Identification and Functional Characterization of the Gene Cluster Responsible for Fusaproliferin Biosynthesis in Fusarium proliferatum

 , , , , , , and
, , , , , , and
Abstract
:
1. Introduction
2. Results
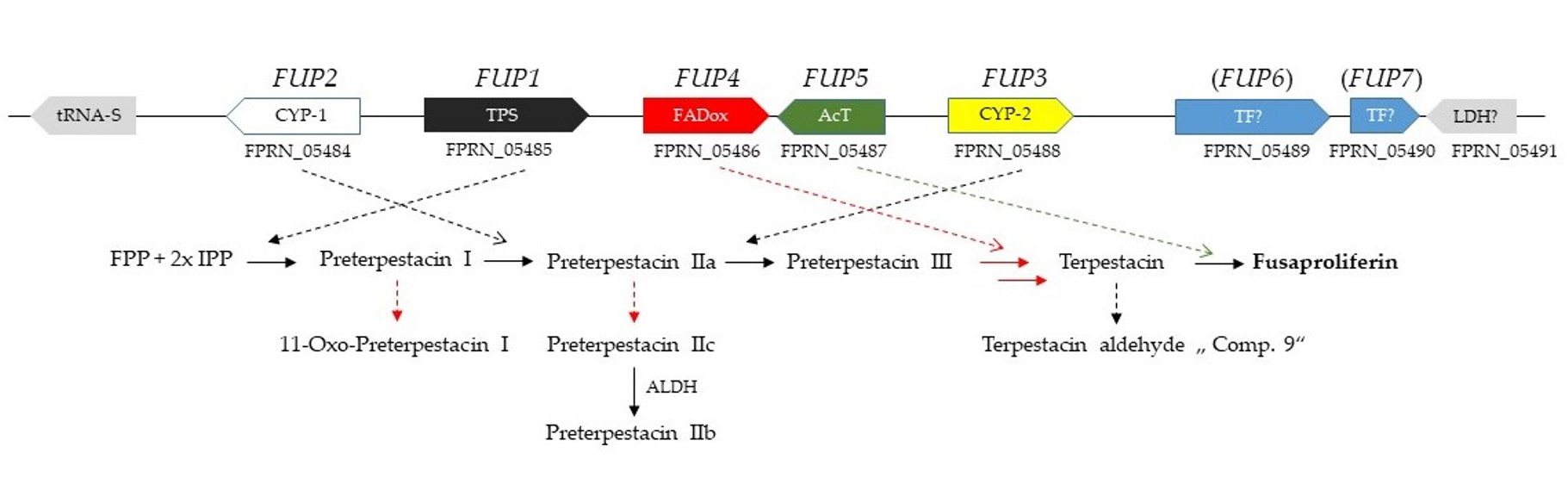
2.1. Bioinformatic Identification of the Terpenoid Synthase Candidate Gene
2.2. Functional Testing of FPRN_05485
2.3. Functional Analysis of the FUP Gene Cluster

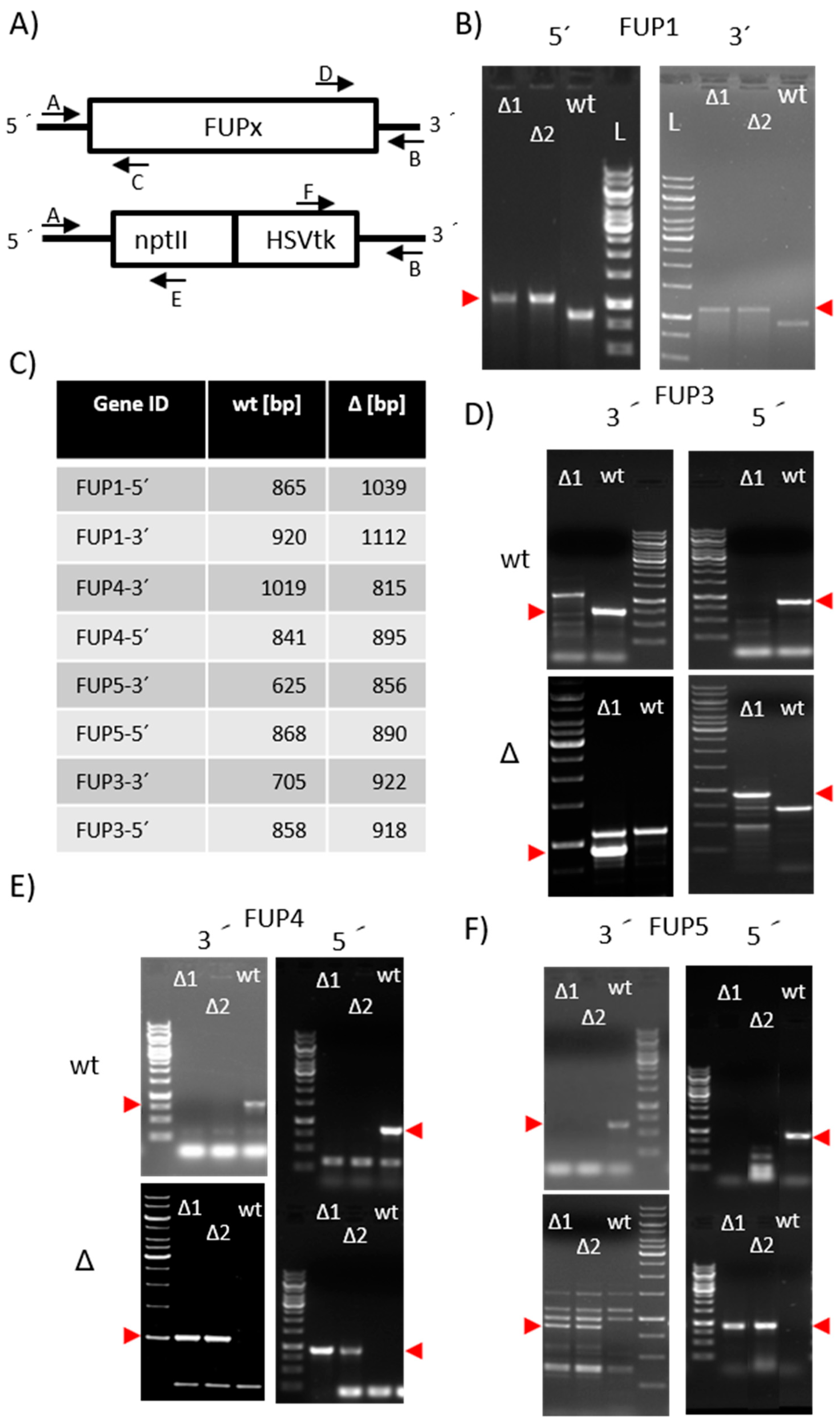
2.3.1. Preparation of Knock-Out Strains
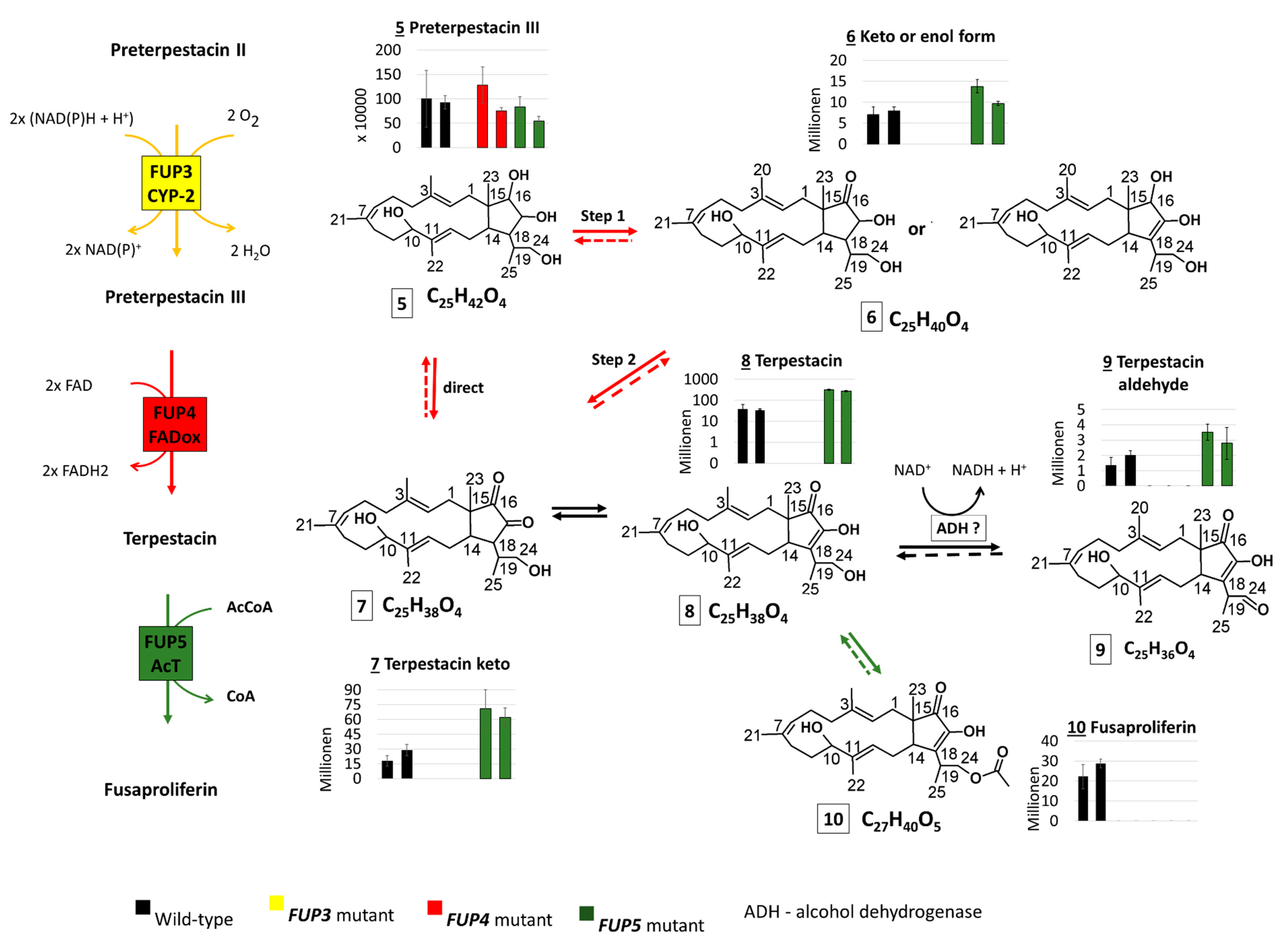
2.3.2. Search for Predicted FUP Intermediates and Structurally Related Compounds
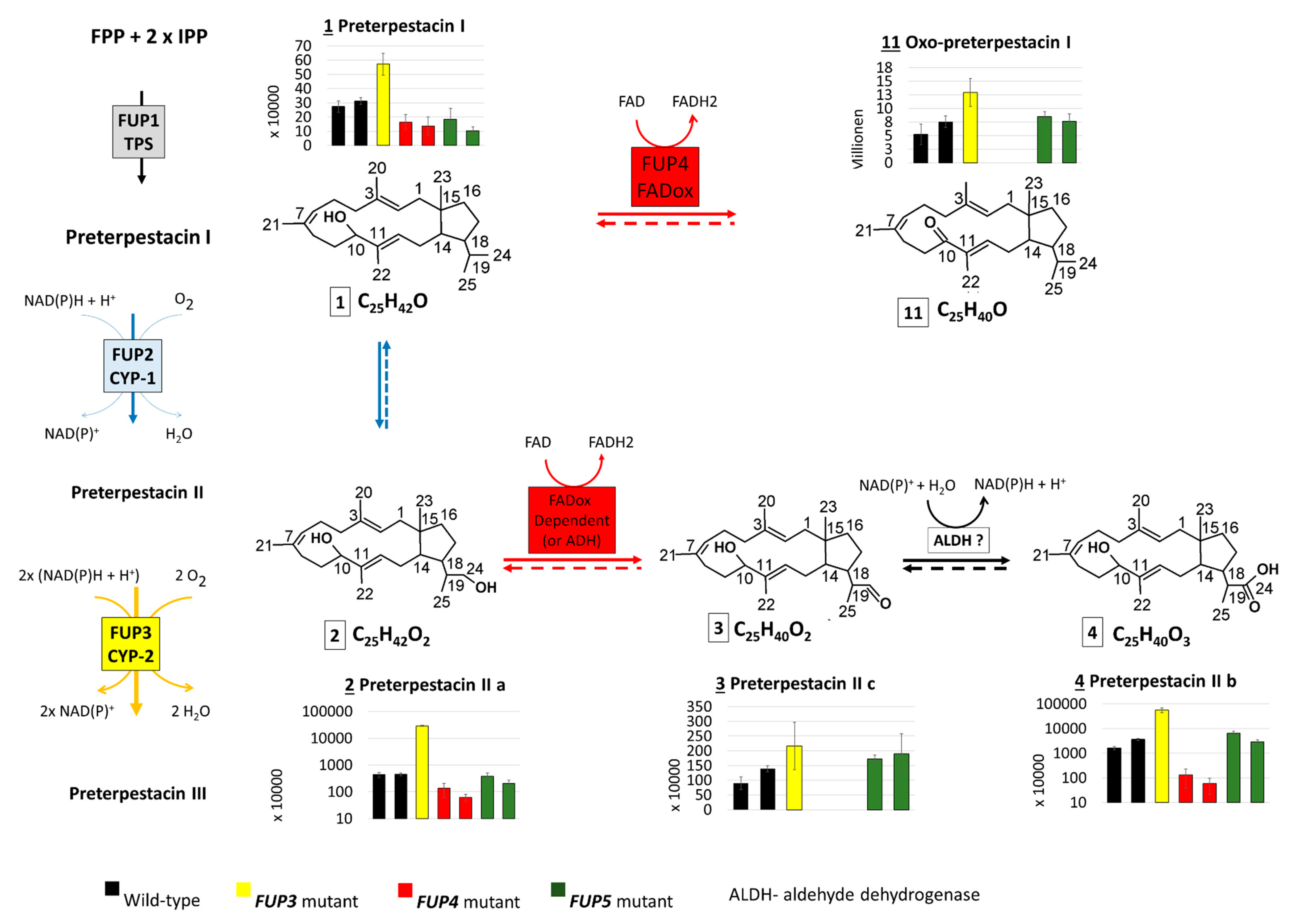
2.3.3. Abundance of FUP and Pathway Precursors in Knock-Out Strains
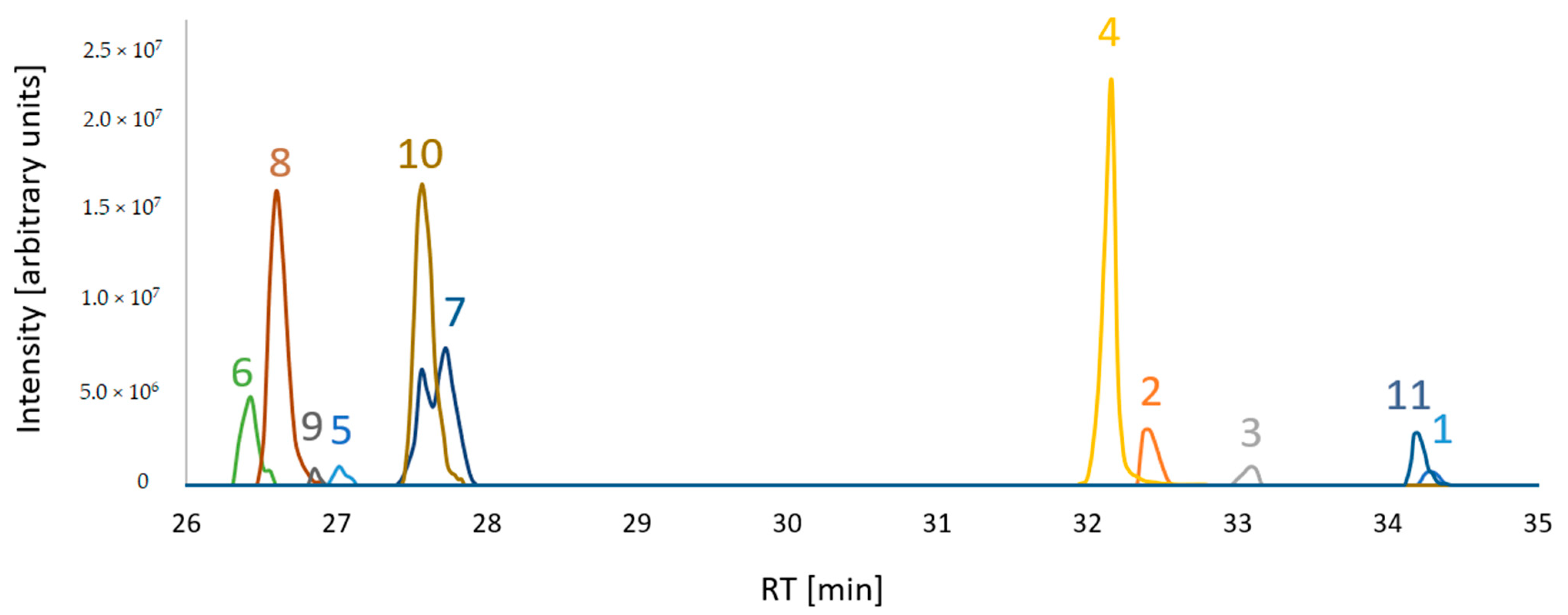
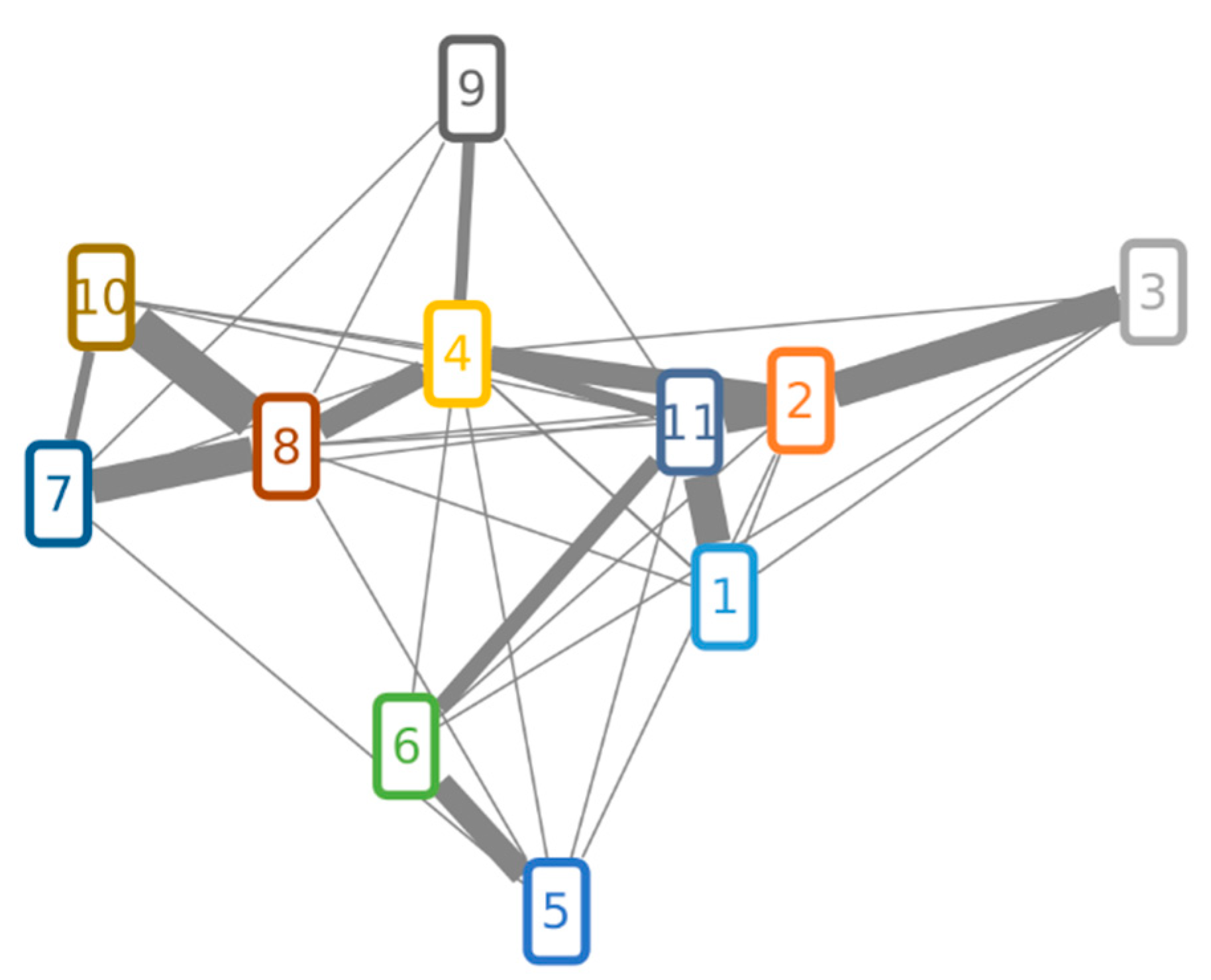
2.3.4. Structure Similarity Determination of Suspect Compounds with Identified Terpestacin and FUP via LC-HRMS/MS
3. Discussion
3.1. The Proposed Fusaproliferin Biosynthetic Pathway in F. proliferatum
3.2. Possible Reasons for the Inability to Obtain a FUP2 (FPRN_05484) Mutant
3.3. Significantly Higher Terpestacin in the fup5 Mutant Strain
3.4. Perspectives and Open Questions
4. Material and methods
4.1. Strains
4.2. Preparation of Knock-Out Strains
4.3. Toxin Test and Extraction of Secondary Metabolites
4.4. LC-HRMS/MS-Based Target Screening and Suspect Analysis of Intermediates of Fusaproliferin Biosynthetic Pathway
4.4.1. Chemicals and Standards
4.4.2. Purification Fusaproliferin and Terpestacin
4.4.3. LC-HRMS/MS Measurement
Liquid Chromatography
Mass Spectrometry
4.4.4. Feature-Based Molecular Networking
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging Mycotoxins: Beyond Traditionally Determined Food Contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef]
- Santini, A.; Meca, G.; Uhlig, S.; Ritieni, A. Fusaproliferin, beauvericin and enniatins: Occurrence in food—A review. World Mycotoxin J. 2012, 5, 71–81. [Google Scholar] [CrossRef]
- Ritieni, A.; Monti, S.M.; Randazzo, G.; Logrieco, A.; Moretti, A.; Peluso, G.; Ferracane, R.; Fogliano, V. Teratogenic effects of fusaproliferin on chicken embryos. J. Agric. Food Chem. 1997, 45, 3039–3043. [Google Scholar] [CrossRef]
