Proteo-Trancriptomic Analyses Reveal a Large Expansion of Metalloprotease-Like Proteins in Atypical Venom Vesicles of the Wasp Meteorus pulchricornis (Braconidae)

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
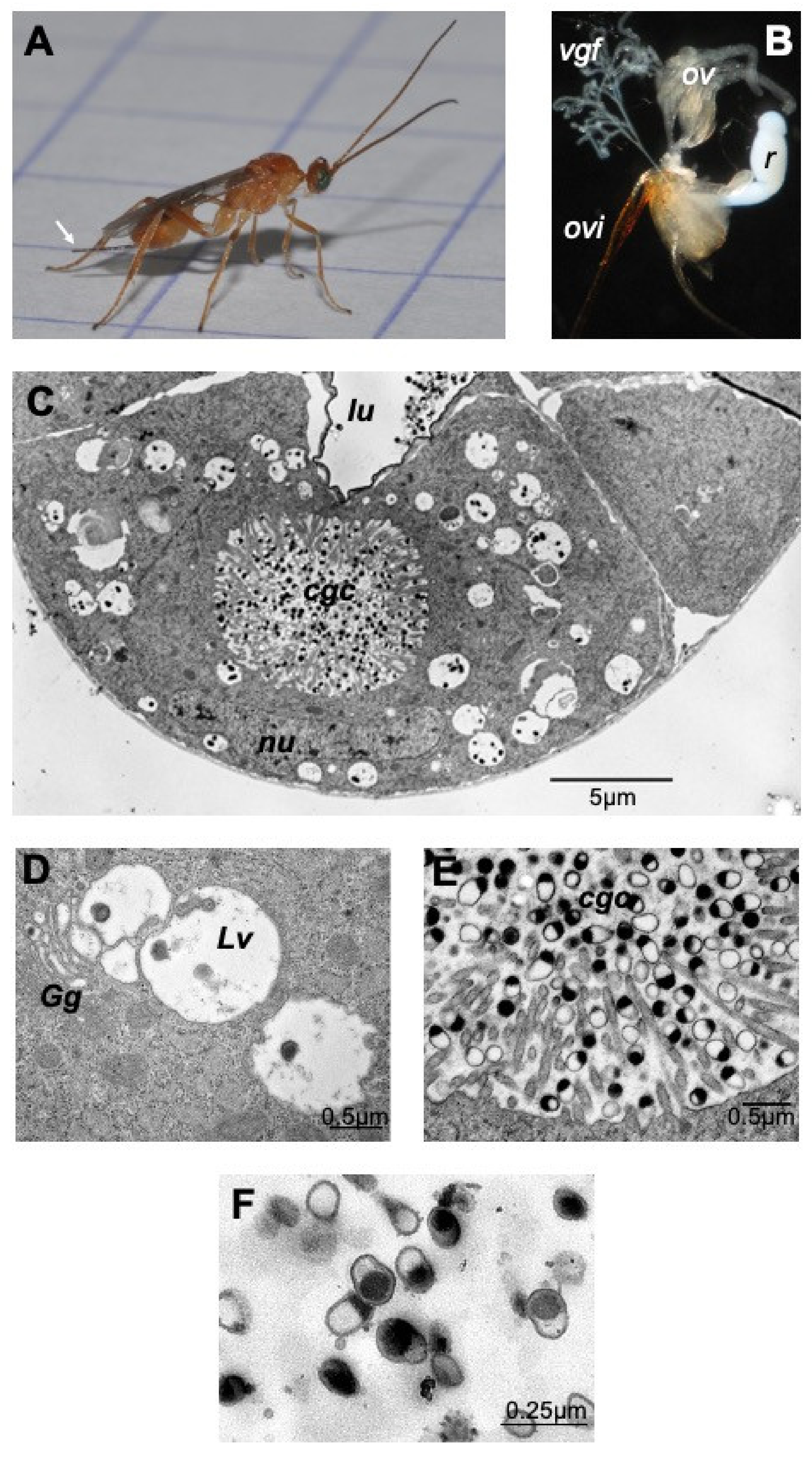
2.1. M. pulchricornis Venom Gland and MpVLPs Secretion
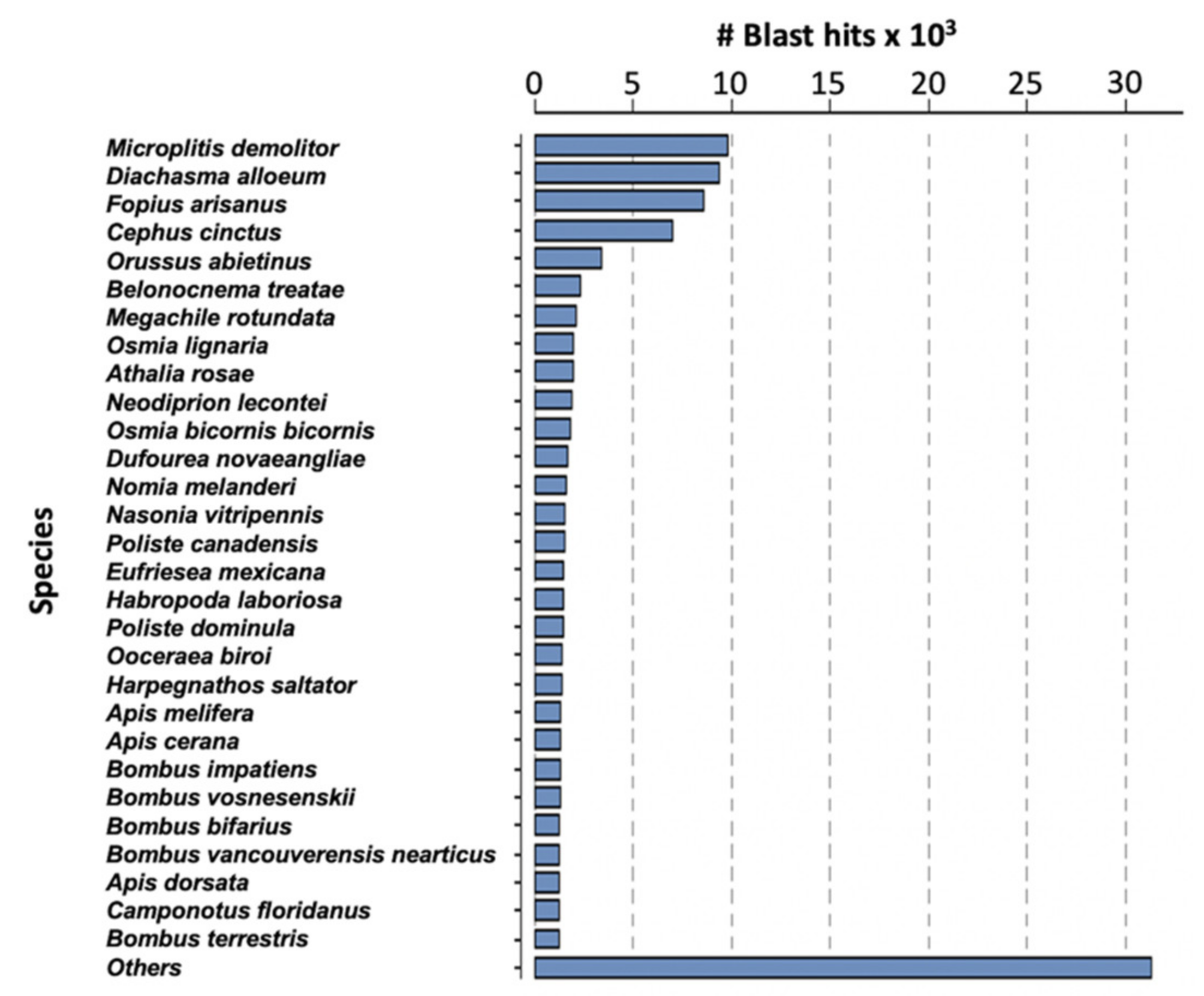
2.2. Analysis of M. pulchricornis Venom Gland Transcriptome
2.3. Search for M. pulchricornis Genes a Priori Preferentially Transcribed in the Venom Gland
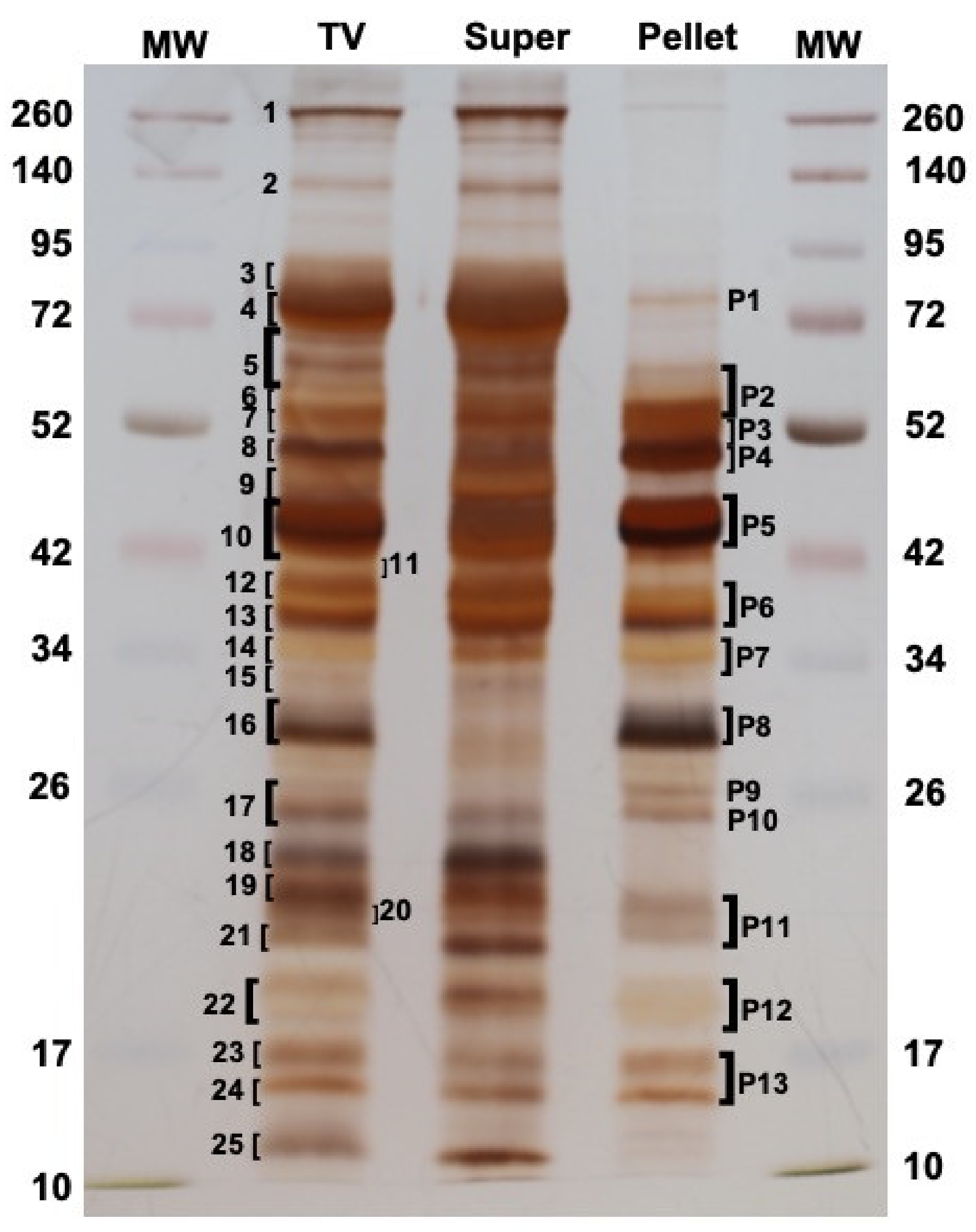
2.4. Proteomic Analysis
2.5. The MpVLPs Proteins
2.6. The Venom Soluble Proteins
2.7. The Most Represented Venom Proteins: Metalloproteases and DUF4803 Proteins
2.7.1. The Venom Metalloproteases
2.7.2. The DUF4803 Containing Proteins
2.8. Other Venom Proteins of Interest
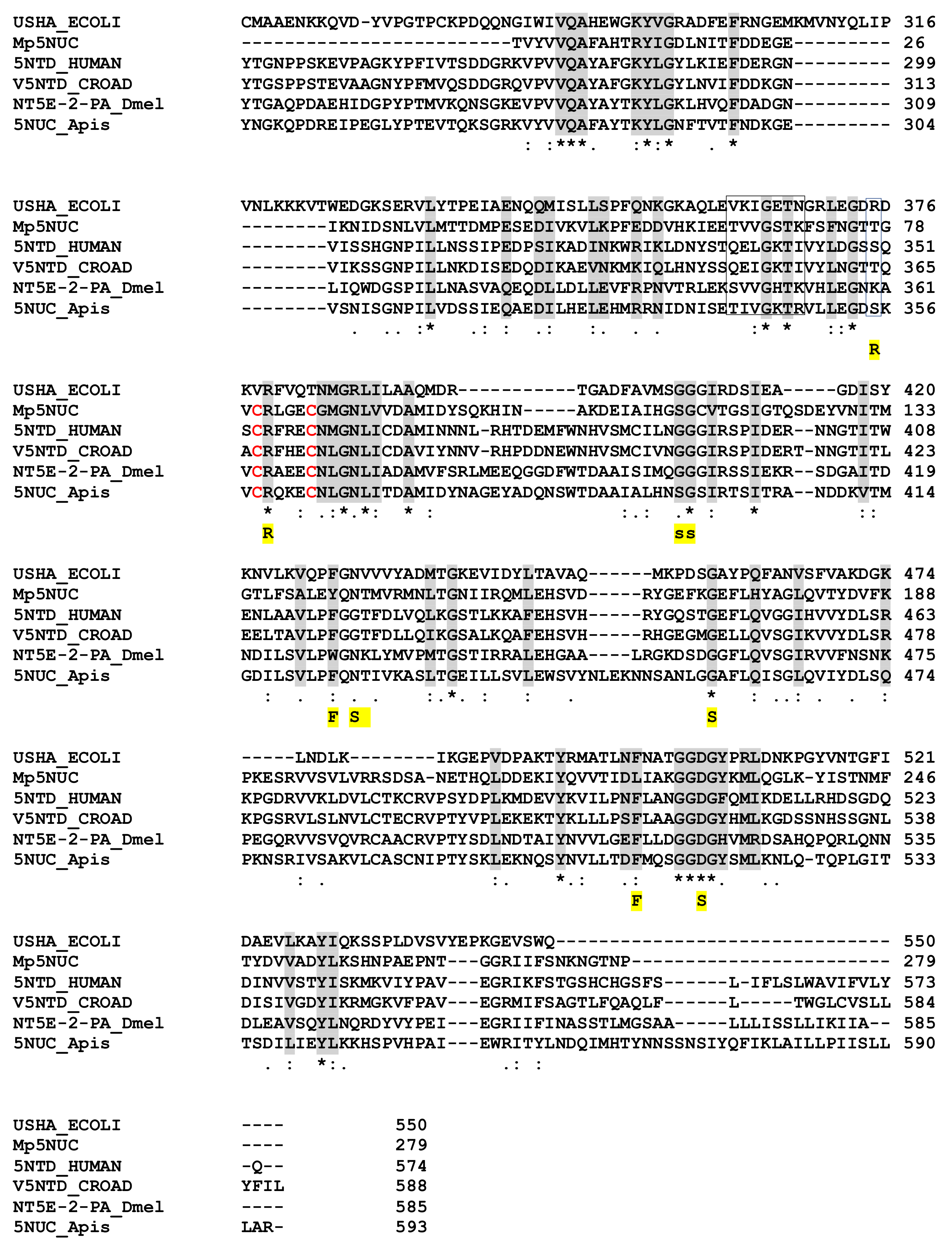
2.8.1. The 5′ Nucleotidase (5NUC)
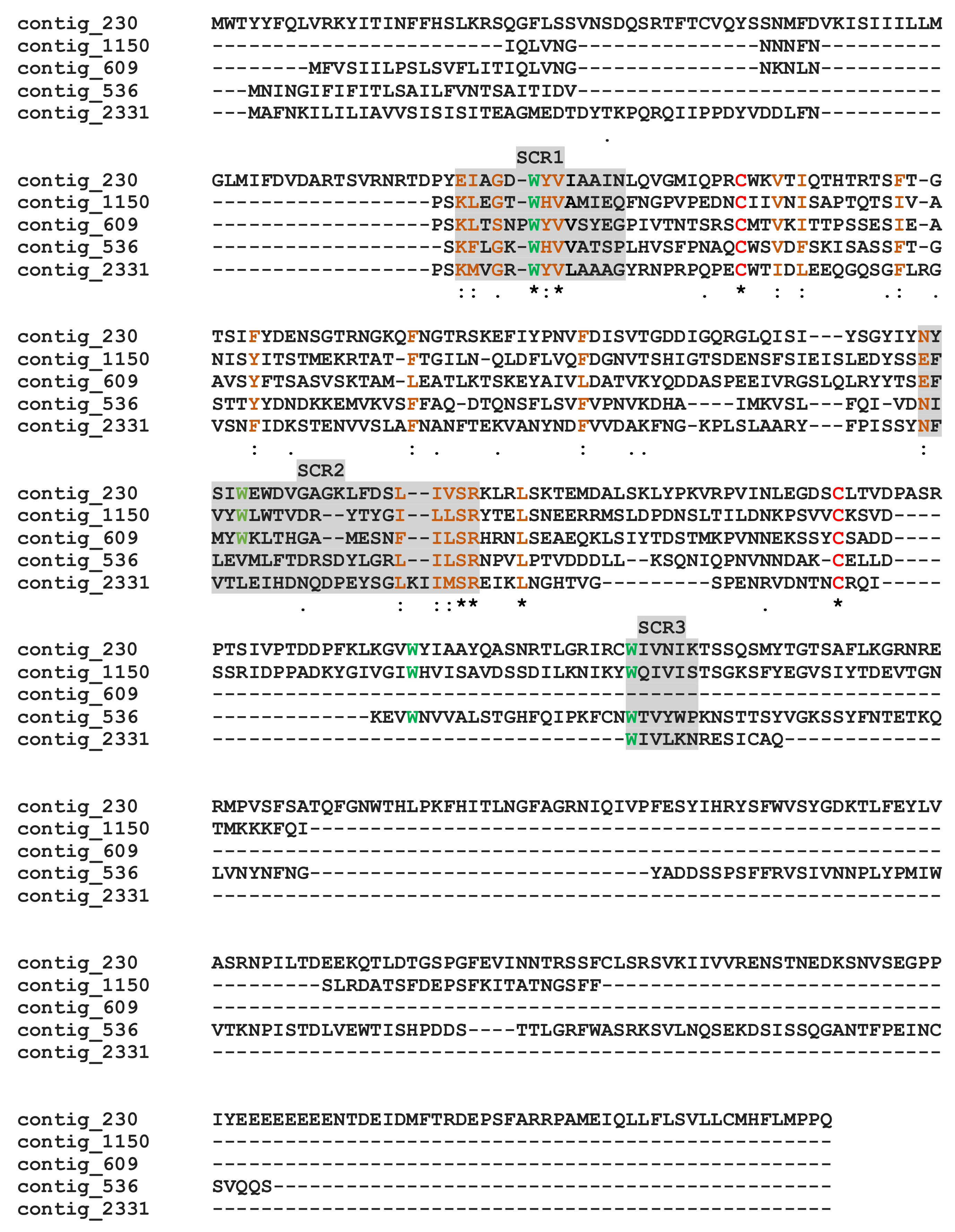
2.8.2. The Calycin/Lipocalin Proteins
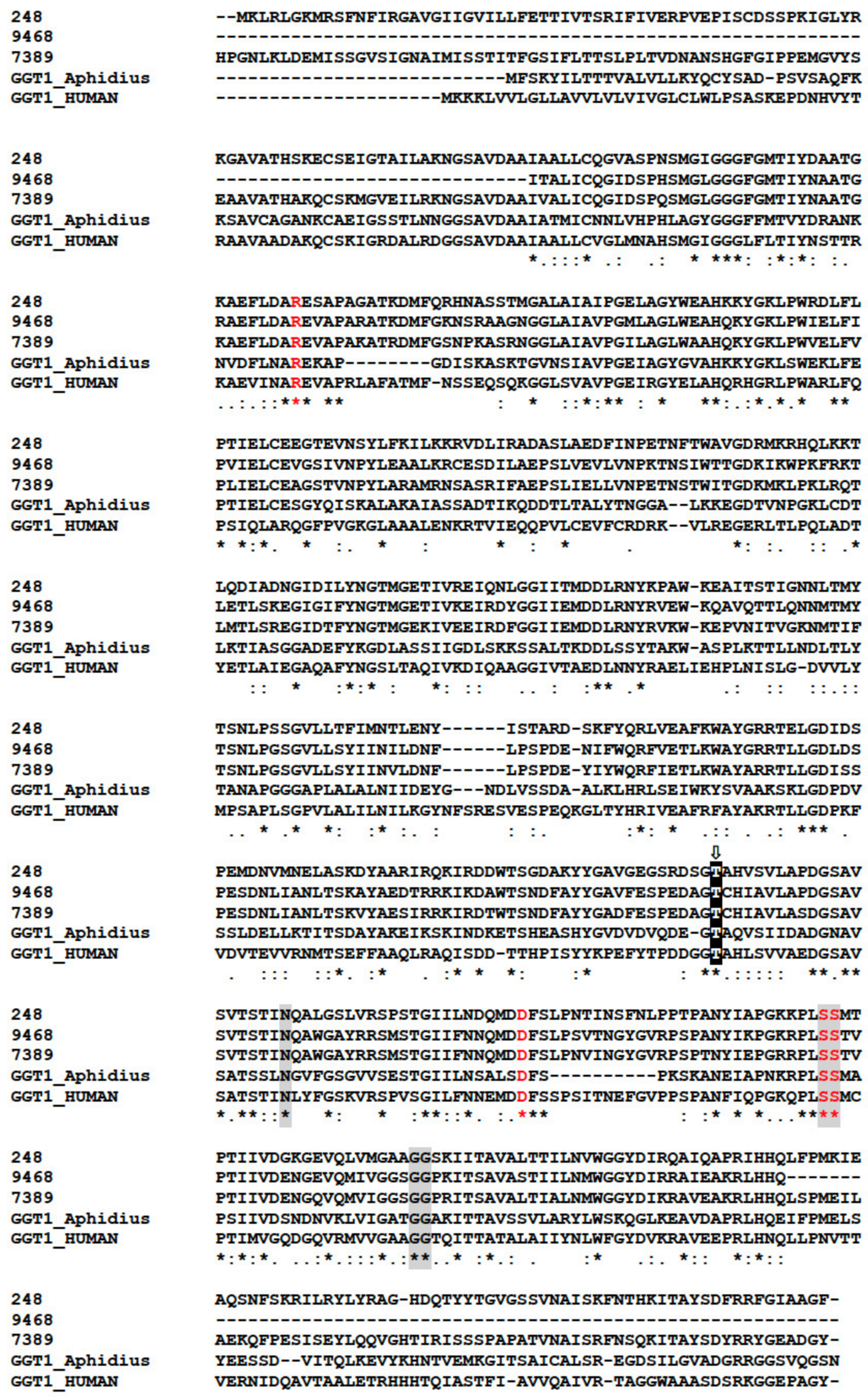
2.8.3. The GGTs
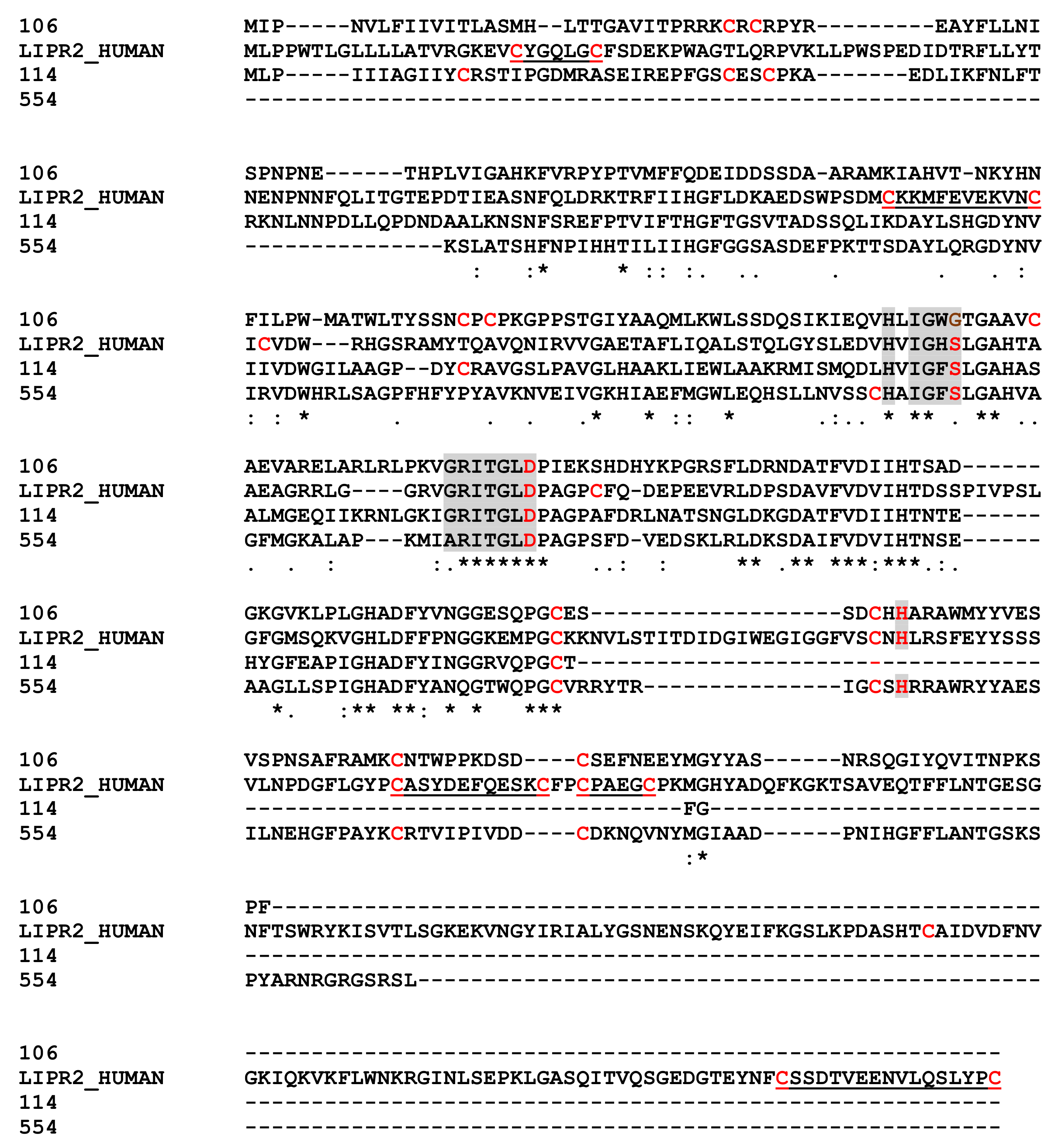
2.8.4. Pancreatic Lipase Like Proteins
2.8.5. Hyaluronidase
2.8.6. CAP Domains Protein
2.8.7. The SERPIN
3. Conclusions
4. Materials and Methods
4.1. Insects
4.2. Transmission Electron Microscopy
4.3. Sequencing and Sequence Analyses
4.4. Venom and VLPs Proteomic
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berry, J.A.; Walker, G.P. Meteorus Pulchricornis (Wesmael) (Hymenoptera: Braconidae: Euphorinae): An Exotic Polyphagous Parasitoid in New Zealand. N. Z. J. Zool. 2004, 31, 33–44. [Google Scholar] [CrossRef]
- Hartnett, D.E.; MacDonald, F.H.; Martin, N.A.; Walker, G.P.; Ward, D.F. A Survey of the Adventive Parasitoid Meteorus Pulchricornis (Hymenoptera: Braconidae) and Native Larval Parasitoids of Native Lepidoptera. N. Z. J. Zool. 2018, 61, 326–343. [Google Scholar] [CrossRef]
- Maeto, K. Systematic Studies on the Tribe Meteorini (Hymenoptera, Braconidae) from Japan V. The Pulchricornis Group of Genus Meteorus (1). Jpn. J. Entomol. 1989, 57, 581–595. [Google Scholar]
- Pennacchio, F.; Strand, M.R. Evolution of Developmental Strategies in Parasitic Hymenoptera. Annu. Rev. Entomol. 2006, 51, 233–258. [Google Scholar] [CrossRef]
- Webb, B.; Fisher, T.; Nusawardani, T. The Natural Genetic Engineering of Polydnaviruses. Ann. N. Y. Acad. Sci. 2009, 1178, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Gatti, J.-L.; Schmitz, A.; Colinet, D.; Poirié, M. Parasitoid Viruses. In Unique Attributes of Viruses and Virus-Like Particles Associated with Parasitoids; Beckage, N.E., Drezen, J.-M., Eds.; Academic Press: Cambridge, MA, USA, 2012; Section B; pp. 181–192. ISBN 9780123848581. [Google Scholar]
- Herniou, E.A.; Huguet, E.; Thézé, J.; Bézier, A.; Periquet, G.; Drezen, J.M. When Parasitic Wasps Hijacked Viruses: Genomic and Functional Evolution of Polydnaviruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20130051. [Google Scholar] [CrossRef]
- Bézier, A.; Herbinière, J.; Lanzrein, B.; Drezen, J.-M. Polydnavirus Hidden Face: The Genes Producing Virus Particles of Parasitic Wasps. J. Invertebr. Pathol. 2009, 101, 194–203. [Google Scholar] [CrossRef]
- Volkoff, A.-N.; Jouan, V.; Urbach, S.; Samain, S.; Bergoin, M.; Wincker, P.; Demettre, E.; Cousserans, F.; Provost, B.; Coulibaly, F.; et al. Analysis of Virion Structural Components Reveals Vestiges of the Ancestral Ichnovirus Genome. PLoS Pathog. 2010, 6, e1000923. [Google Scholar] [CrossRef] [Green Version]
- Pichon, A.; Bézier, A.; Urbach, S.; Aury, J.M.; Jouan, V.; Ravallec, M.; Guy, J.; Cousserans, F.; Thézé, J.; Gauthier, J.; et al. Recurrent DNA Virus Domestication Leading to Different Parasite Virulence Strategies. Sci. Adv. 2015, 1, e1501150. [Google Scholar] [CrossRef] [Green Version]
- Burke, G.R.; Walden, K.K.O.; Whitfield, J.B.; Robertson, H.M.; Strand, M.R. Genome Report: Whole Genome Sequence of the Parasitoid Wasp Microplitis DemolitorThat Harbors an Endogenous Virus Mutualist. G3 Genes Genom. Genet. 2003, 8, 2875–2880. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.R.; Burke, G.R. Polydnaviruses: Evolution and Function. Curr. Issues Mol. Biol. 2020, 34, 163–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauthier, J.; Boulain, H.; van Vugt, J.J.F.A.; Baudry, L.; Persyn, E.; Aury, J.-M.; Noel, B.; Bretaudeau, A.; Legeai, F.; Warris, S.; et al. Chromosomal Scale Assembly of Parasitic Wasp Genome Reveals Symbiotic Virus Colonization. Commun. Biol. 2021, 4, 104. [Google Scholar] [CrossRef]
- Suzuki, M.; Miura, K.; Tanaka, T. The Virus-like Particles of a Braconid Endoparasitoid Wasp, Meteorus Pulchricornis, Inhibit Hemocyte Spreading in Its Noctuid Host, Pseudaletia separata. J. Insect Physiol. 2008, 54, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Tanaka, T. Virus-like Particles in Venom of Meteorus Pulchricornis Induce Host Hemocyte Apoptosis. J. Insect Physiol. 2006, 52, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Miura, K.; Tanaka, T. Effects of the Virus-like Particles of a Braconid Endoparasitoid, Meteorus Pulchricornis, on Hemocytes and Hematopoietic Organs of Its Noctuid Host, Pseudaletia separata. Appl. Entomol. Zool. 2009, 44, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Rizki, R.M.; Rizki, T.M. Parasitoid Virus-like Particles Destroy Drosophila Cellular Immunity. Proc. Natl. Acad. Sci. USA 1990, 87, 8388–8392. [Google Scholar] [CrossRef] [Green Version]
