The Regulatory Mechanism of Water Activities on Aflatoxins Biosynthesis and Conidia Development, and Transcription Factor AtfB Is Involved in This Regulation


Abstract
:1. Introduction
2. Results
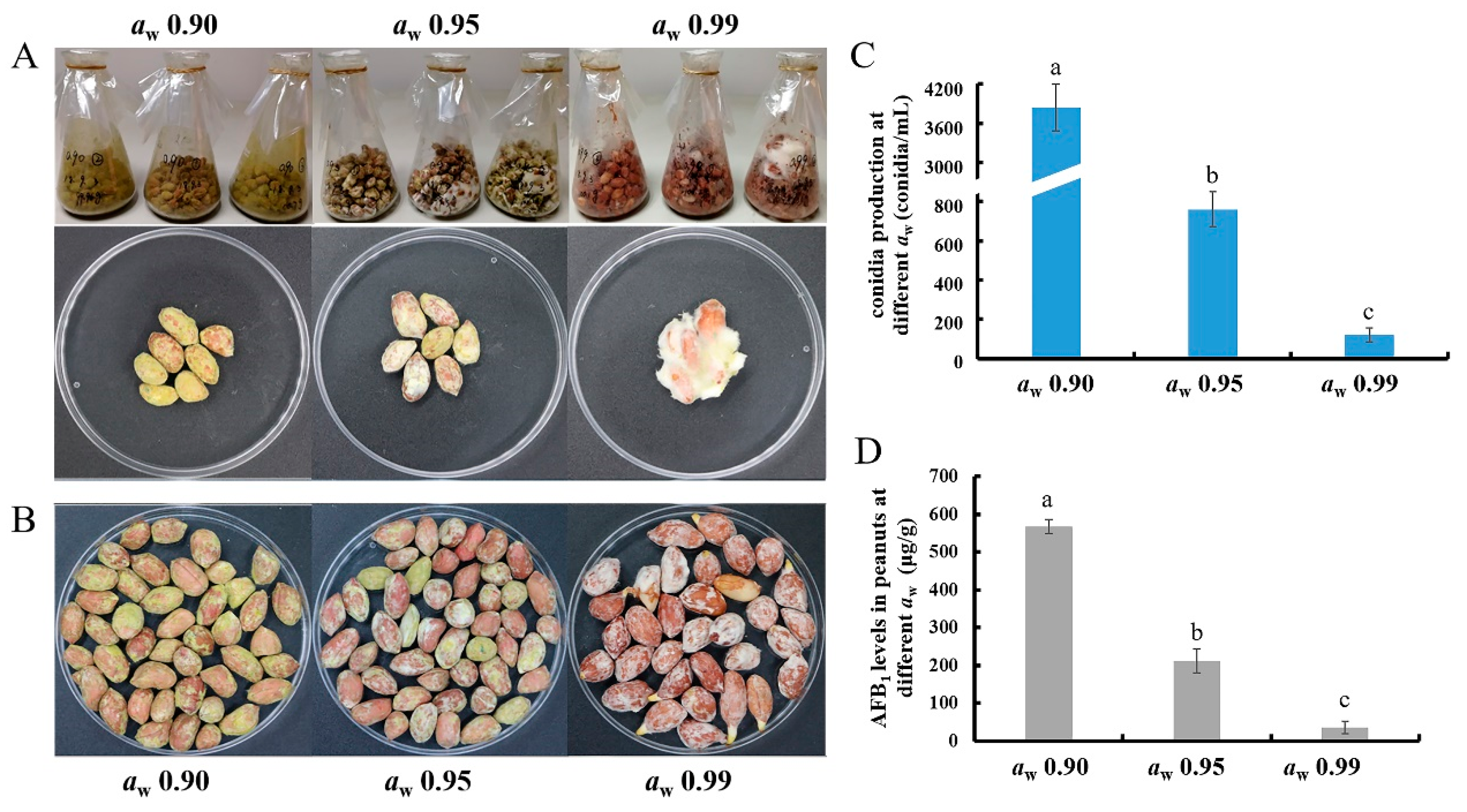
2.1. Water Activity Affects the Conidia Production and the AFB1 Production of A. flavus in Peanuts
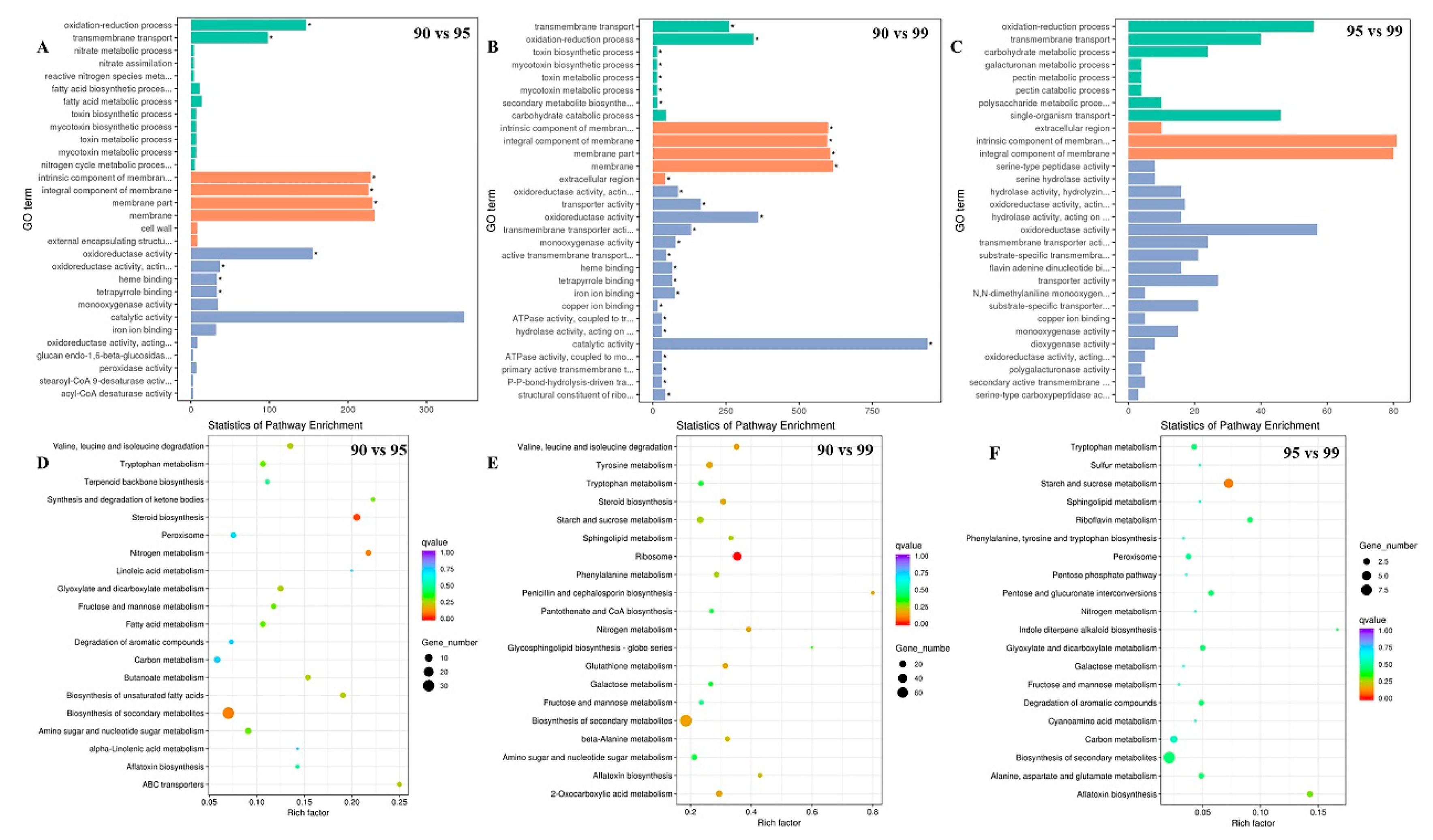
2.2. Transcriptome Analyses of the A. flavus Genes Expressions in Different Water Activity
2.3. Expression Changes of AF Cluster Genes in Different aw Conditions
2.4. Varying Expressions of Diverse Regulator-Associated AF Biosynthesis in Different aw Conditions
2.5. Different Expression of the Genes Controlling Conidia Production in Different Water Activities
2.6. The Effects of Diverse Water Activities on Transcription Factors
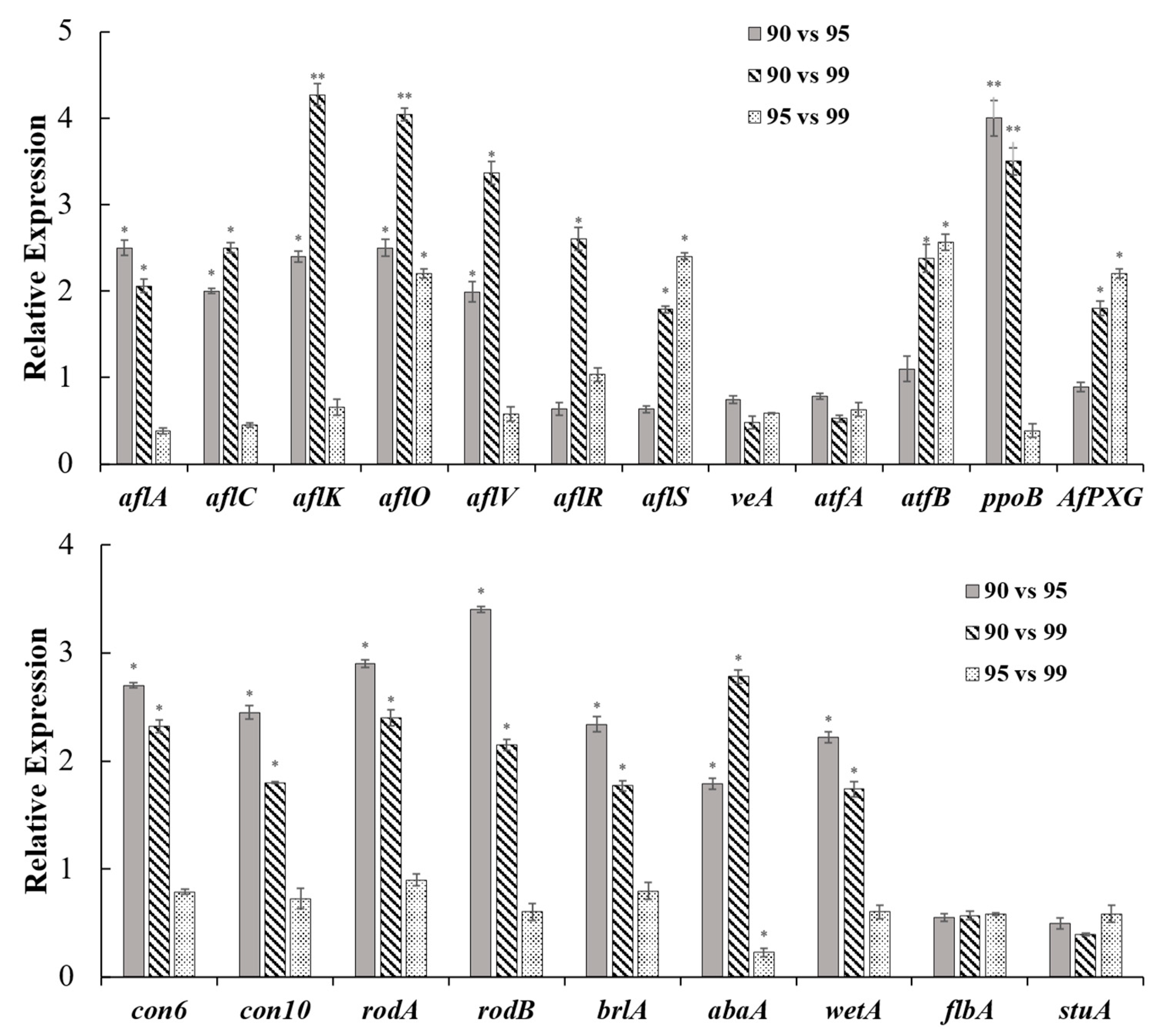
2.7. RT-qRCR Analyses of Genes Expressions Involved in AF Biosynthesis and Conidia Development
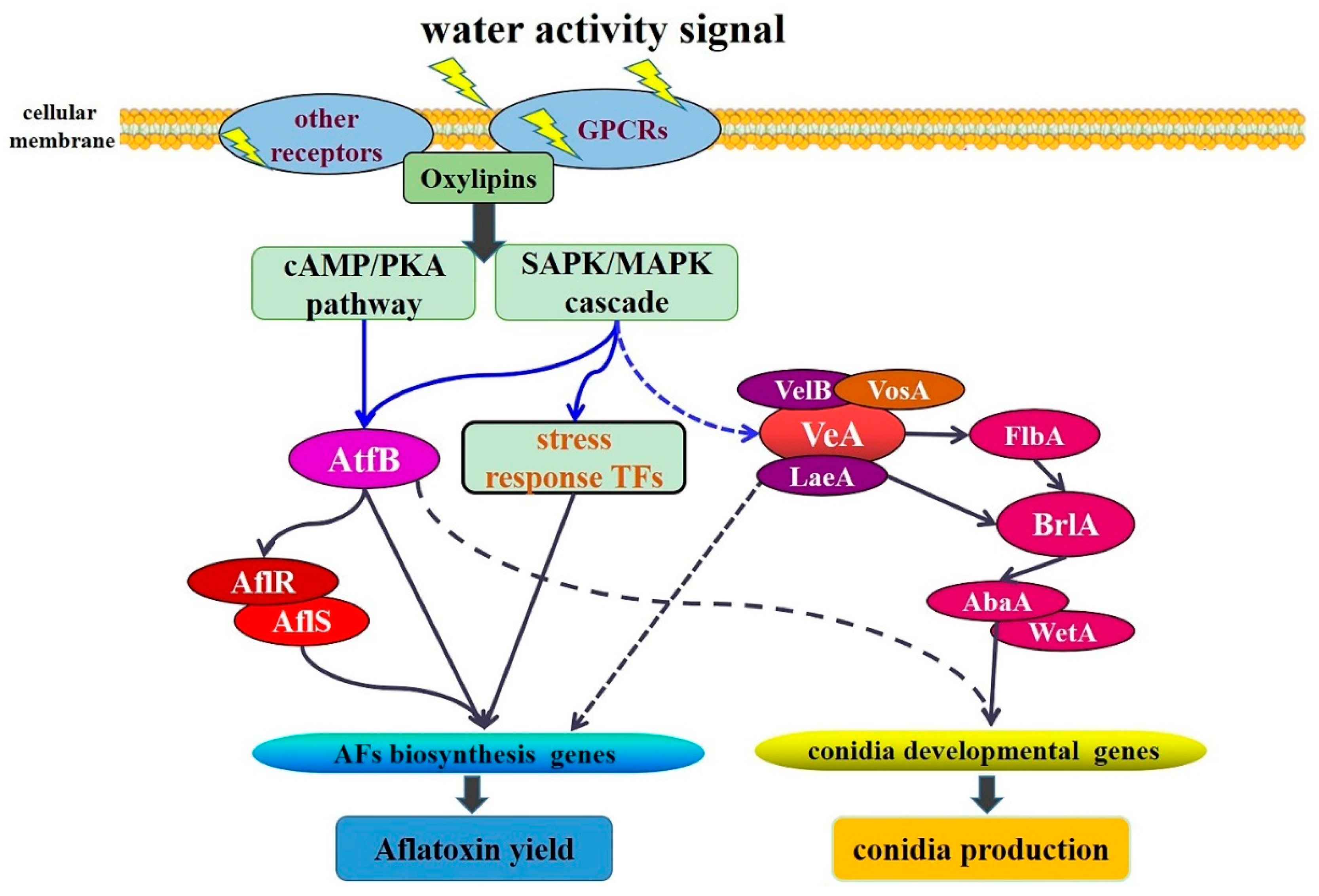
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Strain and Conidia Suspension Preparation
5.2. Adjustment of Peanut Water Activities and Inoculation of A. flavus Conidia Suspension
5.3. Conidia Assessment and AFB1 Detection
5.4. Total RNA Extraction
5.5. RNA Sequencing and Transcriptome Processing
5.6. RT-qPCR Analysis
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, F.; Zhong, H.; Han, X.; Guo, Z.; Yang, W.; Liu, Y.; Yang, K.; Zhuang, Z.; Wang, S. Proteomic profile of Aspergillus flavus in response to water activity. Fungal Biol. 2015, 119, 114–124. [Google Scholar] [CrossRef]
- Ren, Y.; Jin, J.; Zheng, M.; Yang, Q.; Xing, F. Ethanol Inhibits Aflatoxin B(1) Biosynthesis in Aspergillus flavus by Up-Regulating Oxidative Stress-Related Genes. Front. Microbiol. 2019, 10, 2946. [Google Scholar] [CrossRef] [Green Version]
- Wu, F. Perspective: Time to face the fungal threat. Nature 2014, 516, S7. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hadi, A.; Schmidt-Heydt, M.; Parra, R.; Geisen, R.; Magan, N. A systems approach to model the relationship between aflatoxin gene cluster expression, environmental factors, growth and toxin production by Aspergillus flavus. J. R. Soc. Interface 2012, 9, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Tai, B.; Chang, J.; Liu, Y.; Xing, F. Recent progress of the effect of environmental factors on Aspergillus flavus growth and aflatoxins production on foods. Food Qual. Saf. 2020, 4, 21–28. [Google Scholar] [CrossRef]