- Munkvold, G.; Stahr, H.M.; Logrieco, A.; Moretti, A.; Ritieni, A. Occurrence of fusaproliferin and beauvericin in Fusarium-contaminated livestock feed in Iowa? Appl. Environ. Microb. 1998, 64, 3923–3926. [Google Scholar] [CrossRef] [Green Version]
- Fumero, M.V.; Sulyok, M.; Chulze, S. Ecophysiology of Fusarium temperatum isolated from maize in Argentina. Food Addit. Contam. A 2016, 33, 147–156. [Google Scholar] [CrossRef]
- Ritieni, A.; Fogliano, V.; Randazzo, G.; Scarallo, A.; Logrieco, A.; Moretti, A.; Mannina, L.; Bottalico, A. Isolation and characterization of fusaproliferin, a new toxic metabolite from Fusarium proliferatum. Nat. Toxins 1995, 3, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Oka, M.; Iimura, S.; Tenmyo, O.; Sawada, Y.; Sugawara, M.; Ohkusa, N.; Yamamoto, H.; Kawano, K.; Hu, S.L.; Fukagawa, Y.; et al. Terpestacin, a New Syncytium Formation Inhibitor from Arthrinium Sp. J. Antibiot. 1993, 46, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Tatsuta, K.; Masuda, N. The first total synthesis of natural (+)-terpestacin, syncytium formation inhibitor. J. Antibiot. 1998, 51, 602–606. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, B.; Schmidtke, M.; Dorfelt, H.; Kleinwachter, P.; Grafe, U. (-)-terpestacin and L-tenuazonic acid, inducers of pigment and aerial mycelium formation by Fusarium culmorum JP 15. J. Basic Microb. 2001, 41, 179–183. [Google Scholar] [CrossRef]
- Nihashi, Y.; Lim, C.H.; Tanaka, C.; Miyagawa, H.; Ueno, T. Phytotoxic sesterterpene, 11-epiterpestacin, from Bipolaris sorokiniana NSDR-011. Biosci. Biotech. Bioch. 2002, 66, 685–688. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.; Jamison, T.F. Enantioselective synthesis of (-)-terpestacin and structural revision of siccanol using catalytic stereoselective fragment couplings and macrocyclizations. J. Am. Chem. Soc. 2004, 126, 10682–10691. [Google Scholar] [CrossRef]
- Myers, A.G.; Siu, M.; Ren, F. Enantioselective synthesis of (-)-terpestacin and (-)-fusaproliferin: Clarification of optical rotational measurements and absolute configurational assignments establishes a homochiral structural series. J. Am. Chem. Soc. 2002, 124, 4230–4232. [Google Scholar] [CrossRef]