- Dupas, S.; Brehelin, M.; Frey, F.; Carton, Y. Immune Suppressive Virus-like Particles in a Drosophila Parasitoid: Significance of Their Intraspecific Morphological Variations. Parasitology 1996, 113, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Heavner, M.E.; Ramroop, J.; Gueguen, G.; Ramrattan, G.; Dolios, G.; Scarpati, M.; Kwiat, J.; Bhattacharya, S.; Wang, R.; Singh, S.; et al. Novel Organelles with Elements of Bacterial and Eukaryotic Secretion Systems Weaponize Parasites of Drosophila. Curr. Biol. 2017, 27, 2869–2877.e6. [Google Scholar] [CrossRef] [PubMed]
- Colinet, D.; Deleury, E.; Anselme, C.; Cazes, D.; Poulain, J.; Azema-Dossat, C.; Belghazi, M.; Gatti, J.-L.; Poirié, M. Extensive Inter- and Intraspecific Venom Variation in Closely Related Parasites Targeting the Same Host: The Case of Leptopilina Parasitoids of Drosophila. Insect Biochem. Mol. Biol. 2013, 43, 601–611. [Google Scholar] [CrossRef]
- Wan, B.; Goguet, E.; Ravallec, M.; Pierre, O.; Lemauf, S.; Volkoff, A.-N.; Gatti, J.-L.; Poirié, M. Venom Atypical Extracellular Vesicles as Interspecies Vehicles of Virulence Factors Involved in Host Specificity: The Case of a Drosophila Parasitoid Wasp. Front. Immunol. 2019, 10, 1688. [Google Scholar] [CrossRef] [Green Version]
- Di Giovanni, D.; Lepetit, D.; Guinet, B.; Bennetot, B.; Boulesteix, M.; Couté, Y.; Bouchez, O.; Ravallec, M.; Varaldi, J. A Behavior-Manipulating Virus Relative as a Source of Adaptive Genes for Drosophila Parasitoids. Mol. Biol. Evol. 2020, 37, 2791–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, J.; Chiu, H.; Oo, T.; Plaza, R.; Hoskins, S.; Govind, S. Biogenesis, Structure, and Immune-Suppressive Effects of Virus-like Particles of a Drosophila Parasitoid, Leptopilina victoriae. J. Insect Physiol. 2005, 51, 181–195. [Google Scholar] [CrossRef]
- Yokoi, K.; Sano, T.; Suzuki, M.; Tanaka, T.; Minakuchi, C.; Miura, K. The Major Constituents of the Venom Gland of a Braconid Endoparasitoid, Meteorus Pulchricornis (Hymenoptera: Braconidae). Appl. Entomol. Zool. 2017, 52, 271–285. [Google Scholar] [CrossRef]
- Ferrarese, R.; Morales, J.; Fimiarz, D.; Webb, B.A.; Govind, S. A Supracellular System of Actin-Lined Canals Controls Biogenesis And Release Of Virulence Factors In Parasitoid Venom Glands. J. Exp. Biol. 2009, 212, 2261–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, S.; Liao, C.-W.; Zheng, Y.; Zhou, Y.; Xu, Y.; Song, W.-M.; He, P.; Zhang, J.; Wu, F.-A. Candidate Chemosensory Genes Identified in the Endoparasitoid Meteorus Pulchricornis (Hymenoptera: Braconidae) by Antennal Transcriptome Analysis. Comp. Biochem. Physiol. D Genom. Proteom. 2017, 22, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond Chemoreception: Diverse Tasks of Soluble Olfactory Proteins in Insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrenk, S.; Cenzi, C.; Bertalot, T.; Conconi, M.T.; Liddo, R.D. Structural and Functional Failure of Fibrillin-1 in Human Diseases (Review). Int. J. Mol. Med. 2018, 41, 1213–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handke, B.; Poernbacher, I.; Goetze, S.; Ahrens, C.H.; Omasits, U.; Marty, F.; Simigdala, N.; Meyer, I.; Wollscheid, B.; Brunner, E.; et al. The Hemolymph Proteome of Fed and Starved Drosophila Larvae. PLoS ONE 2013, 8, e67208. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Dong, Z.; Wang, D.; Wu, Y.; Song, Q.; Gu, P.; Zhao, P.; Xia, Q. Proteomics of Larval Hemolymph in Bombyx Mori Reveals Various Nutrient-Storage and Immunity-Related Proteins. Amino Acids 2014, 46, 1021–1031. [Google Scholar] [CrossRef]
- Bajgar, A.; Dolezal, T. Extracellular Adenosine Modulates Host-Pathogen Interactions through Regulation of Systemic Metabolism during Immune Response in Drosophila. PLoS Pathog. 2018, 14, e1007022. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.A.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The Toxicogenomic Multiverse: Convergent Recruitment of Proteins into Animal Venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häse, C.C.; Finkelstein, R.A. Bacterial Extracellular Zinc-Containing Metalloproteases. Microbiol. Rev. 1993, 57, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.; Escalante, T.; Rucavado, A.; Herrera, C. Hemorrhage Caused by Snake Venom Metalloproteinases: A Journey of Discovery and Understanding. Toxins 2016, 8, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, J.M.; Rucavado, A. Snake Venom Metalloproteinases: Their Role in the Pathogenesis of Local Tissue Damage. Biochimie 2000, 82, 841–850. [Google Scholar] [CrossRef]
- Consoli, F.L.; Vinson, S.B. Host Regulation and the Embryonic Development of the Endoparasitoid Toxoneuron Nigriceps (Hymenoptera: Braconidae). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 137, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Price, D.R.G.; Bell, H.A.; Hinchliffe, G.; Fitches, E.; Weaver, R.; Gatehouse, J.A. A Venom Metalloproteinase from the Parasitic Wasp Eulophus Pennicornis Is Toxic towards Its Host, Tomato Moth (Lacanobia Oleracae). Insect Mol. Biol. 2009, 18, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Danneels, E.L.; Rivers, D.B.; Graaf, D.C. De Venom Proteins of the Parasitoid Wasp Nasonia vitripennis: Recent Discovery of an Untapped Pharmacopee. Toxins 2010, 2, 494. [Google Scholar] [CrossRef] [Green Version]
- Danneels, E.L.; Vaerenbergh, M.V.; Debyser, G.; Devreese, B.; Graaf, D.C. de Honeybee Venom Proteome Profile of Queens and Winter Bees as Determined by a Mass Spectrometric Approach. Toxins 2015, 7, 4468. [Google Scholar] [CrossRef] [Green Version]
