMF. Heat-Labile Toxin from Enterotoxigenic Escherichia coli Causes Systemic Impairment in Zebrafish Model. Toxins. 2021; 13(6):419. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060419
Chicago/Turabian StyleHenrique, Camila, Maria Alice Pimentel Falcão, Luciana De Araújo Pimenta, Adolfo Luís Almeida Maleski, Carla Lima, Thais Mitsunari, Sandra Coccuzzo Sampaio, Mônica Lopes-Ferreira, and Roxane Maria Fontes Piazza. 2021. "Heat-Labile Toxin from Enterotoxigenic Escherichia coli Causes Systemic Impairment in Zebrafish Model" Toxins 13, no. 6: 419. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060419