- Logrieco, A.; Moretti, A.; Fornelli, F.; Fogliano, V.; Ritieni, A.; Caiaffa, M.F.; Randazzo, G.; Bottalico, A.; Macchia, L. Fusaproliferin production by Fusarium subglutinans and its toxicity to Artemia salina, SF-9 insect cells, and IARC/LCL 171 human B lymphocytes. Appl. Environ. Microb. 1996, 62, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Hoque, N.; Hasan, C.M.; Rana, M.S.; Varsha, A.; Sohrab, M.H.; Rahman, K.M. Fusaproliferin, a Fungal Mycotoxin, Shows Cytotoxicity against Pancreatic Cancer Cell Lines. Molecules 2018, 23, 3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosperini, A.; Meca, G.; Font, G.; Ruiz, M.J. Study of the cytotoxic activity of beauvericin and fusaproliferin and bioavailability in vitro on Caco-2 cells. Food Chem. Toxicol. 2012, 50, 2356–2361. [Google Scholar] [CrossRef]
- Pocsfalvi, G.; Ritieni, A.; Randazzo, G.; Dobo, A.; Malorni, A. Interaction of fusarium mycotoxins, fusaproliferin and fumonisin B1, with DNA studied by electrospray ionization mass spectrometry. J. Agric. Food Chem. 2000, 48, 5795–5801. [Google Scholar] [CrossRef]
- Jung, H.J.; Lee, H.B.; Kim, C.J.; Rho, J.R.; Shin, J.; Kwon, H.J. Anti-angiogenic activity of terpestacin, a bicyclo sesterterpene from Embellisia chlamydospora. J. Antibiot. 2003, 56, 492–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.J.; Shim, J.S.; Lee, J.; Song, Y.M.; Park, K.C.; Choi, S.H.; Kim, N.D.; Yoon, J.H.; Mungai, P.T.; Schumacker, P.T.; et al. Terpestacin Inhibits Tumor Angiogenesis by Targeting UQCRB of Mitochondrial Complex III and Suppressing Hypoxia-induced Reactive Oxygen Species Production and Cellular Oxygen Sensing. J. Biol. Chem. 2010, 285, 11584–11595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šrobárová, A.; Čiamporová, M. Is fusaproliferin associated with disease symptoms in maize plants? Cereal Res. Commun. 2006, 34, 1117–1122. [Google Scholar] [CrossRef]
- Masi, M.; Meyer, S.; Gorecki, M.; Pescitelli, G.; Clement, S.; Cimmino, A.; Evidente, A. Phytotoxic Activity of Metabolites Isolated from Rutstroemia sp.n., the Causal Agent of Bleach Blonde Syndrome on Cheatgrass (Bromus tectorum). Molecules 2018, 23, 1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masi, M.; Aloi, F.; Nocera, P.; Cacciola, S.O.; Surico, G.; Evidente, A. Phytotoxic Metabolites Isolated from Neufusicoccum batangarum, the Causal Agent of the Scabby Canker of Cactus Pear (Opuntia ficus-indica L.). Toxins 2020, 12, 126. [Google Scholar] [CrossRef] [Green Version]