- Souza, J.S.M.; Lisboa, A.B.P.; Santos, T.M.; Andrade, M.V.S.; Neves, V.B.S.; Teles-Souza, J.; Jesus, H.N.R.; Bezerra, T.G.; Falcão, V.G.O.; Oliveira, R.C.; et al. The Evolution of ADAM Gene Family in Eukaryotes. Genomics 2020, 112, 3108–3116. [Google Scholar] [CrossRef]
- Edwards, D.R.; Handsley, M.M.; Pennington, C.J. The ADAM Metalloproteinases. Mol. Asp. Med. 2008, 29, 258–289. [Google Scholar] [CrossRef]
- Seals, D.F.; Courtneidge, S.A. The ADAMs Family of Metalloproteases: Multidomain Proteins with Multiple Functions. Gene Dev. 2003, 17, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Moura-da-Silva, A.M.; Almeida, M.T.; Portes-Junior, J.A.; Nicolau, C.A.; Gomes-Neto, F.; Valente, R.H. Processing of Snake Venom Metalloproteinases: Generation of Toxin Diversity and Enzyme Inactivation. Toxins 2016, 8, 183. [Google Scholar] [CrossRef]
- Laronha, H.; Caldeira, J. Structure and Function of Human Matrix Metalloproteinases. Cells 2020, 9, 1076. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S. ADAM and ADAMTS Family Proteins and Snake Venom Metalloproteinases: A Structural Overview. Toxins 2016, 8, 155. [Google Scholar] [CrossRef] [Green Version]
- Giorgianni, M.W.; Dowell, N.L.; Griffin, S.; Kassner, V.A.; Selegue, J.E.; Carroll, S.B. The Origin and Diversification of a Novel Protein Family in Venomous Snakes. Proc. Natl. Acad. Sci. USA 2020, 117, 10911–10920. [Google Scholar] [CrossRef] [PubMed]
- Bayes-Genis, A.; Barallat, J.; Richards, A.M. A Test in Context: Neprilysin Function, Inhibition, and Biomarker. J. Am. Coll. Cardiol. 2016, 68, 639–653. [Google Scholar] [CrossRef]
- Turner, A.J.; Isaac, R.E.; Coates, D. The Neprilysin (NEP) Family of Zinc Metalloendopeptidases: Genomics and Function. BioEssays 2001, 23, 261–269. [Google Scholar] [CrossRef]
- Isaac, R.E.; Bland, N.D.; Shirras, A.D. Neuropeptidases and the Metabolic Inactivation of Insect Neuropeptides. Gen. Comp. Endocrinol. 2009, 162, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Bland, N.D.; Pinney, J.W.; Thomas, J.E.; Turner, A.J.; Isaac, R.E. Bioinformatic Analysis of the Neprilysin (M13) Family of Peptidases Reveals Complex Evolutionary and Functional Relationships. BMC Evol. Biol. 2008, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathé-Hubert, H.; Colinet, D.; Deleury, E.; Belghazi, M.; Ravallec, M.; Poulain, J.; Dossat, C.; Poirié, M.; Gatti, J.-L. Comparative Venomics of Psyttalia lounsburyi and P. concolor, two Olive Fruit Fly Parasitoids: A Hypothetical Role for a GH1 β-Glucosidase. Sci. Rep. 2016, 6, 35873. [Google Scholar] [CrossRef] [PubMed]
- Goecks, J.; Mortimer, N.T.; Mobley, J.A.; Bowersock, G.J.; Taylor, J.; Schlenke, T.A. Integrative Approach Reveals Composition of Endoparasitoid Wasp Venoms. PLoS ONE 2013, 8, e64125. [Google Scholar] [CrossRef] [PubMed]
- Cha, W.H.; Lee, D.-W. C-Terminal Conserved Motifs of Neprilysin1 in Cotesia plutellae Are Not Required for Immune Suppression of the Diamondback Moth, Plutella xylostella (L.). J. Asia Pac. Entomol. 2019, 22, 1161–1166. [Google Scholar] [CrossRef]
- Asgari, S.; Reineke, A.; Beck, M.; Schmidt, O. Isolation and Characterization of a Neprilysin-like Protein from Venturia Canescens Virus-like Particles. Insect Mol. Biol. 2002, 11, 477–485. [Google Scholar] [CrossRef]
- Yoon, K.A.; Kim, K.; Kim, W.-J.; Bang, W.Y.; Ahn, N.-H.; Bae, C.-H.; Yeo, J.-H.; Lee, S.H. Characterization of Venom Components and Their Phylogenetic Properties in Some Aculeate Bumblebees and Wasps. Toxins 2020, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.Y.; Tan, C.H.; Chanhome, L.; Tan, N.H. Comparative Venom Gland Transcriptomics of Naja kaouthia (Monocled Cobra) from Malaysia and Thailand: Elucidating Geographical Venom Variation and Insights into Sequence Novelty. PeerJ 2017, 5, e3142. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Yu, S.; Wu, Y.; Shan, B. Effect of Spider Venom on Cell Apoptosis and Necrosis Rates in MCF-7 Cells. DNA Cell Biol. 2007, 26, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Soraia, N.; De Oliveira, U.C.; Nishiyama, M.Y., Jr.; Tashima, A.K.; Da Silva, P.I.J. Presence of a Neprilysin on Avicularia Juruensis (Mygalomorphae: Theraphosidae) Venom. Toxin Rev. 2021, 1–10. [Google Scholar] [CrossRef]
- Becchimanzi, A.; Avolio, M.; Bostan, H.; Colantuono, C.; Cozzolino, F.; Mancini, D.; Chiusano, M.L.; Pucci, P.; Caccia, S.; Pennacchio, F. Venomics of the Ectoparasitoid Wasp Bracon nigricans. BMC Genom. 2020, 21, 34. [Google Scholar] [CrossRef]
- Colinet, D.; Anselme, C.; Deleury, E.; Mancini, D.; Poulain, J.; Azema-Dossat, C.; Belghazi, M.; Tarès, S.; Pennacchio, F.; Poirié, M.; et al. Identification of the Main Venom Protein Components of Aphidius Ervi, a Parasitoid Wasp of the Aphid Model Acyrthosiphon pisum. BMC Genom. 2014, 15, 342. [Google Scholar] [CrossRef] [Green Version]
- Sträter, N. Ecto-5’-Nucleotidase: Structure Function Relationships. Purinergic Signal. 2006, 2, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Aili, S.R.; Touchard, A.; Hayward, R.; Robinson, S.D.; Pineda, S.S.; Lalagüe, H.; Mrinalini, V.I.; Undheim, E.A.B.; Kini, R.M.; Escoubas, P.; et al. An Integrated Proteomic and Transcriptomic Analysis Reveals the Venom Complexity of the Bullet Ant Paraponera clavata. Toxins 2020, 12, 324. [Google Scholar] [CrossRef]
- Péterfi, O.; Boda, F.; Szabó, Z.; Ferencz, E.; Bába, L. Hypotensive Snake Venom Components—A Mini-Review. Molecules 2019, 24, 2778. [Google Scholar] [CrossRef] [Green Version]
- Bhadrapura, L.D.; Cletus, J.M.D. The Pharmacological Role of Nucleotidases in Snake Venoms. Cell Biochem. Funct. 2010, 28, 171. [Google Scholar] [CrossRef] [PubMed]