- Liu, X.; Guan, X.; Xing, F.; Lv, C.; Dai, X.; Liu, Y. Effect of water activity and temperature on the growth of Aspergillus flavus, the expression of aflatoxin biosynthetic genes and aflatoxin production in shelled peanuts. Food Control. 2017, 82, 325–332. [Google Scholar] [CrossRef]
- Passamani, F.R.; Hernandes, T.; Lopes, N.A.; Bastos, S.C.; Santiago, W.D.; Cardoso, M.; Batista, L.R. Effect of temperature, water activity, and pH on growth and production of ochratoxin A by Aspergillus niger and Aspergillus carbonarius from Brazilian grapes. J. Food Prot. 2014, 77, 1947–1952. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Gilbert, M.K.; Mack, B.M.; GR, O.B.; Rodríguez, A.; Bhatnagar, D.; Payne, G.; Magan, N. Interactions between water activity and temperature on the Aspergillus flavus transcriptome and aflatoxin B(1) production. Int. J. Food Microbiol. 2017, 256, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Guo, Z.; Zhong, H.; Wang, S.; Yang, W.; Liu, Y.; Wang, S. RNA-Seq-based transcriptome analysis of aflatoxigenic Aspergillus flavus in response to water activity. Toxins 2014, 6, 3187–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Fedorova, N.D.; Montalbano, B.G.; Bhatnagar, D.; Cleveland, T.E.; Bennett, J.W.; Nierman, W.C. Tight control of mycotoxin biosynthesis gene expression in Aspergillus flavus by temperature as revealed by RNA-Seq. FEMS Microbiol. Lett. 2011, 322, 145–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.Y.; Roze, L.V.; Linz, J.E. Oxidative stress-related transcription factors in the regulation of secondary metabolism. Toxins 2013, 5, 683–702. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hadi, A.; Carter, D.; Magan, N. Temporal monitoring of the nor-1 (aflD) gene of Aspergillus flavus in relation to aflatoxin B₁ production during storage of peanuts under different water activity levels. J. Appl. Microbiol. 2010, 109, 1914–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, C.; Jin, J.; Wang, P.; Dai, X.; Liu, Y.; Zheng, M.; Xing, F. Interaction of water activity and temperature on the growth, gene expression and aflatoxin production by Aspergillus flavus on paddy and polished rice. Food Chem. 2019, 293, 472–478. [Google Scholar] [CrossRef]