- Niehaus, E.M.; Münsterkötter, M.; Proctor, R.H.; Brown, D.W.; Sharon, A.; Idan, Y.; Oren-Young, L.; Sieber, C.M.; Novák, O.; Pěnčík, A.; et al. Comparative “Omics” of the Fusarium fujikuroi Species Complex Highlights Differences in Genetic Potential and Metabolite Synthesis. Genome Biol. Evol. 2016, 8, 3574–3599. [Google Scholar] [CrossRef] [Green Version]
- Tsavkelova, E.; Oeser, B.; Oren-Young, L.; Israeli, M.; Sasson, Y.; Tudzynski, B.; Sharon, A. Identification and functional characterization of indole-3-acetamide-mediated IAA biosynthesis in plant-associated Fusarium species. Fungal Genet. Biol. 2012, 49, 48–57. [Google Scholar] [CrossRef]
- Chiba, R.; Minami, A.; Gomi, K.; Oikawa, H. Identification of Ophiobolin F Synthase by a Genome Mining Approach: A Sesterterpene Synthase from Aspergillus clavatus. Org. Lett. 2013, 15, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Minami, A.; Mandi, A.; Liu, C.W.; Taniguchi, T.; Kuzuyama, T.; Monde, K.; Gomi, K.; Oikawa, H. Genome Mining for Sesterterpenes Using Bifunctional Terpene Synthases Reveals a Unified Intermediate of Di/Sesterterpenes. J. Am. Chem. Soc. 2015, 137, 11846–11853. [Google Scholar] [CrossRef] [PubMed]
- Twaruschek, K.; Sporhase, P.; Michlmayr, H.; Wiesenberger, G.; Adam, G. New Plasmids for Fusarium Transformation Allowing Positive-Negative Selection and Efficient Cre-loxP Mediated Marker Recycling. Front. Microbiol. 2018, 9, 1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narita, K.; Minami, A.; Ozaki, T.; Liu, C.; Kodama, M.; Oikawa, H. Total biosynthesis of antiangiogenic agent (−)-terpestacin by artificial reconstitution of the biosynthetic machinery in Aspergillus oryzae. J. Org. Chem. 2018, 83, 7042–7048. [Google Scholar] [CrossRef]
- Blazenovic, I.; Kind, T.; Ji, J.; Fiehn, O. Software Tools and Approaches for Compound Identification of LC-MS/MS Data in Metabolomics. Metabolites 2018, 8, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nothias, L.F.; Petras, D.; Schmid, R.; Duhrkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Ruttkies, C.; Schymanski, E.L.; Wolf, S.; Hollender, J.; Neumann, S. MetFrag relaunched: Incorporating strategies beyond in silico fragmentation. J. Cheminform 2016, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.; Schmidt, S.; Muller-Hannemann, M.; Neumann, S. In silico fragmentation for computer assisted identification of metabolite mass spectra. BMC Bioinform. 