- Allard, B.; Longhi, M.S.; Robson, S.C.; Stagg, J. The Ectonucleotidases CD39 and CD73: Novel Checkpoint Inhibitor Targets. Immunol. Rev. 2017, 276, 121–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flower, D.R. The Lipocalin Protein Family: Structure and Function. Biochem. J. 1996, 318, 1–14. [Google Scholar] [CrossRef]
- Flower, D.R.; North, A.C.; Sansom, C.E. The Lipocalin Protein Family: Structural and Sequence Overview. Biochim. Biophys. Acta 2000, 1482, 9–24. [Google Scholar] [CrossRef]
- Ganfornina, L.; Sanchez, D.; Flower, L.H.G.; Flower, D.R. The Lipocalin Protein Family: Protein Sequence, Structure and Relationship to the Calycin Superfamily. In Lipocalins; Åkerström, B., Borregaard, N., Flower, D.R., Salie, J.-P., Eds.; CRC Press: Boca Raton, FL, USA, 2006; ISBN 9780429089886. [Google Scholar]
- Ganfornina, M.D.; Gutiérrez, G.; Bastiani, M.; Sánchez, D. A Phylogenetic Analysis of the Lipocalin Protein Family. Mol. Biol. Evol. 2000, 17, 114–126. [Google Scholar] [CrossRef] [Green Version]
- Flower, D.R.; North, A.C.T.; Attwood, T.K. Structure and Sequence Relationships in the Lipocalins and Related Proteins. Protein Sci. 1993, 2, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhu, J.-Y.; Qian, C.; Fang, Q.; Ye, G.-Y. Venom of The Parasitoid Wasp Pteromalus puparum Contains an Odorant Binding Protein. Arch. Insect Biochem. Physiol. 2015, 88, 101–110. [Google Scholar] [CrossRef]
- Mariano, D.O.C.; de Oliveira, Ú.C.; Zaharenko, A.J.; Pimenta, D.C.; Rádis-Baptista, G.; Prieto-da-Silva, Á.R. Bottom-Up Proteomic Analysis of Polypeptide Venom Components of the Giant Ant Dinoponera quadriceps. Toxins 2019, 11, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falabella, P.; Riviello, L.; Caccialupi, P.; Rossodivita, T.; Valente, M.T.; Stradis, M.L.D.; Tranfaglia, A.; Varricchio, P.; Gigliotti, S.; Graziani, F.; et al. A Gamma-Glutamyl Transpeptidase of Aphidius Ervi Venom Induces Apoptosis in the Ovaries of Host Aphids. Insect Biochem. Mol. Biol. 2007, 37, 453–465. [Google Scholar] [CrossRef]
- Battistini, B.; Chailler, P.; Brière, N.; Beaudoin, A.R. Secretion of Gamma-Glutamyltranspeptidase by the Pancreas: Evidence for a Membrane Shedding Process during Exocytosis. Life Sci. 1990, 47, 2435–2441. [Google Scholar] [CrossRef]
- Holmes, R.S.; Cox, L.A. Bioinformatics and Evolution of Vertebrate Pancreatic Lipase and Related Proteins and Genes. J. Data Min. Genom. Proteom. 2012, 3, 111. [Google Scholar] [CrossRef] [Green Version]
- Barbara, S.; Francine, F.; Philippe, G.; Dominique, L.; Paul, B.; Alain, D.C.; Bernard, L.; Robert, V.; Frédéric, C. Human Pancreatic Lipase-Related Protein 2 Is a Galactolipase. Biochemistry 2004, 43, 10138–10148. [Google Scholar] [CrossRef]
- Ye, X.; Yan, Z.; Yang, Y.; Xiao, S.; Chen, L.; Wang, J.; Wang, F.; Xiong, S.; Mei, Y.; Wang, F.; et al. A Chromosome-level Genome Assembly of the Parasitoid Wasp Pteromalus puparum. Mol. Ecol. Resour. 2020, 20, 1384–1402. [Google Scholar] [CrossRef] [PubMed]
- Bordon, K.C.F.; Wiezel, G.A.; Amorim, F.G.; Arantes, E.C. Arthropod Venom Hyaluronidases: Biochemical Properties and Potential Applications in Medicine and Biotechnology. J. Venom. Anim. Toxins 2015, 21, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, J.W. A Brief Review of the Scientific History of Several Lesser-Known Snake Venom Proteins: L-Amino Acid Oxidases, Hyaluronidases and Phosphodiesterases. Toxicon 2013, 62, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Sugahara, K.; Özbek, S. Evolution of Glycosaminoglycans. Commun. Integr. Biol. 2011, 4, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Marković-Housley, Z.; Miglierini, G.; Soldatova, L.; Rizkallah, P.J.; Müller, U.; Schirmer, T. Crystal Structure of Hyaluronidase, a Major Allergen of Bee Venom. Structure 2000, 8, 1025–1035. [Google Scholar] [CrossRef]
- Abraham, A.; Chandler, D.E. Tracing the Evolutionary History of the CAP Superfamily of Proteins Using Amino Acid Sequence Homology and Conservation of Splice Sites. J. Mol. Evol. 2017, 85, 137–157. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, T.; Modahl, C.M.; Maenaka, K.; Aoki-Shioi, N. Cysteine-Rich Secretory Proteins (CRISPs) from Venomous Snakes: An Overview of the Functional Diversity in a Large and Underappreciated Superfamily. Toxins 2020, 12, 175. [Google Scholar] [CrossRef] [Green Version]
- Sunagar, K.; Johnson, W.E.; O’Brien, S.J.; Vasconcelos, V.; Antunes, A. Evolution of CRISPs Associated with Toxicoferan-Reptilian Venom and Mammalian Reproduction. Mol. Biol. Evol. 2012, 29, 1807–1822. [Google Scholar] [CrossRef] [Green Version]
- Blank, S.; Bazon, M.L.; Grosch, J.; Schmidt-Weber, C.B.; Brochetto-Braga, M.R.; Bilò, M.B.; Jakob, T. Antigen 5 Allergens of Hymenoptera Venoms and Their Role in Diagnosis and Therapy of Venom Allergy. Curr. Allergy Asthm. Rep. 2020, 20, 58. [Google Scholar] [CrossRef]
- Sanrattana, W.; Maas, C.; de Maat, S. SERPINs—From Trap to Treatment. Front. Med. 2019, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanost, M.R.; Jiang, H. Clip-Domain Serine Proteases as Immune Factors in Insect Hemolymph. Curr. Opin. Insect Sci. 2015, 11, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colinet, D.; Dubuffet, A.; Cazes, D.; Moreau, S.; Drezen, J.-M.; Poirié, M. A Serpin from the Parasitoid Wasp Leptopilina Boulardi Targets the Drosophila Phenoloxidase Cascade. Dev. Comp. Immunol. 2009, 33, 681–689. [Google Scholar] [CrossRef]