- Peromingo, B.; Rodríguez, A.; Bernáldez, V.; Delgado, J.; Rodríguez, M. Effect of temperature and water activity on growth and aflatoxin production by Aspergillus flavus and Aspergillus parasiticus on cured meat model systems. Meat Sci. 2016, 122, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Solfrizzo, M.; Epifani, F.; Panzarini, G.; Perrone, G. Effect of temperature and water activity on gene expression and aflatoxin biosynthesis in Aspergillus flavus on almond medium. Int. J. Food Microbiol. 2016, 217, 162–169. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Abdel-Hadi, A.; Magan, N.; Geisen, R. Complex regulation of the aflatoxin biosynthesis gene cluster of Aspergillus flavus in relation to various combinations of water activity and temperature. Int. J. Food Microbiol. 2009, 135, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, K.C. Predicted roles of the uncharacterized clustered genes in aflatoxin biosynthesis. Toxins 2009, 1, 37–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered pathway genes in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Caceres, I.; El Khoury, R.; Bailly, S.; Oswald, I.P.; Puel, O.; Bailly, J.D. Piperine inhibits aflatoxin B1 production in Aspergillus flavus by modulating fungal oxidative stress response. Fungal Genet Biol. 2017, 107, 77–85. [Google Scholar] [CrossRef]
- Zhuang, Z.; Lohmar, J.M.; Satterlee, T.; Cary, J.W.; Calvo, A.M. The Master Transcription Factor mtfA Governs Aflatoxin Production, Morphological Development and Pathogenicity in the Fungus Aspergillus flavus. Toxins 2016, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.V.; Chanda, A.; Wee, J.; Awad, D.; Linz, J.E. Stress-related transcription factor AtfB integrates secondary metabolism with oxidative stress response in aspergilli. J. Biol. Chem. 2011, 286, 35137–35148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wee, J.; Hong, S.Y.; Roze, L.V.; Day, D.M.; Chanda, A.; Linz, J.E. The Fungal bZIP Transcription Factor AtfB Controls Virulence-Associated Processes in Aspergillus parasiticus. Toxins 2017, 9, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Ren, Y.; Jing, J.; Jiang, Y.; Yang, Q.; Luo, S.; Xing, F. The inhibitory mechanism of methyl jasmonate on Aspergillus flavus growth and aflatoxin biosynthesis and two novel transcription factors are involved in this action. Food Res. Int. 2021, 140, 110051. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.V.; Miller, M.J.; Rarick, M.; Mahanti, N.; Linz, J.E. A novel cAMP-response element, CRE1, modulates expression of nor-1 in Aspergillus parasiticus. J. Biol. Chem. 2004, 279, 27428–27439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, N.; Vasseur, V.; Coroller, L.; Dantigny, P.; Rigalma, K. Temperature, water activity and pH during conidia production affect the physiological state and germination time of Penicillium species. Int. J. Food Microbiol. 2017, 241, 151–160. [Google Scholar] [CrossRef]
- Pardo, E.; Lagunas, U.; Sanchis, V.; Ramos, A.J.; Marín, S. Influence of water activity and temperature on conidial germination and mycelial growth of ochratoxigenic isolates of Aspergillus ochraceus on grape juice synthetic medium. Predictive models. J. Sci. Food Agric. 2005, 85, 1681–1686. [Google Scholar] [CrossRef]
- Olmedo, M.; Ruger-Herreros, C.; Luque, E.M.; Corrochano, L.M. A complex photoreceptor system mediates the regulation by light of the conidiation genes con-10 and con-6 in Neurospora crassa. Fungal Genet Biol. 2010, 47, 352–363. [Google Scholar] [CrossRef]
- Suzuki, S.; Sarikaya Bayram, Ö.; Bayram, Ö.; Braus, G.H. conF and conJ contribute to conidia germination and stress response in the filamentous fungus Aspergillus nidulans. Fungal Genet Biol. 2013, 56, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, M.H.; Borodina, I.; Moresco, J.L.; Svendsen, W.E.; Frisvad, J.C.; Søndergaard, I. High-yield production of hydrophobins RodA and RodB from Aspergillus fumigatus in Pichia pastoris. Appl. Microbiol. Biotechnol. 2011, 90, 1923–1932. [Google Scholar] [CrossRef]
- Twumasi-Boateng, K.; Yu, Y.; Chen, D.; Gravelat, F.N.; Nierman, W.C.; Sheppard, D.C. Transcriptional profiling identifies a role for BrlA in the response to nitrogen depletion and for StuA in the regulation of secondary metabolite clusters in Aspergillus fumigatus. Eukaryot Cell 2009, 8, 104–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewall, T.C.; Mims, C.W.; Timberlake, W.E. abaA controls phialide differentiation in Aspergillus nidulans. Plant Cell 1990, 2, 731–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrianopoulos, A.; Timberlake, W.E. The Aspergillus nidulans abaA gene encodes a transcriptional activator that acts as a genetic switch to control development. Mol. Cell. Biol. 1994, 14, 2503–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, M.A.; Timberlake, W.E. Aspergillus nidulans wet A activates spore-specific gene expression. Mol. Cell. Biol. 1991, 11, 55–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.S.; Nam, T.Y.; Han, K.H.; Kim, S.C.; Yu, J.H. VelC positively controls sexual development in Aspergillus nidulans. PLoS ONE 2014, 9, e89883. [Google Scholar] [CrossRef] [Green Version]