2010, 11, 148. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Hu, H.; Ma, L.; Zhou, Q.; Yu, L.; Zeng, S. Carbon-carbon double-bond reductases in nature. Drug Metab. Rev. 2014, 46, 362–378. [Google Scholar] [CrossRef] [PubMed]
- He, W.J.; Zhang, L.M.; Yi, S.Y.; Tang, X.L.; Yuan, Q.S.; Guo, M.W.; Wu, A.B.; Qu, B.; Li, H.P.; Liao, Y.C. An aldo-keto reductase is responsible for Fusarium toxin-degrading activity in a soil Sphingomonas strain. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.Y.; Bui, D.C.; Lee, Y.; Nam, H.; Jung, S.; Fang, M.; Kim, J.C.; Lee, T.; Kim, H.; Choi, G.J.; et al. Functional characterization of cytochrome P450 monooxygenases in the cereal head blight fungus Fusarium graminearum. Environ. Microbiol. 2017, 19, 2053–2067. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Sarrocco, S.; Masi, M.; Diquattro, S.; Evidente, M.; Vannacci, G.; Evidente, A. Fusaproliferin, Terpestacin and Their Derivatives Display Variable Allelopathic Activity Against Some Ascomycetous Fungi. Chem. Biodivers. 2016, 13, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Zonno, M.C.; Boari, A.; Vurro, M.; Evidente, A. Terpestacin, a toxin produced by Phoma exigua var. heteromorpha, the causal agent of a severe foliar disease of oleander (Nerium oleander L.). Nat. Prod. Res. 2021, 1–7. [Google Scholar] [CrossRef]
- Gao, M.; Yao, S.; Liu, Y.; Yu, H.; Xu, P.; Sun, W.; Pu, Z.; Hou, H.; Bao, Y. Transcriptome Analysis of Tomato Leaf Spot Pathogen Fusarium proliferatum: De novo Assembly, Expression Profiling, and Identification of Candidate Effectors. Int. J. Mol. Sci. 2017, 19, 31. [Google Scholar] [CrossRef] [Green Version]
- Stankovic, S.; Levic, J.; Petrovic, T.; Logrieco, A.; Moretti, A. Pathogenicity and mycotoxin production by Fusarium proliferatum isolated from onion and garlic in Serbia. Eur. J. Plant Pathol. 2007, 118, 165–172. [Google Scholar] [CrossRef]