- Lin, Z.; Cheng, Y.; Wang, R.-J.; Du, J.; Volovych, O.; Li, J.-C.; Hu, Y.; Lu, Z.-Y.; Lu, Z.; Zou, Z. A Metalloprotease Homolog Venom Protein from a Parasitoid Wasp Suppresses the Toll Pathway in Host Hemocytes. Front. Immunol. 2018, 9, 2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formesyn, E.M.; Heyninck, K.; de Graaf, D.C. The Role of Serine- and Metalloproteases in Nasonia Vitripennis Venom in Cell Death Related Processes towards a Spodoptera Frugiperda Sf21 Cell Line. J. Insect Physiol. 2013, 59, 795–803. [Google Scholar] [CrossRef]
- Walker, A.A.; Robinson, S.D.; Yeates, D.K.; Jin, J.; Baumann, K.; Dobson, J.; Fry, B.G.; King, G.F. Entomo-Venomics: The Evolution, Biology and Biochemistry of Insect Venoms. Toxicon 2018, 154, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, G.; Holland, S.R.; De Perez-Rasmussen, J.; Jarvis, B.A.; Telander, T.; Wagner, N.; Waring, A.L.; Anast, A.; Davis, B.; Frank, A.; et al. Bioinformatic Analysis Suggests Potential Mechanisms Underlying Parasitoid Venom Evolution and Function. Genomics 2020, 112, 1096–1104. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows–Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. Subgroup, 1000 Genome Project Data Processing, The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Buchfink, B.; Reuter, K.; Drost, H.-G. Sensitive Protein Alignments at Tree-of-Life Scale Using DIAMOND. Nat. Methods 2021, 18, 366–368. [Google Scholar] [CrossRef] [PubMed]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic Orthology Inference for Comparative Genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legeai, F.; Santos, B.F.; Robin, S.; Bretaudeau, A.; Dikow, R.B.; Lemaitre, C.; Jouan, V.; Ravallec, M.; Drezen, J.-M.; Tagu, D.; et al. Genomic Architecture of Endogenous Ichnoviruses Reveals Distinct Evolutionary Pathways Leading to Virus Domestication in Parasitic Wasps. BMC Biol. 2020, 18, 89. [Google Scholar] [CrossRef] [PubMed]
- Dennis, A.B.; Ballesteros, G.I.; Robin, S.; Schrader, L.; Bast, J.; Berghöfer, J.; Beukeboom, L.W.; Belghazi, M.; Bretaudeau, A.; Buellesbach, J.; et al. Functional Insights from the GC-Poor Genomes of Two Aphid Parasitoids, Aphidius Ervi and Lysiphlebus fabarum. BMC Genom. 2020, 21, 376. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-Scale Protein Function Classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-Throughput Functional Annotation and Data Mining with the Blast2GO Suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Morrissey, J.H. Silver Stain for Proteins in Polyacrylamide Gels: A Modified Procedure with Enhanced Uniform Sensitivity. Anal. Biochem. 1981, 117, 307–310. [Google Scholar] [CrossRef]
- Doerr, A. Proteomics Data Reuse with MassIVE-KB. Nat. Methods 2019, 16, 26. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER Web Server: 2018 Update. Nucleic Acids Res. 2018, 46, gky448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; mi Park, Y.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI Search and Sequence Analysis Tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TPMv Range | Number of Predicted Proteins |
|---|---|
| 75,000–10,000 | 15 |
| 9999–5000 | 24 |
| 4999–1000 | 88 |
| 999–500 | 118 |
| 499–100 | 480 |
| 99–50 | 477 |
| 49–25 | 852 |
| 24–10 | 2370 |
| 9–0 | 12,404 |
| CDS ID | Sequence Description * | Species | TPMv | TPMa | TPMv/ TPMa | Orthogroup | Mp§ Proteins in the OG | Total Proteins in the OG |
|---|---|---|---|---|---|---|---|---|
| 60.p3 | BAL70305.1 hypothetical protein | M. pulchricornis | 70,871 | 549 | 129 | |||
| 64.p1 | BAL70301.1 hypothetical protein | M. pulchricornis | 31,382 | 115 | 272 | OG0042547 | 1 | 2 |
| 32.p1 | BAL70307.1 hypothetical protein | M. pulchricornis | 30,408 | 124 | 245 | |||
| 338.p1 | BAL70305.1 hypothetical protein | M. pulchricornis | 28,057 | 174 | 161 | OG0042502 | 1 | 2 |
| 1043.p6 | uncharacterized protein LOC115116642 | Oncorhynchus nerka | 18,777 | 111 | 169 | |||
| 7952.p1 | No hit | 17,559 | 133 | 132 | ||||
| 38.p1 | uncharacterized protein LOC103570348 | M. demolitor | 15,273 | 94 | 162 | OG0042509 | 1 | 2 |
| 174.p2 | BAL70304.1 hypothetical protein | M. pulchricornis | 15,034 | 133 | 113 | OG0019101 | 5 | 6 |
| 29.p1 | BAL70308.1 hypothetical protein | M. pulchricornis | 13,879 | 66 | 210 | OG0042489 | 2 | 2 |
| 2896.p4 | No hit | 13,698 | 123 | 111 | OG0042486 | 1 | 2 | |
| 35.p1 | No hit | 13,271 | 131 | 101 | OG0021629 | 4 | 5 | |
| 4874.p12 | No hit | 12,545 | 103 | 121 | ||||
| 5.p1 | BAL70306.1 hypothetical protein | M. pulchricornis | 12,037 | 74 | 162 | OG0019101 | 5 | 6 |
| 42.p1 | No hit | 10,282 | 56 | 183 | OG0042514 | 1 | 2 | |
| 57.p1 | No hit | 10,256 | 105 | 97 | OG0042539 | 2 | 2 | |
| 39.p1 | BAL70290.1 hypothetical protein | M. pulchricornis | 9755 | 71 | 137 | OG0019101 | 5 | 6 |
| 121.p1 | BAL70308.1 hypothetical protein | M. pulchricornis | 9660 | 104 | 93 | OG0025719 | 4 | 4 |
| 6996.p3 | No hit | 8834 | 57 | 155 | ||||
| 6621.p3 | No hit | 8315 | 73 | 114 | ||||
| 52.p1 | BAL70302.1 hypothetical protein | M. pulchricornis | 8230 | 13 | 633 | OG0021663 | 4 | 5 |
| 281.p1 | No hit | 8078 | 17 | 475 | ||||
| 86.p1 | No hit | 8049 | 18 | 447 | OG0025738 | 4 | 4 | |
| 30.p1 | No hit | 7603 | 30 | 253 | OG0042491 | 1 | 2 | |
| 421.p1 | Low quality protein, fibrillin-1- partial | Anopheles darlingi | 7188 | 63 | 114 | OG0009646 | 17 | 20 |
| 186.p3 | No hit | 6991 | 129 | 54 | ||||
| 72.p1 | XP.023948444.1 uncharacterized protein LOC112053291 isoform X2 | Bicyclus anynana | 6968 | 36 | 193 | OG0015773 | 7 | 8 |
| 73.p1 | No hit | 6695 | 28 | 239 | OG0042561 | 1 | 2 | |
| 21.p1 | 5NUC-like | Apis cerana | 6656 | 4 | 1664 | OG0031454 | 2 | 3 |