- Park, H.S.; Bayram, O.; Braus, G.H.; Kim, S.C.; Yu, J.H. Characterization of the velvet regulators in Aspergillus fumigatus. Mol. Microbiol. 2012, 86, 937–953. [Google Scholar] [CrossRef]
- Ni, M.; Yu, J.H. A novel regulator couples sporogenesis and trehalose biogenesis in Aspergillus nidulans. PLoS ONE 2007, 2, e970. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, K.; Arima, T.H.; Iwashita, K.; Yamada, O.; Gomi, K.; Akita, O. Aspergillus oryzae atfB encodes a transcription factor required for stress tolerance in conidia. Fungal Genet Biol. 2008, 45, 922–932. [Google Scholar] [CrossRef]
- Vienken, K.; Fischer, R. The Zn(II)2Cys6 putative transcription factor NosA controls fruiting body formation in Aspergillus nidulans. Mol. Microbiol. 2006, 61, 544–554. [Google Scholar] [CrossRef]
- Soukup, A.A.; Farnoodian, M.; Berthier, E.; Keller, N.P. NosA, a transcription factor important in Aspergillus fumigatus stress and developmental response, rescues the germination defect of a laeA deletion. Fungal Genet Biol. 2012, 49, 857–865. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.H.; Hynes, M.J.; Todd, R.B.; Davis, M.A. Transcriptional control of nmrA by the bZIP transcription factor MeaB reveals a new level of nitrogen regulation in Aspergillus nidulans. Mol. Microbiol. 2007, 66, 534–551. [Google Scholar] [CrossRef] [PubMed]
- Downes, D.J.; Davis, M.A.; Kreutzberger, S.D.; Taig, B.L.; Todd, R.B. Regulation of the NADP-glutamate dehydrogenase gene gdhA in Aspergillus nidulans by the Zn(II)2Cys6 transcription factor LeuB. Microbiology 2013, 159, 2467–2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polotnianka, R.; Monahan, B.J.; Hynes, M.J.; Davis, M.A. TamA interacts with LeuB, the homologue of Saccharomyces cerevisiae Leu3p, to regulate gdhA expression in Aspergillus nidulans. Mol. Genet Genom. 2004, 272, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priebe, S.; Linde, J.; Albrecht, D.; Guthke, R.; Brakhage, A.A. FungiFun: A web-based application for functional categorization of fungal genes and proteins. Fungal Genet Biol. 2011, 48, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene_ID (AFLA_) | Gene | Gene Function | Log2 (90/95) | Log2 (90/99) | Log2 (95/99) |