- Malachova, A.; Sulyok, M.; Beltran, E.; Berthiller, F.; Krska, R. Optimization and validation of a quantitative liquid chromatography-tandem mass spectrometric method covering 295 bacterial and fungal metabolites including all regulated mycotoxins in four model food matrices. J. Chromatogr. A 2014, 1362, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Neumann, N.K.; Lehner, S.M.; Kluger, B.; Bueschl, C.; Sedelmaier, K.; Lemmens, M.; Krska, R.; Schuhmacher, R. Automated LC-HRMS(/MS) approach for the annotation of fragment ions derived from stable isotope labeling-assisted untargeted metabolomics. Anal. Chem. 2014, 86, 7320–7327. [Google Scholar] [CrossRef]
- Adusumilli, R.; Mallick, P. Data Conversion with ProteoWizard msConvert. Methods Mol. Biol. 2017, 1550, 339–368. [Google Scholar] [CrossRef] [PubMed]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 18 September 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Designation | Annotation | NRRL 62905 Gene Protein | ET1 Gene Protein | Protein Identity |
|---|---|---|---|---|
| Prolyl-tRNA synthetase | FPRN_05483 (CVK87167) * | FPRO_05644 CZR39164 | Diff. N-terminus 217/365 (59.5%) | |
| FpFUP2 | Cytochrome P-450 monooxygenase (CYP65A1) | FPRN_05484 CVK87169 | FPRO_05645 CZR39163 | 506/509 (99.4%) |
| FpFUP1 | Geranylgeranyl diphosphate synthase, TPS | FPRN_05485 CVK87172 | FPRO_05646 CZR39162 | 744/753 (98.8%) |
| FpFUP4 | FAD linked oxidase, N-terminal | FPRN_05486 CVK87175 | FPRO_05647 CZR39161 | 506/519 (97.5%) |
| FpFUP5 | Acyltransferase-related | FPRN_05487 CVK87178 | FPRO_05648 CZR39160 | 375/390 (96.2%) |
| FpFUP3 | Cytochrome P450 | FPRN_05488 CVK87181 | FPRO_05649 CZR39159 | 521/529 (98.5%) |
| (FpFUP6) | Transcription factor domain, fungi | FPRN_05489 CVK87185 | FPRO_05650 CZR39158 | 694/700 (99.1%) |
| (FpFUP7) | Zn(II)2Cys6 transcription factor | FPRN_05490 CVK87187 | FPRO_05651 CZR39157 | 235/245 (95.9%) |
| Lactate/Malate dehydrogenase | FPRN_05491 CVK87189 | PRO_05652 CZR39156 | 318/325 (97.8%) |
| Fragment | Primer (5´–3´) | Length (bp) |
|---|---|---|
| FUP1-5´ | TAATGGCCGCATAGGCCGCGTCAACAGCTGCTCCC | 620 |
| AAACTAGTCGGTTAGTCTTTGCGAGCTCTCC | ||
| FUP1-3´ | AATGTCGACGCCTCATTTACACGGACTACAAGGTC | 593 |
| ACAAGCTTGAAATGTGCAGCATGGCGGTG | ||
| FUP2-5´ | TAATGGCCGCATAGGCCTTGCATTATCCTGTTACCTTACTC | 640 |
