Acute Hepatopancreatic Necrosis Disease (AHPND): Virulence, Pathogenesis and Mitigation Strategies in Shrimp Aquaculture

 ,
, 
Abstract
:1. Introduction
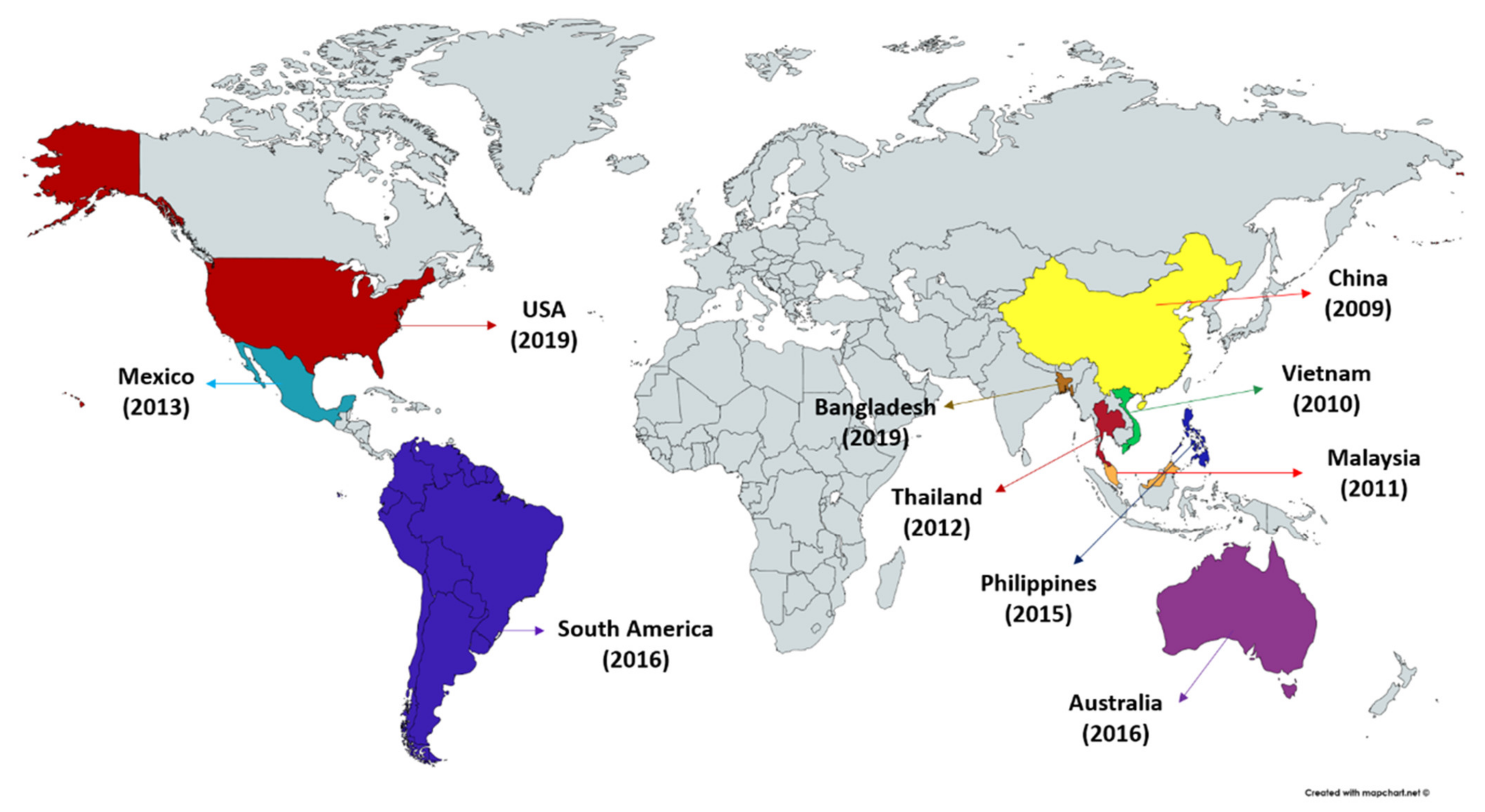
2. Acute Hepatopancreatic Necrosis Disease (AHPND)—An Overview
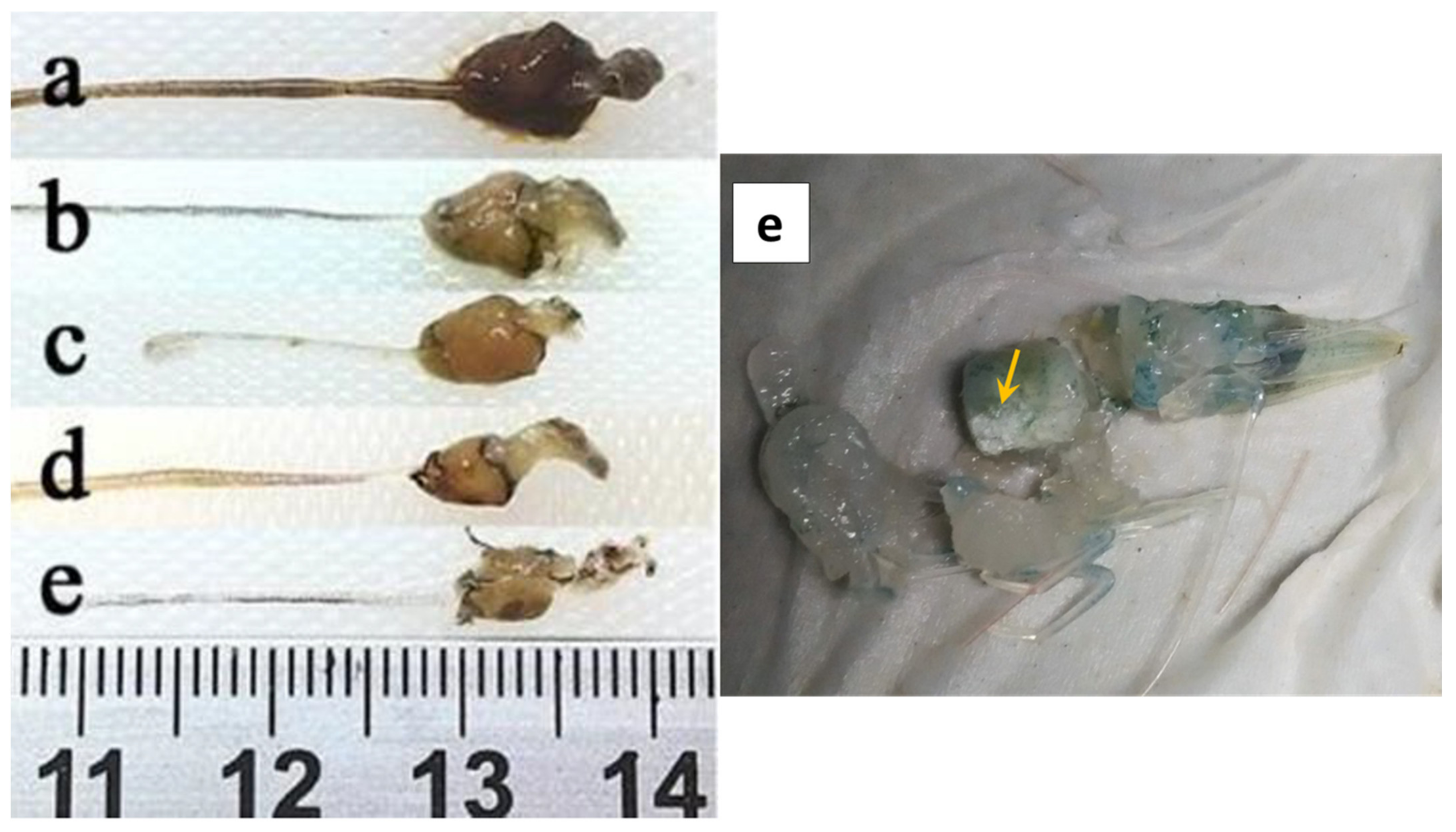
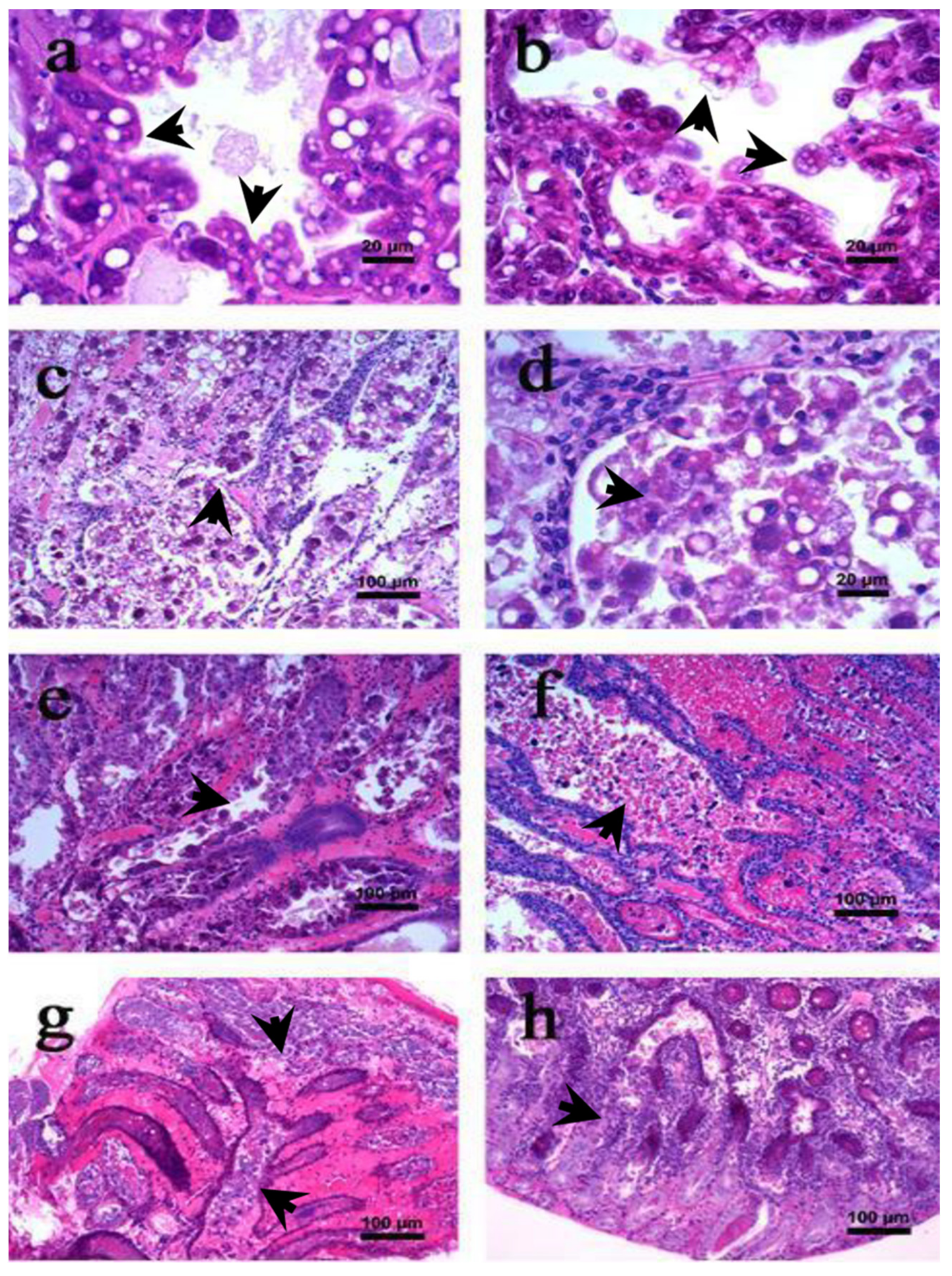
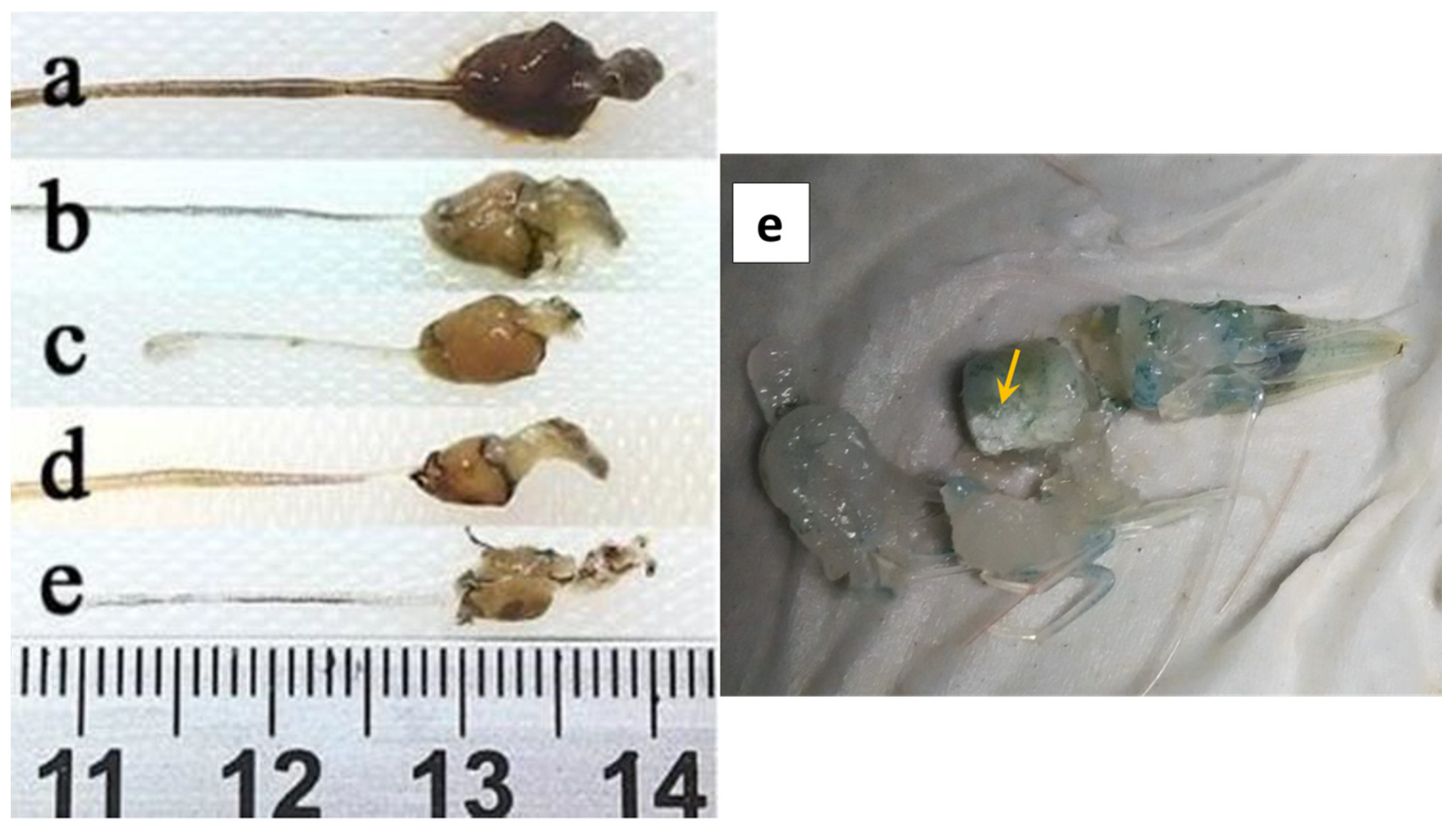
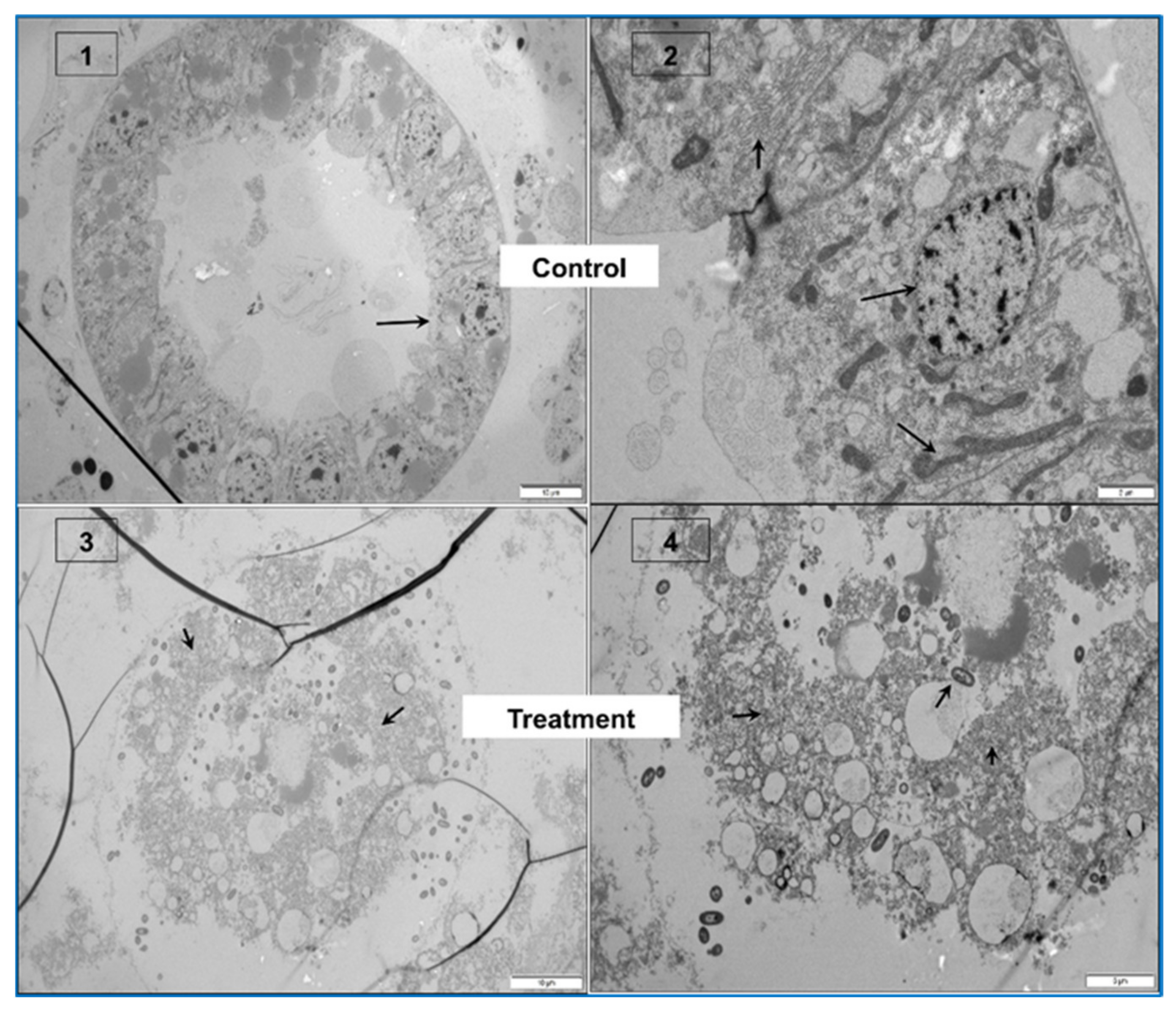
2.1. Gross Signs and Histopathology of AHPND
2.1.1. Initial Phase
2.1.2. Acute Phase
2.1.3. Terminal Phase
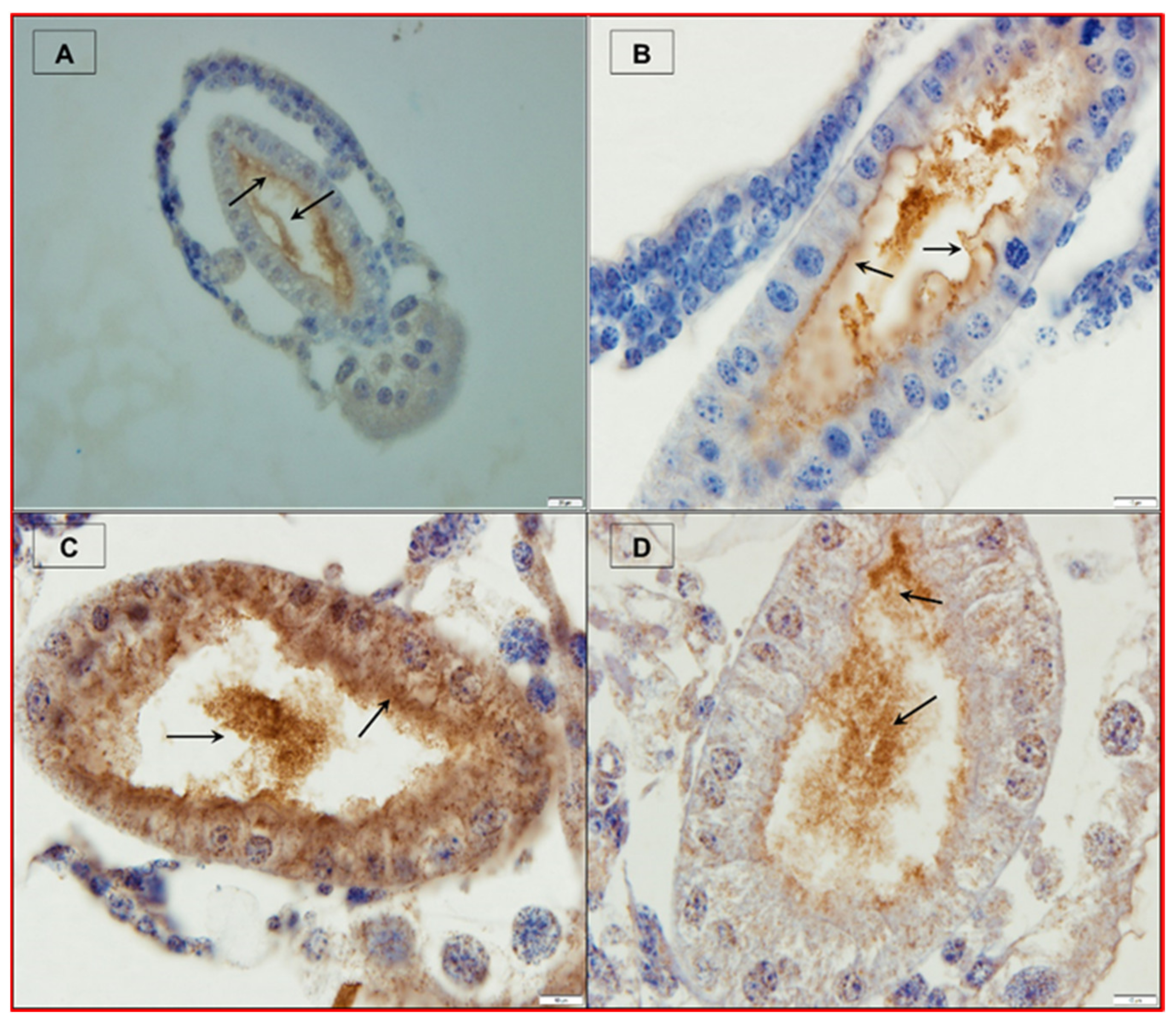
2.2. Causative Agent of AHPND
2.3. Vibrio parahaemolyticus as a Causative Agent of AHPND
3. Control and Management of Acute Hepatopancreatic Necrosis Disease (AHPND)—Current Status
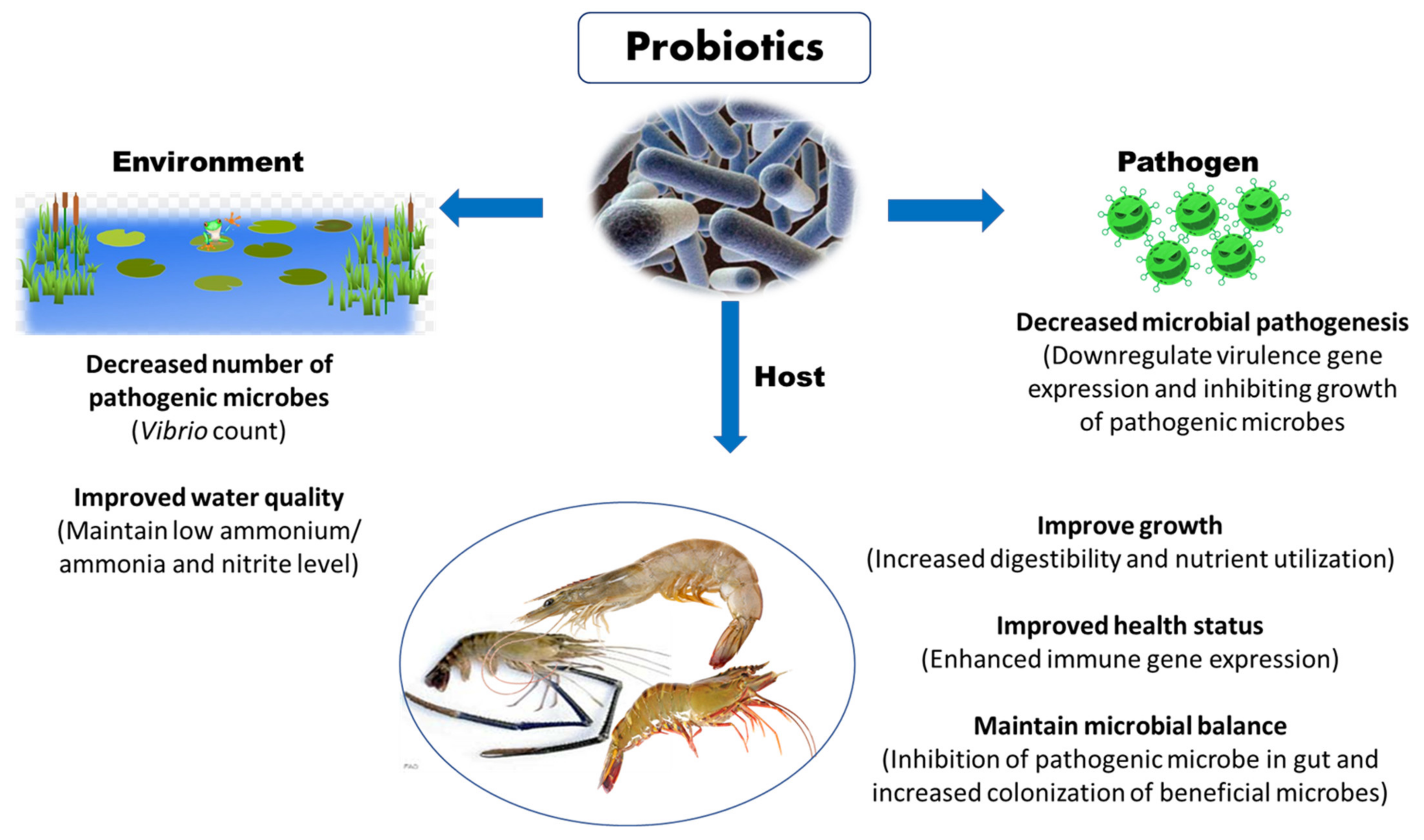
3.1. Probiotics
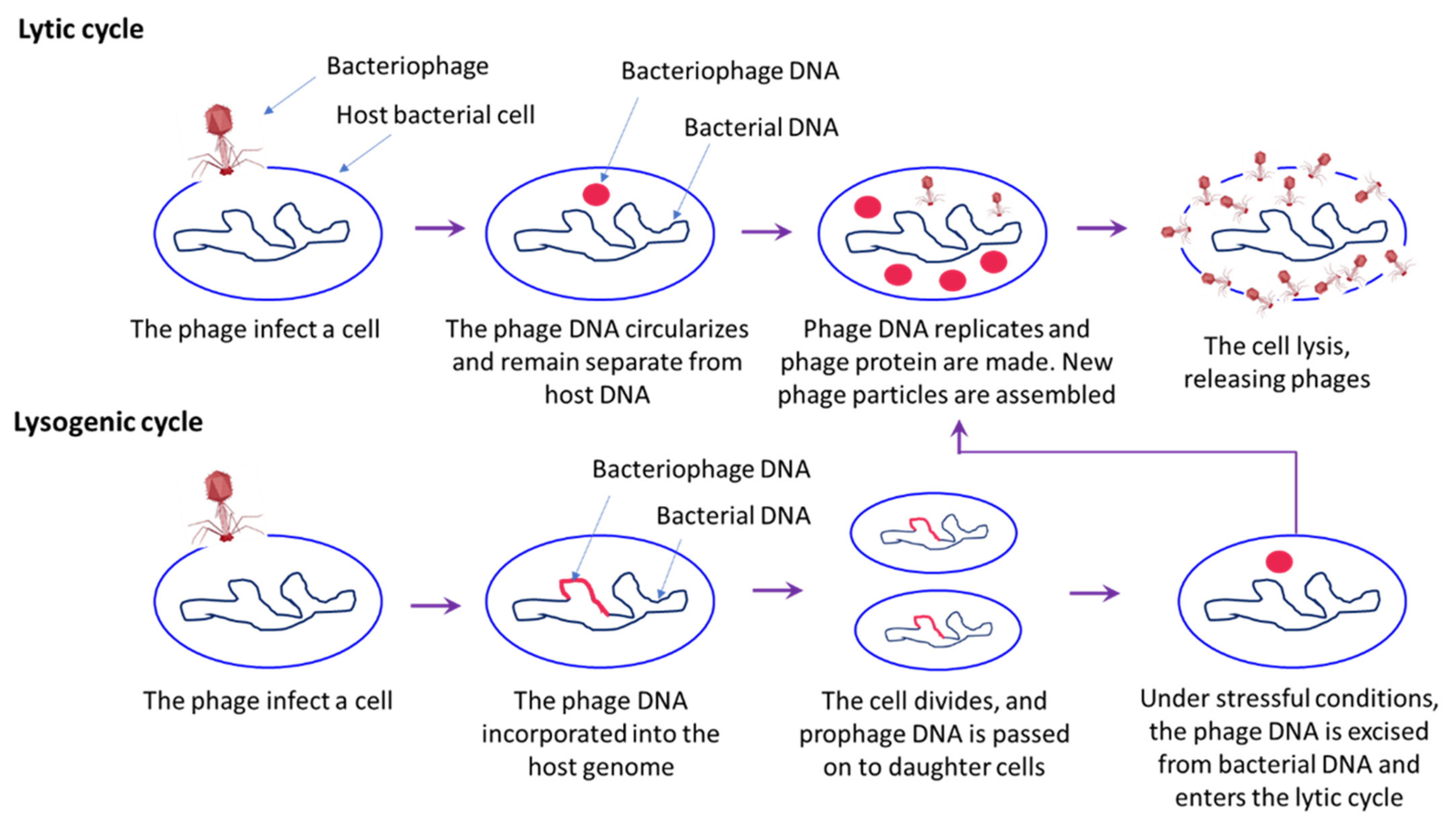
3.2. Phage Therapy
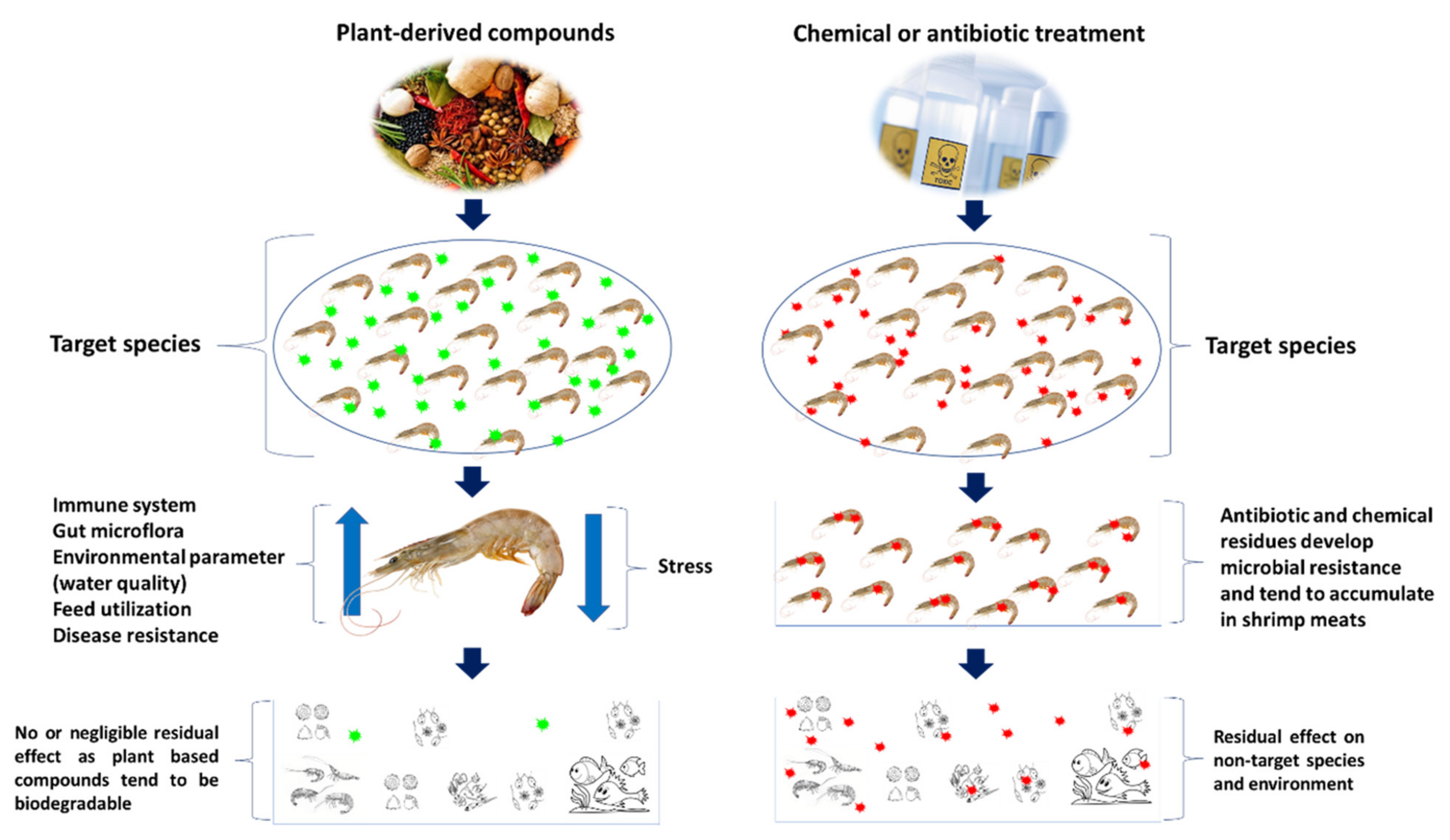
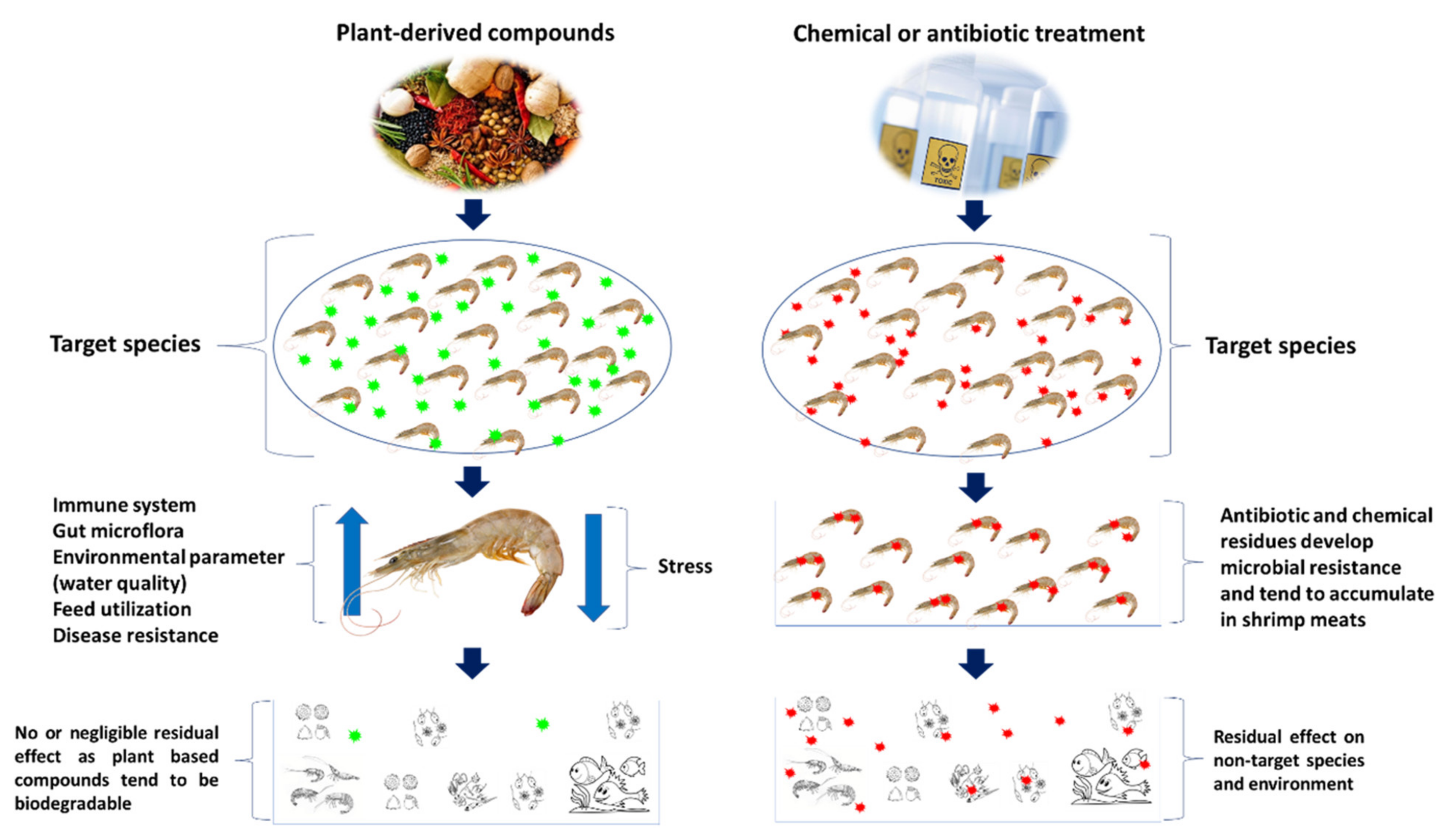
3.3. Plant-Derived and/or Natural Compounds
Immunostimulatory Properties of Plant-Based Compound
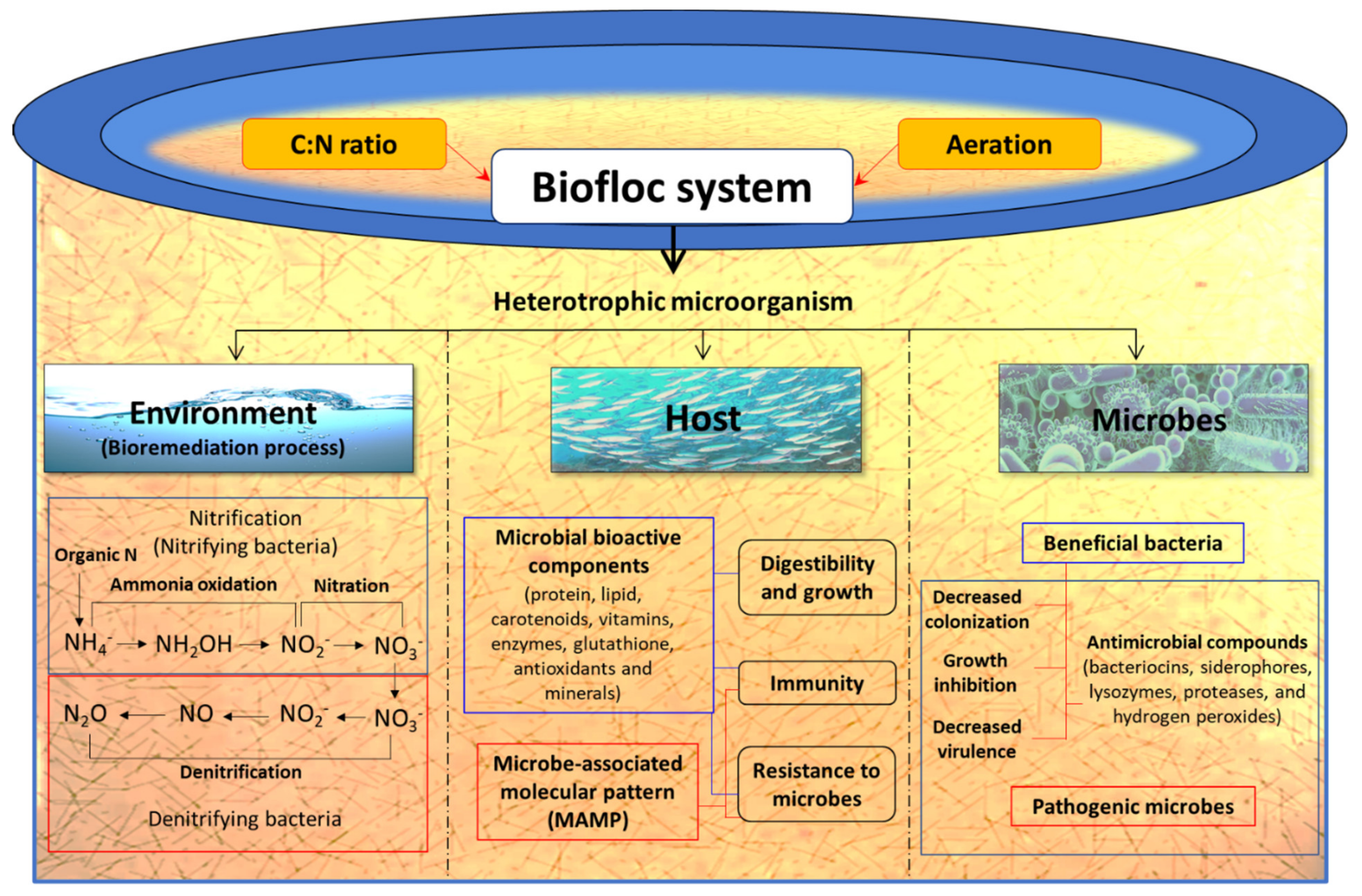
3.4. Environmental Manipulation
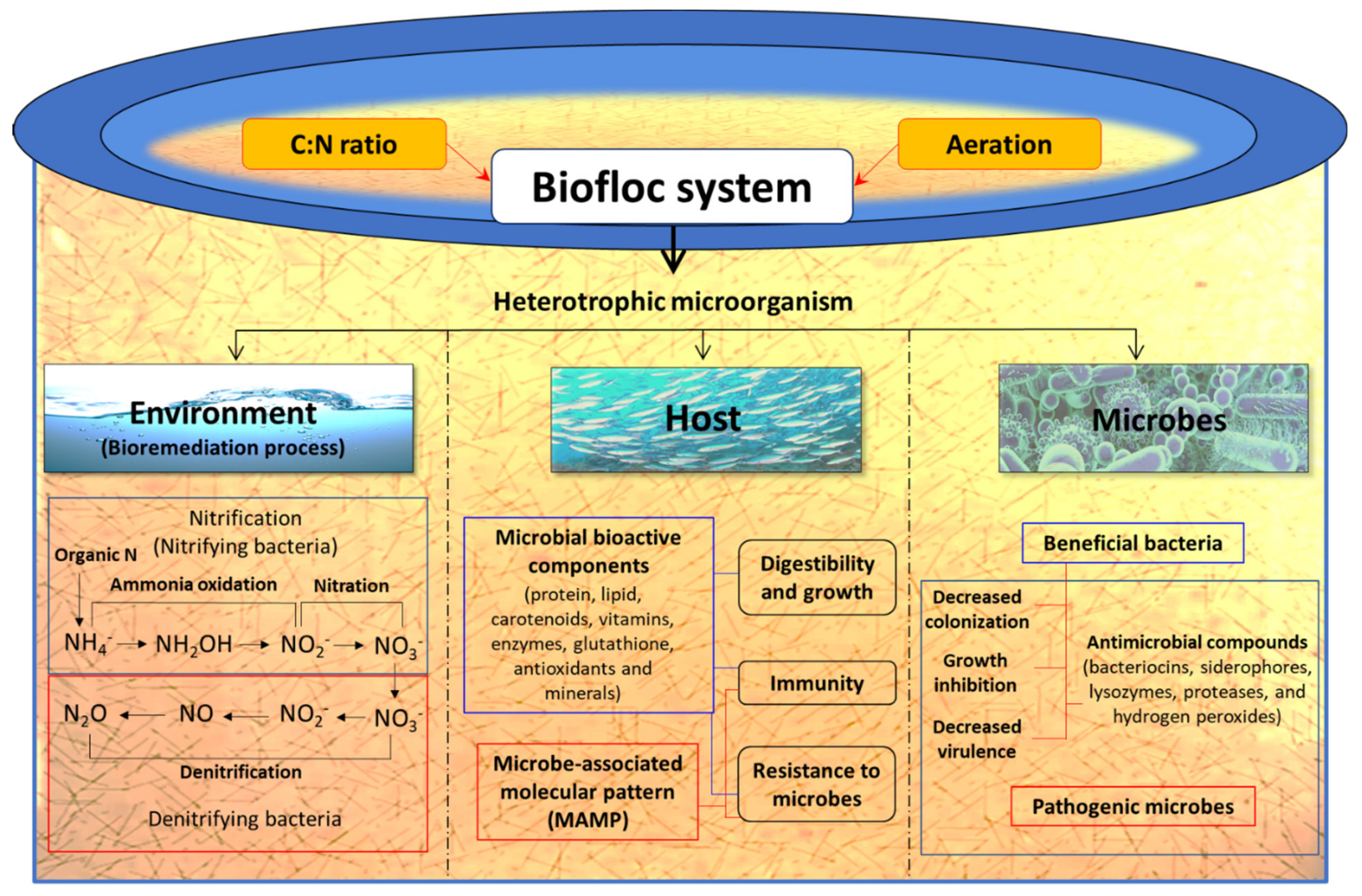
3.4.1. Biofloc Technology
3.4.2. Pond Management
4. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roy, S.; Kumar, V.; Manna, R.K.; Suresh, V.R. Sundarbans mangrove deltaic system—An overview of its biodiversity with special reference to fish diversity. J. Appl. Nat. Sci. 2016, 8, 1090–1099. [Google Scholar] [CrossRef] [Green Version]
- Soltani, M.; Ghosh, K.; Hoseinifar, S.H.; Kumar, V.; Lymbery, A.J.; Roy, S.; Ringø, E. Genus bacillus, promising probiotics in aquaculture: Aquatic animal origin, bio-active components, bioremediation and efficacy in fish and shellfish. Rev. Fish. Sci. Aquac. 2019, 27, 331–379. [Google Scholar] [CrossRef] [Green Version]
- FAO. State of Fisheries and Aquaculture in the World; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019; ISBN 9781424464968. [Google Scholar]
- Tacon, A.G.J. Trends in Global Aquaculture and Aquafeed Production: 2000–2017. Rev. Fish. Sci. Aquac. 2020, 28, 43–56. [Google Scholar] [CrossRef]
- FAO. State of Fisheries and Aquaculture in the World; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; ISBN 9789251305621. [Google Scholar]
- Sui, J.; Luan, S.; Yang, G.; Xia, Z.; Luo, K.; Tang, Q.; Lu, X.; Meng, X.; Kong, J. Genetic parameters and selection response for the harvest body weight of the giant freshwater prawn (Macrobrachium rosenbergii) in a breeding program in China. PLoS ONE 2019, 14, e0218379. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.L.; Valderrama, D.; Jory, D.E. GOAL 2019: Global Shrimp Production Review; GOAL: Portsmouth, NH, USA, 2019. [Google Scholar]
- Roy, S. Modulating Innate Immune Memory in Brine Shrimp (Artemia franciscana) and in Giant Freshwater Prawn (Macrobrachium rosenbergii). Ph.D. Thesis, University of Ghent, Ghent, Belgium, 2020. [Google Scholar]
- Verdegem, M.C.J. Nutrient discharge from aquaculture operations in function of system design and production environment. Rev. Aquac. 2013, 5, 158–171. [Google Scholar] [CrossRef]
- Lee, K.K.; Liu, P.C.; Chuang, W.H. Pathogenesis of gastroenteritis caused by Vibrio carchariae in cultured marine fish. Mar. Biotechnol. 2002, 4, 267–277. [Google Scholar] [CrossRef]
- Aguirre-Guzmán, G.; Ruíz, H.M.; Ascencio, F. A review of extracellular virulence product of Vibrio species important in diseases of cultivated shrimp. Aquac. Res. 2004, 35, 1395–1404. [Google Scholar] [CrossRef]
- Defoirdt, T.; Boon, N.; Sorgeloos, P.; Verstraete, W.; Bossier, P. Alternatives to antibiotics to control bacterial infections: Luminescent vibriosis in aquaculture as an example. Trends Biotechnol. 2007, 25, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Kesarcodi-Watson, A.; Kaspar, H.; Lategan, M.J.; Gibson, L. Two pathogens of greenshellTM mussel larvae, perna canaliculus: Vibrio splendidus and a V. coralliilyticus/neptunius-like isolate. J. Fish Dis. 2009, 32, 499–507. [Google Scholar] [CrossRef]
- Chatterjee, S.; Haldar, S. Vibrio Related Diseases in Aquaculture and Development of Rapid and Accurate Identification Methods. J. Mar. Sci. Res. Dev. 2012, s1, 1–7. [Google Scholar]
- Kumar, V.; Roy, S. Aquaculture Drugs: Sources, Active Ingredients, Pharmaceutic Preparations and Methods of Administration. J. Aquac. Res. Dev. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Tran, P.T.N.; Kumar, V.; Bossier, P. Do acute hepatopancreatic necrosis disease-causing PirABVP toxins aggravate vibriosis? Emerg. Microbes Infect. 2020, 9, 1919–1932. [Google Scholar] [CrossRef]
- Lee, C.-T.; Chen, I.-T.; Yang, Y.-T.; Ko, T.-P.; Huang, Y.-T.; Huang, J.-Y.; Huang, M.-F.; Lin, S.-J.; Chen, C.-Y.; Lin, S.-S.; et al. The opportunistic marine pathogen Vibrio parahaemolyticus becomes virulent by acquiring a plasmid that expresses a deadly toxin. Proc. Natl. Acad. Sci. USA 2015, 112, 10798–10803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, L.; Nunan, L.; Redman, R.M.; Mohney, L.L.; Pantoja, C.R.; Fitzsimmons, K.; Lightner, D.V. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Organ. 2013, 105, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Bi, D.; Wang, H.; Zou, P.; Xie, G.; Wan, X.; Yang, Q.; Zhu, Y.; Chen, M.; Guo, C.; et al. pirABvp-Bearing Vibrio parahaemolyticus and Vibrio campbellii pathogens isolated from the Same AHPND-affected pond possess highly similar pathogenic plasmids. Front. Microbiol. 2017, 8, 1859. [Google Scholar] [CrossRef]
- Choi, M.; Stevens, A.M.; Smith, S.A.; Taylor, D.P.; Kuhn, D.D. Strain and dose infectivity of Vibrio parahaemolyticus: The causative agent of early mortality syndrome in shrimp. Aquac. Res. 2017, 48, 3719–3727. [Google Scholar] [CrossRef]