|---|---|---|---|---|---|
| 139100 | aflYe | Ser-Thr protein phosphatase family protein | −0.45 | −1.21 | −0.78 |
| 139110 | aflYd | sugar regulator | −0.86 | −0.33 | 0.50 |
| 139120 | aflYc | glucosidase | −0.42 | −0.59 | −0.19 |
| 139130 | aflYb | putative hexose transporter | −0.13 | −0.59 | −0.48 |
| 139140 | aflYa | NADH oxidase | 3.94 * | 4.05 * | 0.09 |
| 139150 | aflY | hypothetical protein | 4.96 * | 5.20 * | 0.24 |
| 139160 | aflX | monooxygenase | 4.58 * | 5.92 * | 1.33 |
| 139170 | aflW | monooxygenase | 4.42 * | 6.23 * | 1.81 * |
| 139180 | aflV | cytochrome P450 monooxygenase | 5.33 * | 12.53 * | 7.18 * |
| 139190 | aflK | VERB synthase | 4.79 * | 11.23 * | 6.43 * |
| 139200 | aflQ | cytochrome P450 monooxigenase | 5.14 * | 11.79 * | 6.65 * |
| 139210 | aflP | O-methyltransferase A | 5.05 * | 11.05 * | 5.99 * |
| 139220 | aflO | O-methyltransferase B | 5.03 * | 12.05 * | 10.83 * |
| 139230 | aflI | cytochrome P450 monooxigenase | 6.21 * | 13.05 * | 6.95 * |
| 139240 | aflLa | hypothetical protein | 5.40 * | 14.05 * | 8.11 * |
| 139250 | aflL | P450 monooxygenase | 4.73 * | 13.77 * | 9.03 * |
| 139260 | aflG | cytochrome P450 monooxygenase | 4.22 * | 6.17 * | 1.94 * |
| 139270 | aflNa | hypothetical protein | 0.83 | 1.32 | 0.48 |
| 139280 | aflN | monooxygenase | 4.05* | 7.46 * | 3.39 * |
| 139290 | aflMa | hypothetical protein | 4.30 * | 9.85 * | 5.53 * |
| 139300 | aflM | ketoreductase | 4.53 * | 12.29 * | 7.74 * |
| 139310 | aflE | NOR reductase | 4.34 * | 7.97 * | 3.63 * |
| 139320 | aflJ | esterase | 4.06 * | 6.95 * | 2.89 * |
| 139330 | aflH | short chain alcohol dehydrogenase | 3.64 * | 5.06 * | 1.41 |
| 139340 | aflS | pathway regulator | 0.54 | 3.51 * | 0.96 |
| 139360 | aflR | transcription activator | 0.43 | 1.82 * | 1.37 |
| 139370 | aflB | fatty acid synthase beta subunit | 1.22 | 2.59 * | 1.36 |
| 139380 | aflA | fatty acid synthase alpha subunit | 1.73 | 2.06 * | 0.31 |
| 139390 | aflD | reductase | 3.35 * | 3.73 * | 0.37 |
| 139400 | aflCa | hypothetical protein | 4.19 * | 4.46 * | 0.26 |
| 139410 | aflC | polyketide synthase | 2.85 * | 2.73 * | −0.14 |
| 139420 | aflT | transmembrane protein | −0.10 | 0.22 | 0.31 |
| 139430 | aflU | P450 monooxygenase | −0.83 | 0.15 | 0.96 |
| 139440 | aflF | dehydrogenase | −0.61 | −0.16 | 0.44 |