| 11189.p3 | BAL70308.1 hypothetical protein | M. pulchricornis | 6606 | 42 | 157 | OG0025719 | 4 | 4 |
| 155.p1 | No hit | 5916 | 4 | 1479 | OG0009638 | 18 | 20 | |
| 2291.p1 | PREDICTED: location of vulva defective 1 X1 | M. demolitor | 5903 | 145 | 40 | OG0004499 | 3 | 36 |
| 4719.p2 | No hit | 5735 | 20 | 286 | ||||
| 285.p2 | No hit | 5626 | 29 | 194 | ||||
| 114.p1 | Lipase-related 2 | Cephus cinctus | 5395 | 31 | 174 | OG0010100 | 8 | 18 |
| 380.p3 | No hit | 5346 | 22 | 243 | ||||
| 3115.p10 | No hit | 5227 | 99 | 52 | ||||
| 49.p1 | BAL70302.1 hypothetical protein | M. pulchricornis | 5214 | 10 | 521 | |||
| 932.p4 | No hit | 5076 | 436 | 11 | ||||
| 132.p1 | No hit | 5042 | 47 | 107 | OG0015773 | 7 | 8 |
| CDS ° ID | Gel Bands * | Names and Functions of MpVLPs Proteins (E-Value Protein Blast) § | MpVLPs Mascot Score (Rank) | TV Mascot Score (Rank) | TPMv/TPMa | OG |
|---|---|---|---|---|---|---|
| 38 | TV8-9 P4-5 | Contig containing 2 CDS: - 38.p1, Uncharacterized protein LOC103577632 [Microplitis demolitor], 9 × 10−14. DUF4803 domain, 1.2 × 10−13. - 38.p2, Uncharacterized protein LOC106659714 [Trichogramma pretiosum], 5 × 10−12. DUF4803 domain, 7.2 × 10−12. | 44,936 (1) | 40,770 (1) | 15,273/95 1309/11 | OG0042509 OG0042510 |
| 35 | TV10 P5 | No hit | 24,427 (2) | 28,317 (2) | 13,271/131 | OG0021629 |
| 29 | TV14-17 P6-8 | Hypothetical protein [Meteorus pulchricornis] BAL70308.1, 3 × 10−164. | 13,954 (3) | 21,697 (4) | 13,879/67 | OG0042489 |
| 39 | TV13-17 P8-9 | Hypothetical protein [Meteorus pulchricornis] BAL70290.1., 7 × 10−127. | 11,019 (4) | 9659 (11) | 9755/71 | OG0019101 |
| 281 | TV6-7 P2-3 | No hit | 9946 (5) | 11,374 (10) | 8078/17 | na |
| 72 | TV7 P3 | Uncharacterized protein LOC106655966 [Trichogramma pretiosum], 6 × 10−11. DUF4803 domain, 5.9 × 10−10. | 9842 (6) | 7277 (19) | 6968/36 | OG0015773 |
| 52 | TV14-15 P7 | Hypothetical protein [Meteorus pulchricornis] BAL70302.1, 1 × 10−156. | 7157 (7) | 12,900 (8) | 8230/13 | OG0021663 |
| 1173 | TV21 P13 | Hypothetical protein [Meteorus pulchricornis] BAL70304.1, 9 × 10−42. | 6728 (8) | 5156 (27) | 4196/19 | OG0019101 |
| 110 | TV7 P3 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 2 × 10−15. Metalloproteases (“zincins”), catalytic domain, 9.2 × 10−9. | 6535 (9) | 3149 (41) | 763/5 | OG0000843 |
| 64 | TV22-25 P12-13 | Hypothetical protein [Meteorus pulchricornis] BAL70301.1, 3 × 10−72. | 6517 (10) | 15,886 (6) | 31,382/115 | OG0042547 |
| 42 | TV14-15 P7 | No hit | 5465 (11) | 6617 (21) | 10,282/57 | OG0042514 |
| 16 | TV12-13 P6 | A disintegrin and metalloproteinase with thrombospondin motifs 6-like [Microplitis demolitor], 1 × 10−14. A disintegrin and metalloproteinase with thrombospondin motifs (Domain Metalloproteases (“zincins”), catalytic domain, Reprolysin-like, 4.8 × 10−9. | 5007 (12) | 12,463 (9) | 3528/17 | OG0005678 |
| 6 | TV18-19 P9-10 | Contig containing 2 CDSs: - 6.p2, Uncharacterized protein LOC106128232 isoform X6 [Papilio xuthus], 6 × 10−26. Growth factor receptor domain, 4.5 × 10−8. - 6.p3, Fibrillin-1 [Dufourea novaeangliae], 7 × 10−18. Putative Growth factor receptor domain, 1.2 × 10−5. | 4948 (13) | 4063 (37) | 2924/23 398/28 | OG0009646 OG0009646 |
| 267 | TV13-14 P6-7 | Uncharacterized protein LOC105263636 [Fopius arisanus], 5 × 10−8. Hemolymph juvenile hormone binding domain (JHBP domain), 1.6 × 10−11. | 4058 (14) | 5138 (28) | 2950/28 | OG0025736 |
| 263 | TV13-14 C7 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1., 4 × 10−12. Metalloproteases (“zincins”), catalytic domain, 1.7 × 10−8. | 3397 (15) | 4678 (33) | 1340/6 | OG0001872 |
| 109 | TV13-14 P6-7 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 3 × 10−8. | 2190 (16) | 1630 (85) | 522/1 | OG0001872 |
| 73 | TV23-24 P13 | No hit | 2131 (17) | 7876 (16) | 6695/28 | OG0042561 |
| 285 | TV22 P12 | No hit | 2076 (18) | 1592 (73) | 5625/28 | na |
| 230 | TV5-6 P2 | Unknown. Lipocalins (Retinol binding protein-like) domain, 5 × 10−8. | 2071 (19) | 2654 (48) | 907/10 | OG0031457 |
| 668 | TV8-9 P4 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 4 × 10−11. | 1812 (20) | 2198 (57) | 676/6 | OG0000843 |
| 339 | TV6 P2 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, E = 0.0015. | 1726 (21) | 2811 (46) | 544/5 | OG0001872 |
| 126 | TV11 & TV22 P12 | A disintegrin and metalloproteinase with thrombospondin motifs 18-like [Chelonus insularis], 9 × 10−16. Metalloproteases (“zincins”), catalytic domain, 1.3 × 10−8. | 1566 (22) | 8290 (13) | 1684/1 | OG0000306 |
| 927 | TV8 P3-4 | Uncharacterized protein LOC118072154 [Chelonus insularis], 4 × 10−15. Metalloproteases (“zincins”) catalytic domain, 2 × 10−9. | 1531 (23) | 2074 (61) | 285/23 | OG0000306 |
| 5 | TV20-21 P11 | Hypothetical protein [Meteorus pulchricornis] BAL70306.1, 1 × 10−92. | 1511 (24) | 7780 (17) | 12,037/74 | OG0019101 |
| 8 | TV18-19 P10-11 | No hit | 1486 (25) | 5084 (29) | 1622/7 | OG0031468 |
| 1266 | TV6 P2 | No hit | 1476 (26) | 1598 (72) | 285/1 | OG0000306 |
| 1636 | TV7 P3 | No hit | 1463 (27) | 1578 (88) | 235/0.3 | OG0000843 |
| 875 | TV9 P4 | No hit | 1376 (28) | 1296 (78) | 1114/8 | OG0031455 |
| 208 | TV12-13 P6 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 2 × 10−14. Collagenase catalytic domain, 3.3 × 10−5. | 1368 (29) | 1575 (68) | 1051/5 | OG0001872 |
| 1290 | TV15 P7-8 | Uncharacterized protein LOC118072154 [Chelonus insularis], 4 × 10−19. Metalloproteases (“zincins”), catalytic domain, 6.1 × 10−12. | 1356 (30) | 2543 (57) | 188/3 | OG0005678 |