- Kumar, R.; Ng, T.H.; Wang, H.C. Acute hepatopancreatic necrosis disease in penaeid shrimp. Rev. Aquac. 2020, 12, 1867–1880. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V. Acute Hepatopancreatic Necrosis Disease (AHPND) in Shrimp: Virulence, Pathogenesis and Mitigation Strategies. Ph.D. Thesis, University of Ghent, Ghent, Belgium, 2020. [Google Scholar]
- Leung, T.L.F.; Bates, A.E. More rapid and severe disease outbreaks for aquaculture at the tropics: Implications for food security. J. Appl. Ecol. 2013, 50, 215–222. [Google Scholar] [CrossRef]
- Kumar, V.; Baruah, K.; Nguyen, D.V.; Smagghe, G.; Vossen, E.; Bossier, P. Phloroglucinol mediated Hsp70 production in crustaceans: Protection against Vibrio parahaemolyticus in Artemia franciscana and Macrobrachium rosenbergii. Front. Immunol. 2018, 9, 1091. [Google Scholar] [CrossRef] [Green Version]
- Shinn, A.P.; Pratoomyot, J.; Griffiths, D.; Jiravanichpaisal, J.; Briggs, M. Asian shrimp production and the economic costs of disease. Asian Fish. Sci. J. 2018, 31S, 29–58. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014; Volume 2014, ISBN 9789251072257. [Google Scholar]
- Flegel, T.W. A future vision for disease control in shrimp aquaculture. J. World Aquac. Soc. 2019, 50, 249–266. [Google Scholar] [CrossRef]
- Hong, X.; Lu, L.; Xu, D. Progress in research on acute hepatopancreatic necrosis disease (AHPND). Aquac. Int. 2016, 24, 577–593. [Google Scholar] [CrossRef]
- Roy, S.; Kumar, V.; Bossier, P.; Norouzitallab, P.; Vanrompay, D. Phloroglucinol treatment induces transgenerational epigenetic inherited resistance against Vibrio infections and thermal stress in a brine shrimp (Artemia franciscana) model. Front. Immunol. 2019, 10, 2745. [Google Scholar] [CrossRef] [Green Version]
- Defoirdt, T.; Crab, R.; Wood, T.K.; Sorgeloos, P.; Verstraete, W.; Bossier, P. Quorum sensing-disrupting brominated furanones protect the gnotobiotic brine shrimp Artemia franciscana from pathogenic Vibrio harveyi, Vibrio campbellii, and Vibrio parahaemolyticus Isolates. Appl. Environ. Microbiol. 2006, 72, 6419–6423. [Google Scholar] [CrossRef] [Green Version]
- Crab, R.; Lambert, A.; Defoirdt, T.; Bossier, P.; Verstraete, W. The application of bioflocs technology to protect brine shrimp (Artemia franciscana) from pathogenic Vibrio harveyi. J. Appl. Microbiol. 2010, 109, 1643–1649. [Google Scholar] [CrossRef]
- Kumar, V.; Bels, L.D.; Couck, L.; Baruah, K.; Bossier, P.; Van den Broeck, W. PirABVP toxin binds to epithelial cells of the digestive tract and produce pathognomonic AHPND lesions in germ-free brine shrimp. Toxins 2019, 11, 717. [Google Scholar] [CrossRef] [Green Version]
- Han, J.E.; Tang, K.F.J.; Tran, L.H.; Lightner, D.V. Photorhabdus insect-related (Pir) toxin-like genes in a plasmid of Vibrio parahaemolyticus, the causative agent of acute hepatopancreatic necrosis disease (AHPND) of shrimp. Dis. Aquat. Organ. 2015, 113, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campa-Córdova, A.I.; León-Gallo, A.F.; Romero-Maldonado, A.; Ibarra-Serrano, A.C.; Rosales-Mendoza, S.; Hirono, I.; Angulo, C. Recombinant PirA-like toxin protects shrimp against challenge with Vibrio parahaemolyticus, the aetiological agent of acute hepatopancreatic necrosis disease. J. Fish Dis. 2017, 40, 1725–1729. [Google Scholar] [CrossRef]
- Soto-Rodriguez, S.A.; Gomez-Gil, B.; Lozano-Olvera, R.; Betancourt-Lozano, M.; Morales-Covarrubias, M.S. Field and experimental evidence of Vibrio parahaemolyticus as the causative agent of acute hepatopancreatic necrosis disease of cultured shrimp (Litopenaeus vannamei) in northwestern Mexico. Appl. Environ. Microbiol. 2015, 81, 1689–1699. [Google Scholar] [CrossRef] [Green Version]
- Soto-Rodriguez, S.A.; Gomez-Gil, B.; Lozano-Olvera, R.; Bolanmejia, C.; Aguilar-Rendon, K.G.; Enciso-Ibarra, J. Pathological, genomic and phenotypical characterization of Vibrio parahaemolyticus, causative agent of acute hepatopancreatic necrosis disease (AHPND) in Mexico. Asian Fish. Sci. 2018, 31, 102–111. [Google Scholar] [CrossRef]
- Sirikharin, R.; Taengchaiyaphum, S.; Sanguanrut, P.; Chi, T.D.; Mavichak, R.; Proespraiwong, P.; Nuangsaeng, B.; Thitamadee, S.; Flegel, T.W.; Sritunyalucksana, K. Characterization and PCR detection of binary, pir-like toxins from Vibrio parahaemolyticus isolates that cause acute hepatopancreatic necrosis disease (AHPND) in shrimp. PLoS ONE 2015, 10, e0126987. [Google Scholar] [CrossRef]
- Li, P.; Kinch, L.N.; Ray, A.; Dalia, A.B.; Cong, Q.; Nunan, L.M.; Camilli, A.; Grishin, N.V.; Salomon, D.; Orth, K. Acute hepatopancreatic necrosis disease-causing Vibrio parahaemolyticus strains maintain an antibacterial type VI secretion system with versatile effector repertoires. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Viet, D.; Baruah, K.; Bossier, P. Probing the mechanism of VP AHPND extracellular proteins toxicity purified from Vibrio parahaemolyticus AHPND strain in germ-free Artemia test system. Aquaculture 2019, 504, 414–419. [Google Scholar] [CrossRef]
- Dong, X.; Wang, H.; Zou, P.; Chen, J.; Liu, Z.; Wang, X.; Huang, J. Complete genome sequence of Vibrio campbellii strain 20130629003S01 isolated from shrimp with acute hepatopancreatic necrosis disease. Gut Pathog. 2017, 9, 1–5. [Google Scholar] [CrossRef]