| AAACTAGTCGGTTAGTCTTTGCGAGCTCTCC | ||
| FUP2-3´ | AATGTCGACGCCTCATTTACACGGACTACAAGGTC | 576 |
| ACAAGCTTGAAATGTGCAGCATGGCGGTG | ||
| FUP3-5´ | TAATGGCCGCATAGGCCGCAGGTTTAGCTCTGCCTGTAGG | 732 |
| AAACTAGTCAAGTCGTCTTGAGATGATGACAAGCTC | ||
| FUP3-3´ | AATGTCGACGCTCTTTGAGACGACCGCTG | 599 |
| ACAAGCTTCGAGGCAGAAGAGGGAGTAACC | ||
| FUP4-5´ | TAATGGCCGCATAGGCCGTAAACGCCACCATGCTCAGC | 650 |
| AAACTAGTGCGAAACCAAGGATTCTGAATGCCG | ||
| FUP4-3´ | AATGTCGACGTTCACGACAGCCACCTTCAGG | 671 |
| ACAAGCTTGACCGCATTCATAATTGGGCAA | ||
| FUP5-5´ | TAATGGCCGCATAGGCCGCCGTTGAATTACCCAGATCCCA | 609 |
| AAACTAGTGTTGAGAGTACAGTACAGGTGCAATG | ||
| FUP5-3´ | AATGTCGACCCTGAGCAGATTCTGGAACCGC | 539 |
| ACAAGCTTCCTCATATCCACAACAATACACTAGATTGCC |
| Region | Outside (5’–3’) | Gene specific (5’–3’) |
|---|---|---|
| FUP1-5´ | GATGGGGCGTCAGTTCTTGAGATCG | CCGTCTCTTGAGCTTCCTCTGCC |
| FUP1-3´ | CGCCCTATCAAGCCAGAGTGCC | GAGCACAGCTACGGAGACACCAG |
| FUP4-5´ | GCATAACGACCTCTCTATAGG | GGAGCAAGGAGGACACTGG |
| FUP4-3´ | CGCACCTGGACTTGTGACC | GTCACGTCACTATCGGACCTTG |
| FUP5-5´ | CGTCGGCAACTGACCCTAACC | CAAGACCAGAATCGGAGTAGTGAGC |
| FUP5-3´ | CAAGATCTAATGGTGATCTTC | GTCCACCCAGTAAACGGACCTTG |
| FUP3-5´ | GCGTCTGACATGCCGAGAGATG | CCATCGAACGGAAATGGTATTGGCC |
| FUP3-3´ | CGTTCTGGACTTTGCTGGGTAC | GGGTTTACGGGACGGAGGAAG |
| Nr. | Molecular Formula | Monoisotopic Mass | Retention Time | Adduct Type | m/z | Compound Annotated/Identified |
|---|---|---|---|---|---|---|
| 1 | C25H42O | 358.3236 | 34.28 | [M−H2O+H]+ | 341.3203 | Preterpestacin I |
| [M+H]+ | 359.3308 | |||||
| [M+Na]+ | 381.3128 | |||||
| [M+NH4]+ | 376.3574 | |||||
| 2 | C25H42O2 | 374.3185 | 32.44 | [M−H2O+H]+ | 357.3152 | Preterpestacin II a |
| [M+H]+ | 375.3258 | |||||
| [M+Na]+ | 397.3077 | |||||
| [M+NH4]+ | 392.3523 | |||||
| [M+HCOO]− | 419.3167 | |||||
| [M+Cl]− | 409.2868 | |||||
| 3 | C25H40O2 | 372.3028 | 33.05 | [M−H2O+H]+ | 355.2995 | Preterpestacin II c |
| [M+H]+ | 373.3101 | |||||
| [M+Na]+ | 395.2921 | |||||
| [M+NH4]+ | 390.3367 | |||||
| 4 | C25H40O3 | 388.2977 | 32.15 | [M−H2O+H]+ | 371.2945 | Preterpestacin II b |
| [M+Na]+ | 411.2870 | |||||
| [M−H]− | 387.2905 | |||||
| [M+HCOO]− | 433.2959 | |||||
| [M+Cl]− | 423.2660 | |||||
| 5 | C25H42O4 | 406.3083 | 27.02 | [M−H2O+H]+ | 389.3050 | Preterpestacin III |