| 1198 | TV7 P3 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 4 × 10−13. | 1276 (31) | 635 (146) | 948/6 | OG0000306 |
| 2114 | P3-4 | No hit | 1259 (32) | 918 (108) | 480/3 | OG0000306 |
| 1039 | P2 | Fibrillin-2 like protein [Argiope bruennichi], 8 × 10−24. | 1086 (33) | 800 (125) | 386/2 | OG0009646 |
| 379.p2 | P3 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 6 × 10−17. | 1025 (34) | 1057 (94) | 429/7 | OG0000843 |
| 647 | TV7 P4 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 4 × 10−14. Metalloproteases (“zincins”), catalytic domain, 2 × 10−6. | 1023 (35) | 1282 (80) | 242/2 | OG0000843 |
| 897 | TV3-4 P1 | Venom protein 2 [Microctonus hyperodae], 7 × 10−122. DUF 4803 domain, 3.4 × 10−23. | 952 (36) | 21,507 (5) | 262/7 | OG0007440 |
| 293 | TV8 P3-4 | Hyaluronidase [Diachasma alloeum], 6 × 10−67. Hyaluronidase (Hyaluronidase domain), 5.2 × 10−60. | 934 (37) | 902 (110) | 363/1 | OG0005785 |
| 686 | P2 | No hit | 930 (38) | 956 (104) | 342/2 | OG0042554 |
| 496 | P6 | Venom metalloproteinase 2-like [Microplitis demolitor], 8 × 10−15. Collagenase (Catalytic Domain), 2.5 × 10−14. | 921 (39) | 1168 (89) | 279/4 | OG0009638 |
| 91 | TV6 P9 | Venom metalloproteinase 3-like [Pogonomyrmex barbatus], 3 × 10−17. Metallo-peptidase family M12 domain, 2.7 × 10−6. | 915 (40) | 1185 (110) | 182/1 | OG0000306 |
| 575 | P3 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, E = 0.001. | 910 (41) | 839 (153) | 952/6 | OG0000843 |
| 590 | P3 | Unknown. Putative Serpin domain, 2.8 × 10−6. | 896 (42) | 483 (223) | 641/4 | OG0042523 |
| 153 | TV13 P6 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 9 × 10−14. ADAM cysteine-rich domain, 2.4 × 10−5. | 818 (43) | 2230 (56) | 1637/11 | OG0001872 |
| 651 | TV10-11 P5 & P9-10 | Venom metalloproteinase 3-like [Chelonus insularis], 1 × 10−6. Collagenase Catalytic Domain, 1.2 × 10−6. | 804 (44) | 2022 (63) | 2059/4 | OG0009638 |
| 2103 | P4 | Uncharacterized protein LOC118072154 [Chelonus insularis], 8 × 10−12. | 801 (45) | 1171 (112) | 100/3 | OG0000843 |
| 4685 | TV10 P5 | No hit | 766 (46) | 881 (114) | 4416/30 | na |
| 375 | TV5 P2 | Venom metalloproteinase 2-like [Microplitis demolitor], 3 × 10−10. Metalloproteases (“zincins”), catalytic domain, 3.7 × 10−9. | 760 (47) | 1638 (71) | 161/4 | OG0000843 |
| 216.p1 | TV7 | Uncharacterized protein LOC118072154 [Chelonus insularis], 1 × 10−7. Collagenase catalytic domain, 1.1 × 10−5. | 710 (48) | 1227 (84) | 959/7 | OG0000306 |
| 609 | P2 | Unknown. Calycin beta-barrel core domain, 8.1 × 10−5. | 697 (49) | 601 (153) | 238/4 | OG0021629 |
| 429.p1 | TV10-11 & TV23 P5-P13 | Uncharacterized protein LOC118072154 [Chelonus insularis], 2 × 10−20. Metalloproteases (“zincins”), catalytic domain, 2.8 × 10−10. | 691 (50) | 3131 (42) | 439/11 | OG0000306 |
| 286 | TV14 & TV22 P12 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 2 × 10−9. Metalloproteases (“zincins”), catalytic domain, 1.6 × 10−8. | 690 (51) | 1198 (107) | 506/1 | OG0001872 |
| 244 | TV11-12 P8 | A disintegrin and metalloproteinase with thrombospondin motifs 15-like [Trichogramma pretiosum], 7 × 10−14. Metalloproteases (“zincins”), catalytic domain, 4.1 × 10−11. | 699 (52) | 3022 (44) | 501/4 | OG0009638 |
| 554 | P5 | Pancreatic lipase-related protein 2-like [Temnothorax curvispinosus], 3 × 10−68. Pancreatic lipase-like enzymes domain, 9.5 × 10−91. | 611 (53) | 1258 (82) | 602/9 | OG0010100 |
| 124 | TV20-21 P11 | Hypothetical predicted protein [Cloeon dipterum], 9 × 10−8. | 594 (54) | 1018 (97) | 1243/6 | OG0019104 |
| 185 | TV12-13 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 5 × 10−8. Metalloproteases (“zincins”), catalytic domain, 4.6 × 10−7. | 594 (55) | 2979 (50) | 789/3 | OG0001872 |
| 1150 | - | No hit. Lipocalins superfamily (Retinol binding protein-like), 2.1 × 10−6. | 584 (56) | 530 (164) | 113/1 | OG0021629 |
| 166 | - | No hit | 549 (57) | 438 (181) | 557/5 | OG0000306 |
| 378.p2 | - | A disintegrin and metalloproteinase with thrombospondin motifs 1-like [Microplitis demolitor], 3 × 10−8. ADAM cysteine-rich domain, 3.4 × 10−5. | 532 (58) | 204 (286) | 359/6 | OG0000306 |
| 943 | - | No hit | 530 (59) | 556 (157) | 101/2 | OG0000306 |
| 1411 | P6 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 9 × 10−15. | 523 (60) | 738 (136) | 238/0.3 | OG0001872 |
| CDS ° | Gel Bands * | Names and Functions of Venom Soluble Proteins (E-Value Protein Blast) § | TV Score (Rank) | MpVLPs Score (Rank) | TPMv/TPMa | Contig OG |
|---|---|---|---|---|---|---|
| 10764 | TV9-12 | Uncharacterized protein LOC103570348 [Microplitis demolitor], 6 × 10−22. DUF4803 domain, 5.2 × 10−16. | 23,572 (3) | 16 (489) | 0.9/0.3 | OG0015773 |
| 536 | TV12-13 | No hit. Lipocalins (Retinol binding protein-like) domain, 8.5 × 10−10. | 14,497 (7) | 278 (87) | 581/30 | OG0015781 |
| 341 | TV3-4 | Venom protein 2 [Microctonus hyperodae] 2 × 10−58. DUF4803 domain, 3.2 × 10−14. | 9650 (12) | 429 (69) | 196/4 | OG0025733 |
| 21 | TV4-5 | Protein 5NUC [Apis mellifera], 9 × 10−53. 5’-nucleotidase domain, 1.5 × 10−32. | 8248 (14) | 125 (136) | 6656/4 | OG0002704 |
| 114 | TV11-13 | Pancreatic lipase-related protein 2 [Bombus vancouverensis nearcticus], 5 × 10−52. Pancreatic lipase N-terminus domain, 2.6 × 10−48. | 7962 (15) | 106 (151) | 5394/31 | OG0010100 |
| 8250 | TV3-4 | Uncharacterized protein LOC105686340 isoform X1 [Athalia rosae], 3 × 10−59. DUF4803 domain, 3.7 × 10−11. | 7489 (18) | 60 (214) | 1/4 | OG0010765 |
| 7504 | TV4 | Uncharacterized protein LOC107042423 isoform X1 [Diachasma alloeum], 3 × 10−53. DUF4803 domain, 2.4 × 10−25. | 6704 (20) | 173 (118) | 0.4/5 | OG0010765 |