- Liu, L.; Xiao, J.; Xia, X.; Pan, Y.; Yan, S. Draft Genome Sequence of Vibrio owensii Strain SH-14, Which Causes. Genome Announc. 2015, 3, 3354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo, L.; Bayot, B.; Arciniegas, S.; Bajaña, L.; Betancourt, I.; Panchana, F.; Muñoz, A.R. PirVP genes causing AHPND identified in a new Vibrio species (Vibrio punensis) within the commensal Orientalis clade. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Wechprasit, P.; Panphloi, M.; Thitamadee, S.; Sritunyalucksana, K.; Prachumwat, A. Complete Genome Sequence of Shewanella sp. strain TH2012, isolated from shrimp in a cultivation pond exhibiting early mortality syndrome. Microbiol. Resour. Announc. 2019, 8, e01703–18. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Roy, S.; Baruah, K.; Van Haver, D.; Impens, F.; Bossier, P. Environmental conditions steer phenotypic switching in acute hepatopancreatic necrosis disease-causing Vibrio parahaemolyticus, affecting PirAVP/PirBVP toxins production. Environ. Microbiol. 2020, 22, 4212–4230. [Google Scholar] [CrossRef]
- Tran, L.; Redman, R.M.; Lightner, D.V. EMS/AHPNS: Infectious Disease Caused by Bacteria. Glob. Aquac. Advocate 2013, 20, 19–20. [Google Scholar]
- Kumar, V.; Wille, M.; Lourenço, T.M.; Bossier, P. Biofloc-based enhanced survival of Litopenaeus vannamei Upon AHPND-causing Vibrio parahaemolyticus challenge is partially mediated by reduced expression of its virulence genes. Front. Microbiol. 2020, 11, 1270. [Google Scholar] [CrossRef]
- Nunan, L.; Lightner, D.; Pantoja, C.; Gomez-Jimenez, S. Detection of acute hepatopancreatic necrosis disease (AHPND) in Mexico. Dis. Aquat. Organ. 2014, 111, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Dhar, A.K.; Piamsomboon, P.; Caro, L.F.A.; Kanrar, S.; Adami, R.; Juan, Y.S. First report of acute hepatopancreatic necrosis disease (AHPND) occurring in the USA. Dis. Aquat. Organ. 2019, 132, 241–247. [Google Scholar] [CrossRef]
- Eshik, M.M.E.; Abedin, M.M.; Punom, N.J.; Begum, M.K.; Rahman, M.S. Molecular identification of AHPND positive Vibrio parahaemolyticus causing an outbreak in south-west shrimp farming regions of Bangladesh. J. Bangladesh Acad. Sci. 2017, 41, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Kondo, H.; Van, P.T.; Dang, L.T. Draft genome sequence of non- Vibrio parahaemolyticus acute diseased shrimp in Vietnam. Genome Announc. 2015, 3, 2014–2015. [Google Scholar] [CrossRef] [Green Version]
- Muthukrishnan, S.; Defoirdt, T.; Ina-Salwany, M.Y.; Yusoff, F.M.; Shariff, M.; Ismail, S.I.; Natrah, I. Vibrio parahaemolyticus and Vibrio harveyi causing Acute Hepatopancreatic Necrosis Disease (AHPND) in Penaeus vannamei (Boone, 1931) isolated from Malaysian shrimp ponds. Aquaculture 2019, 511, 734227. [Google Scholar] [CrossRef]
- Prachumwat, A.; Wechprasit, P.; Srisala, J.; Kriangsaksri, R.; Flegel, T.W.; Thitamadee, S. Shewanella khirikhana sp. nov.—A shrimp pathogen isolated from a cultivation pond exhibiting early mortality syndrome. Microb. Biotechnol. 2020, 13, 781–795. [Google Scholar] [CrossRef] [Green Version]
- Ffrench-Constant, R.H.; Waterfield, N.; Burland, V.; Perna, N.T.; Daborn, P.J.; Bowen, D.; Blattner, F.R. A genomic sample sequence of the entomopathogenic bacterium Photorhabdus luminescens W14: Potential implications for virulence. Appl. Environ. Microbiol. 2000, 66, 3310–3329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duchaud, E.; Rusniok, C.; Frangeul, L.; Buchrieser, C.; Givaudan, A.; Taourit, S.; Bocs, S.; Boursaux-Eude, C.; Chandler, M.; Charles, J.F.; et al. The genome sequence of the entomopathogenic bacterium Photorhabdus luminescens. Nat. Biotechnol. 2003, 21, 1307–1313. [Google Scholar] [CrossRef]
- Waterfield, N.; Kamita, S.G.; Hammock, B.D.; Ffrench-Constant, R. The Photorhabdus Pir toxins are similar to a developmentally regulated insect protein but show no juvenile hormone esterase activity. FEMS Microbiol. Lett. 2005, 245, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, M.B.; Farrar, R.R.; Novak, N.G.; Lawrence, S.D. Remarkable susceptibility of the diamondback moth (Plutella xylostella) to ingestion of Pir toxins from Photorhabdus luminescens. Entomol. Exp. Appl. 2006, 121, 31–37. [Google Scholar] [CrossRef]
- Ahantarig, A.; Chantawat, N.; Waterfield, N.R.; Ffrench-Constant, R.; Kittayapong, P. PirAB toxin from Photorhabdus asymbiotica as a larvicide against dengue vectors. Appl. Environ. Microbiol. 2009, 75, 4627–4629. [Google Scholar] [CrossRef] [Green Version]
- Kondo, H.; Tinwongger, S.; Proespraiwong, P.; Mavichak, R.; Unajak, S.; Nozaki, R.; Hirono, I. Draft genome sequences of six strains of Vibrio parahaemolyticus isolated from early mortality syndrome/acute hepatopancreatic necrosis disease shrimp in Thailand. Genome Announc. 2014, 2, e00221–14. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.J.; Hsu, K.C.; Wang, H.C. Structural insights into the cytotoxic mechanism of Vibrio parahaemolyticus PirAvp and PirBvp toxins. Mar. Drugs 2017, 15, 9. [Google Scholar] [CrossRef] [Green Version]
- Almanza-Martínez, N.; Francisco Martínez Díaz, S.; Flores-Ramírez, G.; Zuñiga-Navarrete, F.; Gómez, I.; Cardona-Félix, C.S. An α-amylase-like protein interacts with PirB toxin from Vibrio parahaemolyticus in digestive tract tissue of white shrimp Litopenaeus vannamei. Aquac. Res. 2020, 51, 3910–3914. [Google Scholar] [CrossRef]
- De Los Santos, M.V.; Vibanco-Pérez, N.; Soto-Rodriguez, S.; Pereyra, A.; Zenteno, E.; Cano-Sánchez, P. The B subunit of PirABvp toxin secreted from Vibrio parahaemolyticus causing AHPND is an amino sugar specific lectin. Pathogens 2020, 9, 1–15. [Google Scholar]
- Lai, H.C.; Ng, T.H.; Ando, M.; Te Lee, C.; Chen, I.T.; Chuang, J.C.; Mavichak, R.; Chang, S.H.; De Yeh, M.; Chiang, Y.A.; et al. Pathogenesis of acute hepatopancreatic necrosis disease (AHPND) in shrimp. Fish Shellfish. Immunol. 2015, 47, 1006–1014. [Google Scholar] [CrossRef]