| [M+Na]+ | 429.2975 | |||||
| [M+HCOO]− | 451.3065 | |||||
| 6 | C25H40O4 | 404.2927 | 26.4 | [M−H2O+H]+ | 387.2894 | Two tautomers keto or enol |
| [M+Na]+ | 427.2819 | |||||
| [M−H]− | 403.2854 | |||||
| [M+HCOO]− | 449.2909 | |||||
| 7 | C25H38O4 | 402.2770 | 27.73 | [M−H2O+H]+ | 385.2737 | Terpestacin keto from |
| [M+Na]+ | 425.2662 | |||||
| [M−H]- | 401.2697 | |||||
| [M+HCOO]− | 447.2752 | |||||
| [M+Cl]− | 437.2453 | |||||
| 8 | C25H38O4 | 402.2770 | 26.6 | [M−H2O+H]+ | 385.2737 | Terpestacin enol form TPC |
| [M+H]+ | 403.2843 | |||||
| [M+Na]+ | 425.2662 | |||||
| [M−H]− | 401.2697 | |||||
| [M+HCOO]- | 447.2752 | |||||
| [M+Cl]− | 437.2453 | |||||
| 9 | C25H36O4 | 400.2614 | 26.87 | [M−H2O+H]+ | 383.2581 | Terpestacin aldehyde |
| [M+H]+ | 401.2686 | |||||
| [M−H]− | 399.2541 | |||||
| 10 | C27H40O5 | 444.2876 | 27.57 | [M−H2O+H]+ | 427.2843 | Fusaproliferin FUP |
| [M+H]+ | 445.2949 | |||||
| [M+Na]+ | 467.2768 | |||||
| [M+NH4]+ | 462.3214 | |||||
| [M−H]− | 443.2803 | |||||
| 11 | C25H40O | 356.3079 | 34.2 | [M−H2O+H]+ | 339.3046 | Oxo-preterpestacin I |
| [M+H]+ | 357.3152 | |||||
| [M+Na]+ | 379.2971 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ćeranić, A.; Svoboda, T.; Berthiller, F.; Sulyok, M.; Samson, J.M.; Güldener, U.; Schuhmacher, R.; Adam, G. Identification and Functional Characterization of the Gene Cluster Responsible for Fusaproliferin Biosynthesis in Fusarium proliferatum. Toxins 2021, 13, 468. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13070468
Ćeranić A, Svoboda T, Berthiller F, Sulyok M, Samson JM, Güldener U, Schuhmacher R, Adam G. Identification and Functional Characterization of the Gene Cluster Responsible for Fusaproliferin Biosynthesis in Fusarium proliferatum. Toxins. 2021; 13(7):468. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13070468
Chicago/Turabian StyleĆeranić, Asja, Thomas Svoboda, Franz Berthiller, Michael Sulyok, Jonathan Matthew Samson, Ulrich Güldener, Rainer Schuhmacher, and Gerhard Adam. 2021. "Identification and Functional Characterization of the Gene Cluster Responsible for Fusaproliferin Biosynthesis in Fusarium proliferatum" Toxins 13, no. 7: 468. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13070468