| 2792 | TV10 | Uncharacterized protein LOC103574503 [Microplitis demolitor], 1 × 10−28. DUF4803 domain, 4.9 × 10−8. | 6520 (22) | 73 (183) | 5/6 | OG0005090 |
| 136 | TV13 | No hit. Collagenase (Catalytic Domain), 1.7 × 10−9. | 6406 (23) | 193 (104) | 4945/25 | OG0000843 |
| 106 | TV5-6 | Predicted pancreatic triacylglycerol lipase [Wasmannia auropunctata], 9 × 10−35. Pancreatic lipase N-terminus domain, 4.2 × 10−41. | 6267 (24) | 63 (209) | 3310/21 | OG0031414 |
| 2331 | TV19-21 | No hit. Calycin beta-barrel core domain, 6.3 × 10−8. | 5752 (25) | 175 (114) | 52/8 | OG0015781 |
| 3422 | TV10-11 | Arginine kinase [Microplitis demolitor], E = 0.0. Phosphagen (guanidino) kinases such as arginine kinase and similar enzymes domain, E = 0.0. | 5653 (26) | 49 (238) | 1/764 | OG0001469 |
| 58 | TV11-12 | A disintegrin and metalloproteinase with thrombospondin motifs 6 [Galendromus occidentalis], 5 × 10−12. Metalloproteases (“zincins”), catalytic domain, 1.1 × 10−7. | 5079 (30) | 444 (66) | 1579/8 | OG0009638 |
| 8023 | TV4 | Venom protein [Ampulex compressa], 8 × 10−56. Two DUF4803 domains, 4.9 × 10−14 and 6.5 × 10−10. | 4972 (31) | 276 (88) | 3/2 | OG0010765 |
| 11420 | TV4-5 | Uncharacterized protein LOC115884266 [Sitophilus oryzae], 3 × 10−37. DUF4803 domain, 3.9 × 10−16. | 4758 (32) | 118 (141) | 298/6 | OG0010765 |
| 11339 | TV9-10, TV20 | γ-Glutamyltranspeptidase 1 isoform X1 [Microplitis demolitor] E = 0.0. γ-glutamyltranspeptidase domain, 2.4 × 10−156. | 4507 (34) | 101 (122) | 0.8 /3 | OG0000441 |
| 662.p2 | TV19 | Hypothetical protein [Meteorus pulchricornis] BAL70297.1, 5 × 10−139. | 4465 (35) | 181 (111) | 1321/49 | OG0042432 |
| 1164 | TV3 | Venom protein 2 [Microctonus hyperodae], 2 × 10−54. | 4306 (36) | not found | 11/6 | OG0008161 |
| 2455 | TV3 | Hypothetical protein TSAR.007430 [Trichomalopsis sarcophagae], 2 × 10−85. Eukaryotic CAP domain protein (cysteine-rich secretory proteins, antigen 5, and pathogenesis-related 1 proteins), 7.1 × 10−110. | 3922 (38) | not found | 32/1 | OG0000373 |
| 11685 | TV4 | Uncharacterized protein LOC105686340 isoform X2 [Athalia rosae], 1 × 10−17. DUF4803 domain, 5.7 × 10−25. | 3866 (39) | 118 (142) | 0.4/2 | OG0010765 |
| 275 | TV4 | Uncharacterized protein LOC108086051 [Drosophila kikkawai], 2 × 10−13. DUF4803 domain, 1.4 × 10−22. | 3455 (40) | 65 (195) | 3317 /19 | OG0025743 |
| 6649 | TV1 | Myosin heavy chain, muscle isoform X5 [Microplitis demolitor], E = 0.0. Class II myosin heavy chain 1, motor domain, E = 0.0. | 3032 (43) | not found | 0.5/153 | OG0000051 |
| 69 | TV13 | Glyceraldehyde-3-phosphate dehydrogenase [Athalia rosae], E = 0.0. Glyceraldehyde-3-phosphate dehydrogenase domain, E = 0.0. | 2911 (45) | not found | 205/1324 | OG0003321 |
| 229 | TV22 | No match | 2693 (47) | 109 (147) | 4413/13 | OG0001872 |
| 726 | TV3 | Uncharacterized protein LOC118068317 [Chelonus insularis], 2 × 10−52. DUF4803 domain, 9.3 × 10−36. | 2551 (49) | not found | 197/2 | OG0019097 |
| 53 | TV4 | Receptor-type tyrosine-protein phosphatase epsilon isoform X2 [Tribolium castaneum], 7 × 10−16. | 2514 (51) | 190 (107) | 4243/31 | OG0010765 |
| 155 | TV11 | Hypothetical protein [Meteorus pulchricornis] BAL70303.1, 7 × 10−6. Collagenase domain, 2.1 × 10−7. | 2458 (52) | not found | 5915 /4 | OG0009638 |
| 4840 | TV1 | Apolipophorins [Microplitis demolitor], 4 × 10−48. | 2451 (53) | 51 (232) | 0.4/189 | OG0013776 |
| 763 | TV12 | Predicted aldose reductase [Microplitis demolitor], E = 0.0. AKR1A family of aldo-keto reductase (AKR), 1.2 × 10−167. | 2365 (54) | not found | 112/257 | OG0000155 |
| 14605 | TV1 | Apolipophorins [Chelonus insularis], 2 × 10−48. | 2303 (55) | not found | 0.1/17 | OG0025456 |
| 1036 | TV16 | Putative 14-3-3 protein zeta isoform X1, partial [Cotesia chilonis], 4 × 10−172. 14-3-3 protein domain, 48 × 10−166. | 2188 (58) | 148 (126) | 309/970 | OG0000955 |
| 11280 | TV25 | Odorant-binding protein 3 [Meteorus pulchricornis], 1 × 10−92. Domain PBP/GOBP family, 1 × 10−22. | 2175 (59) | not found | 2/3700 | OG0000832 |
| 6971 | TV3 | Neprilysin-2-like isoform X2 [Belonocnema treatae], 6 × 10−67. Peptidase family M13 includes neprilysin, endothelin-converting enzyme I, 1.98 × 10−73. | 2098 (60) | not found | 1/1 | OG0017160 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gatti, J.-L.; Belghazi, M.; Legeai, F.; Ravallec, M.; Frayssinet, M.; Robin, S.; Aboubakar-Souna, D.; Srinivasan, R.; Tamò, M.; Poirié, M.; et al. Proteo-Trancriptomic Analyses Reveal a Large Expansion of Metalloprotease-Like Proteins in Atypical Venom Vesicles of the Wasp Meteorus pulchricornis (Braconidae). Toxins 2021, 13, 502. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13070502
Gatti J-L, Belghazi M, Legeai F, Ravallec M, Frayssinet M, Robin S, Aboubakar-Souna D, Srinivasan R, Tamò M, Poirié M, et al. Proteo-Trancriptomic Analyses Reveal a Large Expansion of Metalloprotease-Like Proteins in Atypical Venom Vesicles of the Wasp Meteorus pulchricornis (Braconidae). Toxins. 2021; 13(7):502. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13070502
Chicago/Turabian StyleGatti, Jean-Luc, Maya Belghazi, Fabrice Legeai, Marc Ravallec, Marie Frayssinet, Stéphanie Robin, Djibril Aboubakar-Souna, Ramasamy Srinivasan, Manuele Tamò, Marylène Poirié, and et al. 2021. "Proteo-Trancriptomic Analyses Reveal a Large Expansion of Metalloprotease-Like Proteins in Atypical Venom Vesicles of the Wasp Meteorus pulchricornis (Braconidae)" Toxins 13, no. 7: 502. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13070502