- Gomez-gil, B.; Soto-rodríguez, S.; Lozano, R.; Betancourt-lozano, M. Draft genome sequence of Vibrio parahaemolyticus strain M0605, which causes severe mortalities of shrimps in Mexico. Genome Announc. 2014, 2, e00055–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osei-Adjei, G.; Huang, X.; Zhang, Y. The extracellular proteases produced by Vibrio parahaemolyticus. World J. Microbiol. Biotechnol. 2018, 34, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Acosta, J.A.; Martínez-Porchas, M.; Elizalde-Contreras, J.M.; Leyva, J.M.; Ruiz-May, E.; Gollas-Galván, T.; Martínez-Córdova, L.R.; Huerta-Ocampo, J.Á. Proteomic profiling of integral membrane proteins associated to pathogenicity in Vibrio parahaemolyticus strains. Microbiol. Immunol. 2018, 62, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Phiwsaiya, K.; Charoensapsri, W.; Taengphu, S.; Dong, H.T.; Sangsuriya, P.; Nguyen, G.T.; Pham, H.Q.; Amparyup, P.; Sritunyalucksana, K.; Taengchaiyaphum, S.; et al. A Natural Vibrio parahaemolyticus ΔpirAVp pirBVp+ Mutant Kills Shrimp but Produces neither PirVp Toxins nor Acute Hepatopancreatic Necrosis Disease Lesions. Appl. Environ. Microbiol. 2017, 83, e00680–17. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Liu, L.; Ke, Y.; Li, X.; Liu, Y.; Pan, Y.; Yan, S.; Wang, Y. Shrimp AHPND-causing plasmids encoding the PirAB toxins as mediated by pirAB-Tn903 are prevalent in various Vibrio species. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, C.V.; McCormick, B.A. Interactions of the Intestinal Epithelium with the Pathogen and the Indigenous Microbiota: A Three-Way Crosstalk. Interdiscip. Perspect. Infect. Dis. 2008, 2008, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almagro-Moreno, S.; Pruss, K.; Taylor, R.K. Intestinal Colonization Dynamics of Vibrio cholerae. PLoS Pathog. 2015, 11, e1004787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintana-Hayashi, M.; Padra, M.; Padra, J.; Benktander, J.; Lindén, S. Mucus-pathogen interactions in the gastrointestinal tract of farmed animals. Microorganisms 2018, 6, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dharmani, P.; Srivastava, V.; Kissoon-Singh, V.; Chadee, K. Role of intestinal mucins in innate host defense mechanisms against pathogens. J. Innate Immun. 2009, 1, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Rekecki, A.; Ringø, E.; Olsen, R.; Myklebust, R.; Dierckens, K.; Bergh, O.; Laureau, S.; Cornelissen, M.; Ducatelle, R.; Decostere, A.; et al. Luminal uptake of Vibrio (Listonella) anguillarum by shed enterocytes—A novel early defence strategy in larval fish. J. Fish Dis. 2013, 36, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Rekecki, A.; Gunasekara, R.A.Y.S.A.; Dierckens, K.; Laureau, S.; Boon, N.; Favoreel, H.; Cornelissen, M.; Sorgeloos, P.; Ducatelle, R.; Bossier, P.; et al. Bacterial host interaction of GFP-labelled Vibrio anguillarum HI-610 with gnotobiotic sea bass, Dicentrarchus labrax (L.), larvae. J. Fish Dis. 2012, 35, 265–273. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Maldonado-García, M.; Guluarte, C.; León-Gallo, A.; Rosales-Mendoza, S.; Ascencio, F.; Hirono, I.; Angulo, C. Evaluation of ToxA and Vibrio parahaemolyticus lysate on humoral immune response and immune-related genes in Pacific red snapper. Fish Shellfish Immunol. 2016, 56, 310–321. [Google Scholar] [CrossRef]
- Makino, K.; Oshima, K.; Kurokawa, K.; Yokoyama, K.; Uda, T.; Tagomori, K.; Iijima, Y.; Najima, M.; Nakano, M.; Yamashita, A.; et al. Genome sequence of Vibrio parahaemolyticus: A pathogenic mechanism distinct from that of V. cholerae. Lancet 2003, 361, 743–749. [Google Scholar] [CrossRef]
- Thompson, F.; Iida, T.; Swings, J. Biodiversity of Vibrios. Microbiol. Mol. Biol. Rev. 2004, 68, 403–431. [Google Scholar] [CrossRef] [Green Version]
- Karunasagar, I.; Karunasagar, I.; Raghunath, P. Ecology, Virulence, and Detection of Pathogenic and Pandemic Vibrio parahaemolyticus. Front. Microbiol. 2016, 7, 156. [Google Scholar] [CrossRef]
- Karunasagar, I.; Karunasagar, I. Ecology, virulence factors and global spread of vibrio parahaemolyticus. Asian Fish. Sci. 2018, 31, 15–28. [Google Scholar] [CrossRef]
- Beuchat, L.R. Environmental Factors Affecting Survival and Growth of Vibrio parahaemolyticus. A Review. J. Milk Food Technol. 1975, 38, 476–480. [Google Scholar] [CrossRef]
- Jayasree, L.; Janakiram, P.; Madhavi, R. Characterization of Vibrio spp. associated with diseased shrimp from culture ponds of Andhra Pradesh (India). J. World Aquac. Soc. 2006, 37, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Chonsin, K.; Matsuda, S.; Theethakaew, C.; Kodama, T.; Junjhon, J.; Suzuki, Y.; Suthienkul, O.; Iida, T. Genetic diversity of Vibrio parahaemolyticus strains isolated from farmed pacific white shrimp and ambient pond water affected by acute hepatopancreatic necrosis disease outbreak in Thailand. FEMS Microbiol. Lett. 2015, 363. [Google Scholar] [CrossRef] [Green Version]
- Vicente, A.; Taengphu, S.; Hung, A.L.; Mora, C.M.; Dong, H.T.; Senapin, S. Detection of Vibrio campbellii and V. parahaemolyticus carrying full-length pirABVp but only V. campbellii produces PirVp toxins. Aquaculture 2020, 519, 734708. [Google Scholar] [CrossRef]
- Tinwongger, S.; Nochiri, Y.; Thawonsuwan, J.; Nozaki, R.; Kondo, H.; Awasthi, S.P.; Hinenoya, A.; Yamasaki, S.; Hirono, I. Virulence of acute hepatopancreatic necrosis disease PirAB-like relies on secreted proteins not on gene copy number. J. Appl. Microbiol. 2016, 121, 1755–1765. [Google Scholar] [CrossRef]
- Okada, N.; Iida, T.; Park, K.S.; Goto, N.; Yasunaga, T.; Hiyoshi, H.; Matsuda, S.; Kodama, T.; Honda, T. Identification and characterization of a novel type III secretion system in trh-positive Vibrio parahaemolyticus strain TH3996 reveal genetic lineage and diversity of pathogenic machinery beyond the species level. Infect. Immun. 2009, 77, 904–913. [Google Scholar] [CrossRef] [Green Version]
- Santos, H.M.; Tsai, C.Y.; Maquiling, K.R.A.; Tayo, L.L.; Mariatulqabtiah, A.R.; Lee, C.W.; Chuang, K.P. Diagnosis and potential treatments for acute hepatopancreatic necrosis disease (AHPND): A review. Aquac. Int. 2020, 28, 169–185. [Google Scholar] [CrossRef]