| Gene_ID (AFLA_) | Gene Annotation | Log2 (90/95) | Log2 (90/99) | Log2 (95/99) |
|---|---|---|---|---|
| 044790 | conidiation-specific family protein | 0.42 | 3.54 * | 3.11 * |
| 044800 | conidiation protein Con6, putative | 3.18 * | 8.32 * | 5.13 * |
| 083110 | conidiation-specific protein (Con10), putative | 2.78 * | 6.32 * | 3.54 * |
| 098380 | conidial hydrophobin RodA/RolA | 6.49 * | 8.68 * | 2.18 * |
| 014260 | conidial hydrophobin RodB/HypB | 3.21 * | 3.10 * | −0.13 |
| 006180 | conidial pigment biosynthesis oxidase Arb2/brown2 | 5.76 * | 6.39 * | 0.61 |
| 016140 | conidial pigment biosynthesis scytalone dehydratase Arp1 | −1.57 | −1.47 | −0.08 |
| 079710 | conidiophore development protein HymA | −0.01 | 0.88 | 0.87 |
| 082850 | C2H2 type conidiation transcription factor BrlA | 3.62 * | 5.90 * | 2.27 * |
| 029620 | transcription factor AbaA | 4.19 * | 2.52 * | −1.69 |
| 134030 | developmental regulator FlbA | −0.11 | −1.10 | −1.01 |
| 137320 | C2H2 conidiation transcription factor FlbC | −1.10 | 1.12 | 1.21 |
| 080170 | MYB family conidiophore development protein FlbD | −0.60 | −0.87 | 0.28 |
| 026900 | developmental regulator VosA | 2.45 * | 1.42 * | −1.05 |
| 046990 | APSES transcription factor StuA | 0.24 | 1.07 | 0.81 |
| 052030 | developmental regulatory protein WetA | 2.10 * | 2.60 * | 0.48 |
| 101920 | extracellular developmental signal biosynthesis protein FluG | 0.06 | 0.40 | 0.32 |
| Gene ID (AFLA_) | Gene Description | log2 (90/95) | log2 (90/99) | log2 (95/99) |
|---|---|---|---|---|
| 013240 | C6 transcription factor, putative | −2.41 | −2.10 * | 0.30 |
| 015790 | C6 transcription factor (Leu3), putative | 0.19 | 1.96 * | 1.74 |
| 021930 | C6 transcription factor RosA | 0.53 | 1.74 * | 1.19 |
| 023040 | C6 transcription factor, putative | −3.02 | −4.27 * | −1.25 |
| 025720 | C6 transcription factor NosA | 2.46 | 2.46 * | 2.21 * |
| 029620 | transcription factor AbaA | 4.19 * | 2.52 * | −1.69 |
| 030580 | C2H2 transcription factor PacC, putative | −0.50 | −2.02 * | −1.53 |
| 031790 | bZIP transcription factor (MeaB), putative | −0.56 | −1.80 * | −1.26 |
| 033480 | C6 transcription factor, putative | 1.02 | 1.85 * | 0.81 |
| 035590 | C6 transcription factor, putative | −0.16 | 2.75 * | 2.25 |
| 040300 | C6 transcription factor, putative | 2.36 * | 2.75 | 0.37 |
| 051900 | zinc knuckle transcription factor (CnjB), putative | 0.48 | 2.73 * | 2.23 |