- González-Gómez, J.P.; Soto-Rodriguez, S.; López-Cuevas, O.; Castro-del Campo, N.; Chaidez, C.; Gomez-Gil, B. Phylogenomic Analysis Supports Two Possible Origins for Latin American Strains of Vibrio parahaemolyticus Associated with Acute Hepatopancreatic Necrosis Disease (AHPND). Curr. Microbiol. 2020, 77, 3851–3860. [Google Scholar] [CrossRef]
- De Schryver, P.; Defoirdt, T.; Sorgeloos, P. Early mortality syndrome outbreaks: A microbial management issue in shrimp farming? PLoS Pathog. 2014, 10, 10–11. [Google Scholar] [CrossRef] [PubMed]
- Smith, P. Antimicrobial Resistance: The Use of Antimicrobials in the livestock sector. Rev. Sci. Tech. 2008, 27, 243–264. [Google Scholar] [CrossRef] [PubMed]
- Han, J.E.; Mohney, L.L.; Tang, K.F.J.; Pantoja, C.R.; Lightner, D.V. Plasmid mediated tetracycline resistance of Vibrio parahaemolyticus associated with acute hepatopancreatic necrosis disease (AHPND) in shrimps. Aquac. Rep. 2015, 2, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Bossier, P.; Norouzitallab, P.; Vanrompay, D. Trained immunity and perspectives for shrimp aquaculture. Rev. Aquac. 2020, 12, 2351–2370. [Google Scholar] [CrossRef]
- Hostin, B.; Wasielesky, W.; Decamp, O.; Bossier, P.; De Schryver, P. Managing input C/N ratio to reduce the risk of Acute Hepatopancreatic Necrosis Disease (AHPND) outbreaks in biofloc systems—A laboratory study. Aquaculture 2019, 508, 60–65. [Google Scholar] [CrossRef]
- Defoirdt, T. Virulence mechanisms of bacterial aquaculture pathogens and antivirulence therapy for aquaculture. Rev. Aquac. 2014, 6, 100–114. [Google Scholar] [CrossRef]
- Kumar, V.; Roy, S.; Meena, D.K.; Sarkar, U.K. Application of probiotics in shrimp aquaculture: Importance, mechanisms of action, and methods of administration. Rev. Fish. Sci. Aquac. 2016, 24, 342–368. [Google Scholar] [CrossRef]
- Aguilera-Rivera, D.; Prieto-Davó, A.; Escalante, K.; Chávez, C.; Cuzon, G.; Gaxiola, G. Probiotic effect of FLOC on Vibrios in the pacific white shrimp Litopenaeus vannamei. Aquaculture 2014, 424–425, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, C.; Tang, Y.; Sun, B.; Huang, J.; Song, X. Pseudoalteromonas probiotics as potential biocontrol agents improve the survival of Penaeus vannamei challenged with acute hepatopancreatic necrosis disease (AHPND)-causing Vibrio parahaemolyticus. Aquaculture 2018, 494, 30–36. [Google Scholar] [CrossRef]
- Pinoargote, G.; Flores, G.; Cooper, K.; Ravishankar, S. Effects on survival and bacterial community composition of the aquaculture water and gastrointestinal tract of shrimp (Litopenaeus vannamei) exposed to probiotic treatments after an induced infection of acute hepatopancreatic necrosis disease. Aquac. Res. 2018, 49, 3270–3288. [Google Scholar] [CrossRef] [Green Version]
- Kewcharoen, W.; Srisapoome, P. Probiotic effects of Bacillus spp. from Pacific white shrimp (Litopenaeus vannamei) on water quality and shrimp growth, immune responses, and resistance to Vibrio parahaemolyticus (AHPND strains). Fish Shellfish. Immunol. 2019, 94, 175–189. [Google Scholar] [CrossRef]
- Chomwong, S.; Charoensapsri, W.; Amparyup, P.; Tassanakajon, A. Two host gut-derived lactic acid bacteria activate the proPO system and increase resistance to an AHPND-causing strain of Vibrio parahaemolyticus in the shrimp Litopenaeus vannamei. Dev. Comp. Immunol. 2018, 89, 54–65. [Google Scholar] [CrossRef]
- Lim, S.Y.; Loo, K.W.; Wong, W.L. Synergistic antimicrobial effect of a seaweed-probiotic blend against acute hepatopancreatic necrosis disease (AHPND)-causing Vibrio parahaemolyticus. Probiotics Antimicrob. Proteins 2019, 12, 906–917. [Google Scholar] [CrossRef]
- Wang, Y.B. Effect of probiotics on growth performance and digestive enzyme activity of the shrimp Penaeus vannamei. Aquaculture 2007, 269, 259–264. [Google Scholar] [CrossRef]
- Loh, J. The role of probiotics and their mechanisms of action: An aquaculture perspective. World Aquac. 2017, 48, 19–23. [Google Scholar]
- Nguyen Thi Truc, L.; Trinh Ngoc, A.; Tran Thi Hong, T.; Nguyen Thanh, T.; Huynh Kim, H.; Pham Kim, L.; Huynh Truong, G.; Truong Quoc, P.; Nguyen Thi Ngoc, T. Selection of Lactic Acid Bacteria (LAB) Antagonizing Vibrio parahaemolyticus: The Pathogen of Acute Hepatopancreatic Necrosis Disease (AHPND) in Whiteleg Shrimp (Penaeus vannamei). Biology 2019, 8, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Li, J.; Zhu, G.; Zhao, K.; Jiang, W.; Li, H.; Wang, W.; Kumar, V.; Dong, S.; Zhu, W.; et al. Mechanism of the Potential therapeutic candidate Bacillus subtilis BSXE-1601 against shrimp pathogenic vibrios and multifunctional metabolites biosynthetic capability of the strain as predicted by genome analysis. Front. Microbiol. 2020, 11, 581802. [Google Scholar] [CrossRef] [PubMed]
- Twort, F.W. An Investigation on the Nature of Ultra-Microscopic Viruses. Lancet 1915, 186, 1241–1243. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.R.; March, J.B. Bacteriophages and biotechnology: Vaccines, gene therapy and antibacterials. Trends Biotechnol. 2006, 24, 212–218. [Google Scholar] [CrossRef]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage treatment of human infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, D.; Martínez, B.; Rodríguez, A.; García, P. Isolation and characterization of bacteriophages infecting Staphylococcus epidermidis. Curr. Microbiol. 2010, 61, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Jun, J.W.; Han, J.E.; Giri, S.S.; Tang, K.F.J.; Zhou, X.; Aranguren, L.F.; Kim, H.J.; Yun, S.; Chi, C.; Kim, S.G.; et al. Phage Application for the protection from acute hepatopancreatic necrosis disease (AHPND) in Penaeus vannamei. Indian J. Microbiol. 2018, 58, 114–117. [Google Scholar] [CrossRef]
- Angulo, C.; Loera-Muro, A.; Trujillo, E.; Luna-González, A. Control of AHPND by phages: A promising biotechnological approach. Rev. Aquac. 2019, 11, 989–1004. [Google Scholar] [CrossRef]
- Oakey, H.J.; Owens, L. A new bacteriophage, VHML, isolated from a toxin-producing strain of Vibrio harveyi in tropical Australia. J. Appl. Microbiol. 2000, 89, 702–709. [Google Scholar] [CrossRef]
- Shivu, M.M.; Rajeeva, B.C.; Girisha, S.K.; Karunasagar, I.; Krohne, G.; Karunasagar, I. Molecular characterization of Vibrio harveyi bacteriophages isolated from aquaculture environments along the coast of India. Environ. Microbiol. 2007, 9, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Crothers-Stomps, C.; Høj, L.; Bourne, D.G.; Hall, M.R.; Owens, L. Isolation of lytic bacteriophage against Vibrio harveyi. J. Appl. Microbiol. 2010, 108, 1744–1750. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Liang, Y.; Huang, S.; Zhang, J.; Wang, J. Isolation and Characterization of the Novel Phages vB_VpS_BA3 and vB_VpS_CA8 for Lysing Vibrio parahaemolyticus. Front. Microbiol. 2020, 11, 259. [Google Scholar] [CrossRef] [PubMed]
- Vinod, M.G.; Shivu, M.M.; Umesha, K.R.; Rajeeva, B.C.; Krohne, G.; Karunasagar, I.; Karunasagar, I. Isolation of Vibrio harveyi bacteriophage with a potential for biocontrol of luminous vibriosis in hatchery environments. Aquaculture 2006, 255, 117–124. [Google Scholar] [CrossRef]
- Karunasagar, I.; Shivu, M.M.; Girisha, S.K.; Krohne, G.; Karunasagar, I. Biocontrol of pathogens in shrimp hatcheries using bacteriophages. Aquaculture 2007, 268, 288–292. [Google Scholar] [CrossRef]
- Jun, J.W.; Han, J.E.; Tang, K.F.J.; Lightner, D.V.; Kim, J.; Seo, S.W.; Park, S.C. Potential application of bacteriophage pVp-1: Agent combating Vibrio parahaemolyticus strains associated with acute hepatopancreatic necrosis disease (AHPND) in shrimp. Aquaculture 2016, 457, 100–103. [Google Scholar] [CrossRef] [Green Version]
- Citarasu, T. Herbal biomedicines: A new opportunity for aquaculture industry. Aquac. Int. 2010, 18, 403–414. [Google Scholar] [CrossRef]
- Wunderlich, A.C.; Zica, É.D.O.P.; dos Santos Ayres, V.F.; Guimarães, A.C.; Takeara, R. Plant-Derived Compounds as an Alternative Treatment Against Parasites in Fish Farming: A Review; IntechOpen: London, UK, 2016; pp. 115–135. [Google Scholar]
- Ramudu, K.R.; Dash, G. A review on herbal drugs against harmful pathogens in aquaculture. Am. J. Drug Discov. Dev. 2013, 3, 209–219. [Google Scholar] [CrossRef]
- Sivasankar, P.; Santhiya, A.V.; Kanaga, V. A review on plants and herbal extracts against viral diseases in aquaculture. J. Med. Plants Stud. 2015, 3, 75–79. [Google Scholar]
- Jha, R.K.; Babikian, H.Y.; Babikian, H.Y.; Khoa, L.V.; Wisoyo, D.; Srisombat, S.; Jiaravanon, B. Efficacy of natural herbal formulation against acute hepatopancreatic necrosis disease (AHPND) causing Vibrio parahaemolyticus in Penaeus vannamei. Vet. Med. Open J. 2017, 2, 1–6. [Google Scholar] [CrossRef]
- Phuong, T.V.; Hai Yen, P.T.; Linh, N.Q. Antibacterial activity of extracts from dried and fresh herbal plant (Phyllanthus amarus) against pathogens causing acute hepatopancreatic necrosis disease (ahpnd) in white leg shrimp (Litopenaeus vannamei) at Thua Thien Hue province, Vietnam. Asploro J. Biomed. Clin. Case Rep. 2019, 2, 120–128. [Google Scholar]
- Boonsri, N.; Rudtanatip, T.; Withyachumnarnkul, B.; Wongprasert, K. Protein extract from red seaweed Gracilaria fisheri prevents acute hepatopancreatic necrosis disease (AHPND) infection in shrimp. J. Appl. Phycol. 2017, 29, 1597–1608. [Google Scholar] [CrossRef]
- Chang, Y.H.; Kuo, W.C.; Wang, H.C.; Chen, Y.M. Biocontrol of acute hepatopancreatic necrosis disease (AHPND) in shrimp using a microalgal-bacterial consortium. Aquaculture 2020, 521, 734990. [Google Scholar] [CrossRef]
- Tinh, T.H.; Elayaraja, S.; Mabrok, M.; Gallantiswara, P.C.D.; Vuddhakul, V.; Rodkhum, C. Antibacterial spectrum of synthetic herbal-based polyphenols against Vibrio parahaemolyticus isolated from diseased Pacific whiteleg shrimp (Penaeus vannamei) in Thailand. Aquaculture 2020, 533, 736070. [Google Scholar] [CrossRef]
- Kumar, V.; Bossier, P. Importance of plant-derived compounds and/or natural products in aquaculture. Aquafeed 2018, 10, 28–31. [Google Scholar]
- Kumar, V.; Bossier, P. Novel plant-based compounds could be useful in protecting shrimp species against AHPND Vibrio parahaemolyticus. J. Inl. Fish. Soc. India 2019, 51, 03–05. [Google Scholar]
- Dong, H.; Zheng, X.; Kumar, V.; Roy, S.; Duan, Y.; Gao, H.; Zhang, J. Dietary supplementation of teprenone potentiates thermal and hypoxia tolerance as well as cellular stress protection of Epinephelus coioides juveniles reared under multiple stressors. Aquaculture 2020, 514, 734413. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, S.; Pandey, P.K.; Raman, R.P.; Prasad, K.P.; Roy, S.; Kumar, A.; Kumar, K. Growth and Hemato-Immunological Response to Dietary i-Carrageenan in Labeo rohita (Hamilton, 1822) Juveniles. Isr. J. Aquac. 2014, 66, 20742. [Google Scholar]
- Dong, H.; Roy, S.; Zheng, X.; Kumar, V.; Das, B.K.; Duan, Y.; Sun, Y.; Zhang, J. Dietary teprenone enhances non-specific immunity, antioxidative response and resistance to hypoxia induced oxidative stress in Lateolabrax maculatus. Aquaculture 2020, 533, 736126. [Google Scholar] [CrossRef]
- Baruah, K.; Norouzitallab, P.; Linayati, L.; Sorgeloos, P.; Bossier, P. Reactive oxygen species generated by a heat shock protein (Hsp) inducing product contributes to Hsp70 production and Hsp70-mediated protective immunity in Artemia franciscana against pathogenic vibrios. Dev. Comp. Immunol. 2014, 46, 470–479. [Google Scholar] [CrossRef]