| 056780 | C6 transcription factor, putative | −0.84 | −2.27 * | −1.44 |
| 059510 | fungal specific transcription factor, putative | −0.95 | −1.76 * | −0.84 |
| 070970 | C6 transcription factor, putative | 0.60 | 1.61 * | 1.00 |
| 074200 | C6 transcription factor, putative | −0.76 | −1.90 * | −1.16 |
| 076320 | C6 transcription factor, putative | 1.24 | 2.61 * | 1.35 |
| 078500 | bZIP transcription factor, putative | 0.92 | 2.65 * | 1.72 |
| 082850 | C2H2 type conidiation transcription factor BrlA | 3.62 * | 5.90 * | 2.27 * |
| 083460 | C6 transcription factor RosA-like, putative | −1.64 | −1.91 * | −0.28 |
| 083560 | C6 transcription factor, putative | 0.72 | 2.01 * | 1.28 |
| 084720 | C6 transcription factor, putative | 0.68 | 2.56 * | 1.87 |
| 085880 | BTB domain transcription factor, putative | 1.14 | 1.42 * | 0.27 |
| 087810 | bZIP transcription factor, putative | 0.51 | 2.69 * | 2.17 |
| 094010 | bZIP transcription factor (Atf21), putative | 1.06 | 3.69 * | 2.60 * |
| 095090 | C6 transcription factor, putative | 1.87 | 5.79 * | 3.90 |
| 109220 | C6 transcription factor, putative | 0.77 | 1.95 * | 1.16 |
| Novel00457 | fungal specific transcription factor [Aspergillus oryzae RIB40] | 1.72 * | 2.25 * | −0.52 |
| Novel00611 | transcription factor [Aspergillus oryzae RIB40] | −1.08 | −3.22 * | −2.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Li, X.; Ma, X.; Yu, Q.; Yu, X.; Liu, Y.; Nie, C.; Zhang, Y.; Xing, F. The Regulatory Mechanism of Water Activities on Aflatoxins Biosynthesis and Conidia Development, and Transcription Factor AtfB Is Involved in This Regulation. Toxins 2021, 13, 431. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060431
Ma L, Li X, Ma X, Yu Q, Yu X, Liu Y, Nie C, Zhang Y, Xing F. The Regulatory Mechanism of Water Activities on Aflatoxins Biosynthesis and Conidia Development, and Transcription Factor AtfB Is Involved in This Regulation. Toxins. 2021; 13(6):431. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060431
Chicago/Turabian StyleMa, Longxue, Xu Li, Xiaoyun Ma, Qiang Yu, Xiaohua Yu, Yang Liu, Chengrong Nie, Yinglong Zhang, and Fuguo Xing. 2021. "The Regulatory Mechanism of Water Activities on Aflatoxins Biosynthesis and Conidia Development, and Transcription Factor AtfB Is Involved in This Regulation" Toxins 13, no. 6: 431. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060431