- Baruah, K.; Huy, T.T.; Norouzitallab, P.; Niu, Y.; Gupta, S.K.; De Schryver, P.; Bossier, P. Probing the protective mechanism of poly-ß-hydroxybutyrate against vibriosis by using gnotobiotic Artemia franciscana and Vibrio campbellii as host-pathogen model. Sci. Rep. 2015, 5, 1–8. [Google Scholar] [CrossRef]
- Baruah, K.; Norouzitallab, P.; Roberts, R.J.; Sorgeloos, P.; Bossier, P. A novel heat-shock protein inducer triggers heat shock protein 70 production and protects Artemia franciscana nauplii against abiotic stressors. Aquaculture 2012, 334, 152–158. [Google Scholar] [CrossRef]
- Niu, Y.; Norouzitallab, P.; Baruah, K.; Dong, S.; Bossier, P. A plant-based heat shock protein inducing compound modulates host-pathogen interactions between Artemia franciscana and Vibrio campbellii. Aquaculture 2014, 430, 120–127. [Google Scholar] [CrossRef]
- Baruah, K.; Phong, H.P.P.D.; Norouzitallab, P.; Defoirdt, T.; Bossier, P. The gnotobiotic brine shrimp (Artemia franciscana) model system reveals that the phenolic compound pyrogallol protects against infection through its prooxidant activity. Free Radic. Biol. Med. 2015, 89, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Baruah, K.; Norouzitallab, P.; Phong, H.P.P.D.; Smagghe, G.; Bossier, P. Enhanced resistance against Vibrio harveyi infection by carvacrol and its association with the induction of heat shock protein 72 in gnotobiotic Artemia franciscana. Cell Stress Chaperones 2017, 22, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindquist, S. The heat-shock response. Annu. Rev. Biochem. 1986, 55, 1151–1191. [Google Scholar] [CrossRef]
- Richter, K.; Haslbeck, M.; Buchner, J. The Heat Shock Response: Life on the Verge of Death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.J.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, T.; Akpinar-Bayizit, A.; Yilmaz-Ersan, L.; Delikanli, B. Phenolics in Human Health. Int. J. Chem. Eng. Appl. 2014, 5, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Çelik, T.A.; Aslantürk, Ö.S. Cytotoxic and genotoxic effects of Lavandula stoechas aqueous extracts. Biologia 2007, 62, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Hamidi, R.M.; Jovanova, B.; Kadifkova Panovska, T. Toxicological evaluation of the plant products using brine shrimp (Artemia salina L.) model. Maced. Pharm. Bull. 2014, 60, 9–18. [Google Scholar] [CrossRef]
- Sultana, T.; Haque, M.; Salam, M.; Alam, M. Effect of aeration on growth and production of fish in intensive aquaculture system in earthen ponds. J. Bangladesh Agric. Univ. 2017, 15, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Welker, T.L.; Overturf, K.; Abernathy, J. Effect of aeration and oxygenation on growth and survival of rainbow trout in a commercial serial-pass, flow-through raceway system. Aquac. Rep. 2019, 14, 100194. [Google Scholar] [CrossRef]
- Guo, M.; Gross, C.A. Stress induced remodeling of the bacterial proteome. Curr. Biol. 2014, 24, R424–R434. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, A.P.; Sousa, J.C. Effect of shear stress on growth, adhesion and biofilm formation of Pseudomonas aeruginosa with antibiotic-induced morphological changes. Int. J. Antimicrob. Agents 2007, 30, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Kent, A.G.; Garcia, C.A.; Martiny, A.C.; Biology, E. Increased biofilm formation due to high temperature adaptation in marine Roseobacter. Nat. Microbiol. 2018, 3, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Yang, C.; Qiu, H.; Wang, H.; Yang, R.; Falush, D. The landscape of coadaptation in Vibrio parahaemolyticus. bioRxiv 2019, 9, 373936. [Google Scholar] [CrossRef]
- Franken, J.; Brandt, B.A.; Tai, S.L.; Bauer, F.F. Biosynthesis of Levan, a Bacterial Extracellular Polysaccharide, in the Yeast Saccharomyces cerevisiae. PLoS ONE 2013, 8, e77499. [Google Scholar] [CrossRef]
- Kumar, V.; Baruah, K.; Bossier, P. Bamboo powder protects gnotobiotically-grown brine shrimp against AHPND-causing Vibrio parahaemolyticus strains by cessation of PirABVP toxin secretion. Aquaculture 2021, 5539, 736624. [Google Scholar] [CrossRef]
- Ekasari, J.; Hanif, M.; Surawidjaja, E.H.; Nuryati, S.; De Schryver, P.; Bossier, P. Immune response and disease resistance of shrimp fed biofloc grown on different carbon sources. Fish Shellfish Immunol. 2014, 41, 332–339. [Google Scholar] [CrossRef]
- Avnimelech, Y. Carbon/nitrogen ratio as a control element in aquaculture systems. Aquaculture 1999, 176, 227–235. [Google Scholar] [CrossRef]
- Crab, R.; Defoirdt, T.; Bossier, P.; Verstraete, W. Biofloc technology in aquaculture: Beneficial effects and future challenges. Aquaculture 2012, 356–357, 351–356. [Google Scholar] [CrossRef]
- Kumar, V.S.; Pandey, P.K.; Anand, T.; Bhuvaneswari, G.R.; Dhinakaran, A.; Kumar, S. Biofloc improves water, effluent quality and growth parameters of Penaeus vannamei in an intensive culture system. J. Environ. Manag. 2018, 215, 206–215. [Google Scholar] [CrossRef]
- Bossier, P.; Ekasari, J. Biofloc technology application in aquaculture to support sustainable development goals. Microb. Biotechnol. 2017, 10, 1012–1016. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Forster, I.; Conquest, L.; Dominy, W. Enhanced growth effects on shrimp (Litopenaeus vannamei) from inclusion of whole shrimp floc or floc fractions to a formulated diet. Aquac. Nutr. 2008, 14, 533–543. [Google Scholar] [CrossRef]
- Burford, M.A.; Thompson, P.J.; Bauman, H.; Pearson, D.C. Microbial communities affect water quality, shrimp performance at Belize Aquaculture. Glob. Aquac. Advocate 2003, 6, 64–65. [Google Scholar]
- Wasielesky, W.; Atwood, H.; Stokes, A.; Browdy, C.L. Effect of natural production in a zero exchange suspended microbial floc based super-intensive culture system for white shrimp Litopenaeus vannamei. Aquaculture 2006, 258, 396–403. [Google Scholar] [CrossRef]
- Trichet, V.V. Nutrition and immunity: An update. Aquac. Res. 2010, 41, 356–372. [Google Scholar] [CrossRef]
- Cardona, E.; Saulnier, D.; Lorgeoux, B.; Chim, L.; Gueguen, Y. Rearing effect of biofloc on antioxidant and antimicrobial transcriptional response in Litopenaeus stylirostris shrimp facing an experimental sub-lethal hydrogen peroxide stress. Fish Shellfish Immunol. 2015, 45, 933–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, D.D.; Boardman, G.D.; Lawrence, A.L.; Marsh, L.; Flick, G.J. Microbial floc meal as a replacement ingredient for fish meal and soybean protein in shrimp feed. Aquaculture 2009, 296, 51–57. [Google Scholar] [CrossRef]
- Anand, P.S.S.; Kumar, S.; Kohli, M.P.S.; Sundaray, J.K.; Sinha, A.; Pailan, G.H.; Roy, S. Dietary biofloc supplementation in black tiger shrimp, Penaeus monodon: Effects on immunity, antioxidant and metabolic enzyme activities. Aquac. Res. 2017, 48, 4512–4523. [Google Scholar] [CrossRef]
- Cohen, J.M.; Samocha, T.M.; Fox, J.M.; Gandy, R.L.; Lawrence, A.L. Characterization of water quality factors during intensive raceway production of juvenile Litopenaeus vannamei using limited discharge and biosecure management tools. Aquac. Eng. 2005, 32, 425–442. [Google Scholar] [CrossRef]
- Azim, M.E.; Little, D.C. The biofloc technology (BFT) in indoor tanks: Water quality, biofloc composition, and growth and welfare of Nile tilapia (Oreochromis niloticus). Aquaculture 2008, 283, 29–35. [Google Scholar] [CrossRef]
- Lee, C.; Kim, S.; Lim, S.; Lee, K. Supplemental effects of biofloc powder on growth performance, innate immunity, and disease resistance of Pacific white shrimp Litopenaeus vannamei. Fish. Aquat. Sci. 2017, 20, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Shinn, A.P.; Jiravanichpaisal, J.; Grifffiths, D.; Pokharatsiri, A.; Burana, P.; Sumon, T.; Tongmee, C.; Decamp, O.; Galli, L. Effect of Biofloc on the Survival of Whiteleg Shrimp, Penaeus vannamei Boone 1931, When challenged with a pathogenic strain of Vibrio parahaemolyticus, the causative agent of acute hepatopancreatic necrosis disease. Asian Fish. Sci. 2018, 31, 210–225. [Google Scholar]
- Sajali, U.S.B.A.; Atkinson, N.L.; Desbois, A.P.; Little, D.C.; Murray, F.J.; Shinn, A.P. Prophylactic properties of bio floc- or Nile tilapia-conditioned water against Vibrio parahaemolyticus infection of whiteleg shrimp (Penaeus vannamei). Aquaculture 2019, 498, 496–502. [Google Scholar] [CrossRef]
- Putth, S.; Polchana, J. Current status and impact of early mortality syndrome (EMS)/acute hepatopancreatic necrosis disease (AHPND) and hepatopancreatic microsporidiosis (HPM) outbreaks on Thailand s shrimp farming. Southeast Asian Fish. Dev. 2016, 79–87. [Google Scholar]
- Tran, L.H.; Ftizsimmons, K.M.; Lightner, D.V. Tilapia could enhance water conditions, help control EMS in shrimp ponds. Glob. Aquac. Advocate 2014, 40, 11–12. [Google Scholar]
- Boonyawiwat, V.; Patanasatienkul, T.; Kasornchandra, J.; Poolkhet, C.; Yaemkasem, S.; Hammell, L.; Davidson, J. Impact of farm management on expression of early mortality syndrome/acute hepatopancreatic necrosis disease (EMS/AHPND) on penaeid shrimp farms in Thailand. J. Fish Dis. 2017, 40, 649–659. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Host Range | Geographical Distribution | References |
|---|---|---|---|
| Vibrio parahaemolyticus | P. monodon, L. vannamei | Worldwide | [17,18,33,45,46,47,48,49] |
| V. parahaemolyticus | Artemia franciscana | Laboratory condition | [16,24,29,32,39,44] |
| V. parahaemolyticus | Macrobrachium rosenbergii | Laboratory condition | [24,44] |
| V. punensis | L. vannamei | South America | [42] |
| V. harveyi | L. vannamei | China, Malaysia, Vietnam | [42,50,51] |
| V. owensii | L. vannamei | China | [41] |
| V. campbelli | L. vannamei | China | [40] |
| Shewanella sp. | L. vannamei | Thailand | [43,52] |
| Class | Chemical Structure | Sub-Class | Example | Role in Aquatic Species |
|---|---|---|---|---|
| Phenolics |  | Quinones, lavonoids, flavones, tannins, flavonols | Allium spp. (A. cepa, A. sativum, A. tuberosum), Cynodon dactylon, Viscum album, etc. | Immunostimulant, antioxidant, antimicrobial, growth promotor, anti-helminthic, antiviral |
| Alkaloids |  | Camellia sinensis, Nicotiana tabacum, Aconitum napellus, Atropa belladonna, Conium maculatum, etc. | Immunostimulant, antioxidant, antimicrobial, growth promotor, anti-helminthic, antiviral | |
| Terpenoids and essential oils |  | Pistacia terebinthus, Lavandula angustifolia, Mentha piperita, Melaleuca alternifolia, etc. | Immunostimulant, antimicrobial, antioxidant, anti-helminthic, growth promotor | |
| Lectins and polypeptides |  | Glycine max, Arachis hypogaea, Triticum aestivum, Cocos nucifera, etc. | Antioxidant, antiviral, immunostimulant | |
| Polyacetylenes |   | Anethum graveolens, Carum, carvi, Daucus carota, etc. | Immunostimulant, antimicrobial, antioxidant |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, V.; Roy, S.; Behera, B.K.; Bossier, P.; Das, B.K. Acute Hepatopancreatic Necrosis Disease (AHPND): Virulence, Pathogenesis and Mitigation Strategies in Shrimp Aquaculture. Toxins 2021, 13, 524. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080524
Kumar V, Roy S, Behera BK, Bossier P, Das BK. Acute Hepatopancreatic Necrosis Disease (AHPND): Virulence, Pathogenesis and Mitigation Strategies in Shrimp Aquaculture. Toxins. 2021; 13(8):524. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080524
Chicago/Turabian StyleKumar, Vikash, Suvra Roy, Bijay Kumar Behera, Peter Bossier, and Basanta Kumar Das. 2021. "Acute Hepatopancreatic Necrosis Disease (AHPND): Virulence, Pathogenesis and Mitigation Strategies in Shrimp Aquaculture" Toxins 13, no. 8: 524. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080524