Changes of DNA Damage Effect of T-2 or Deoxynivalenol Toxins during Three Weeks Exposure in Common Carp (Cyprinus carpio L.) Revealed by LORD-Q PCR

 , and
, and
Abstract
:1. Introduction
2. Results
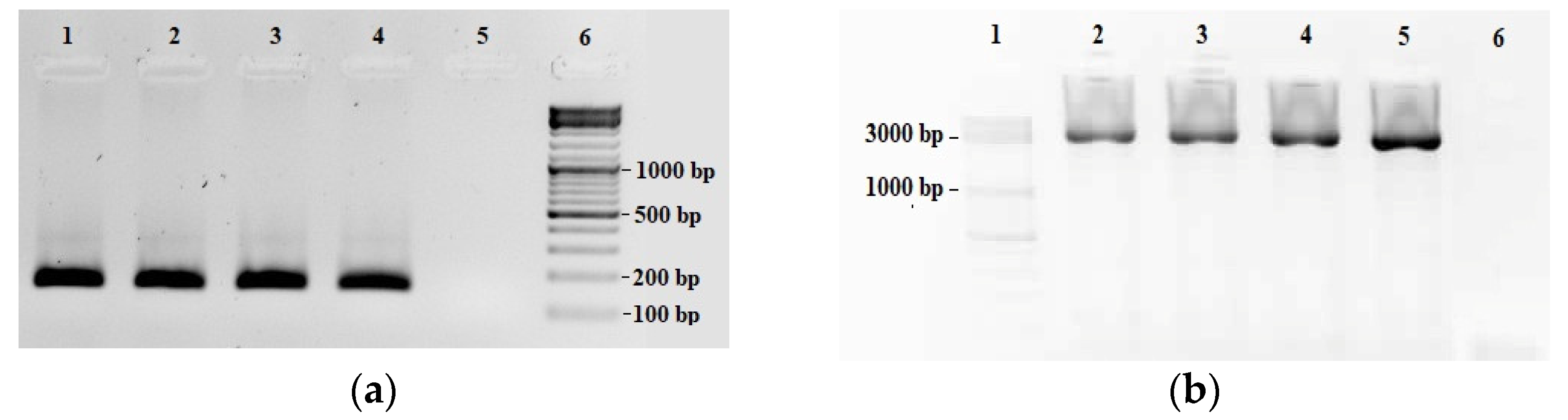
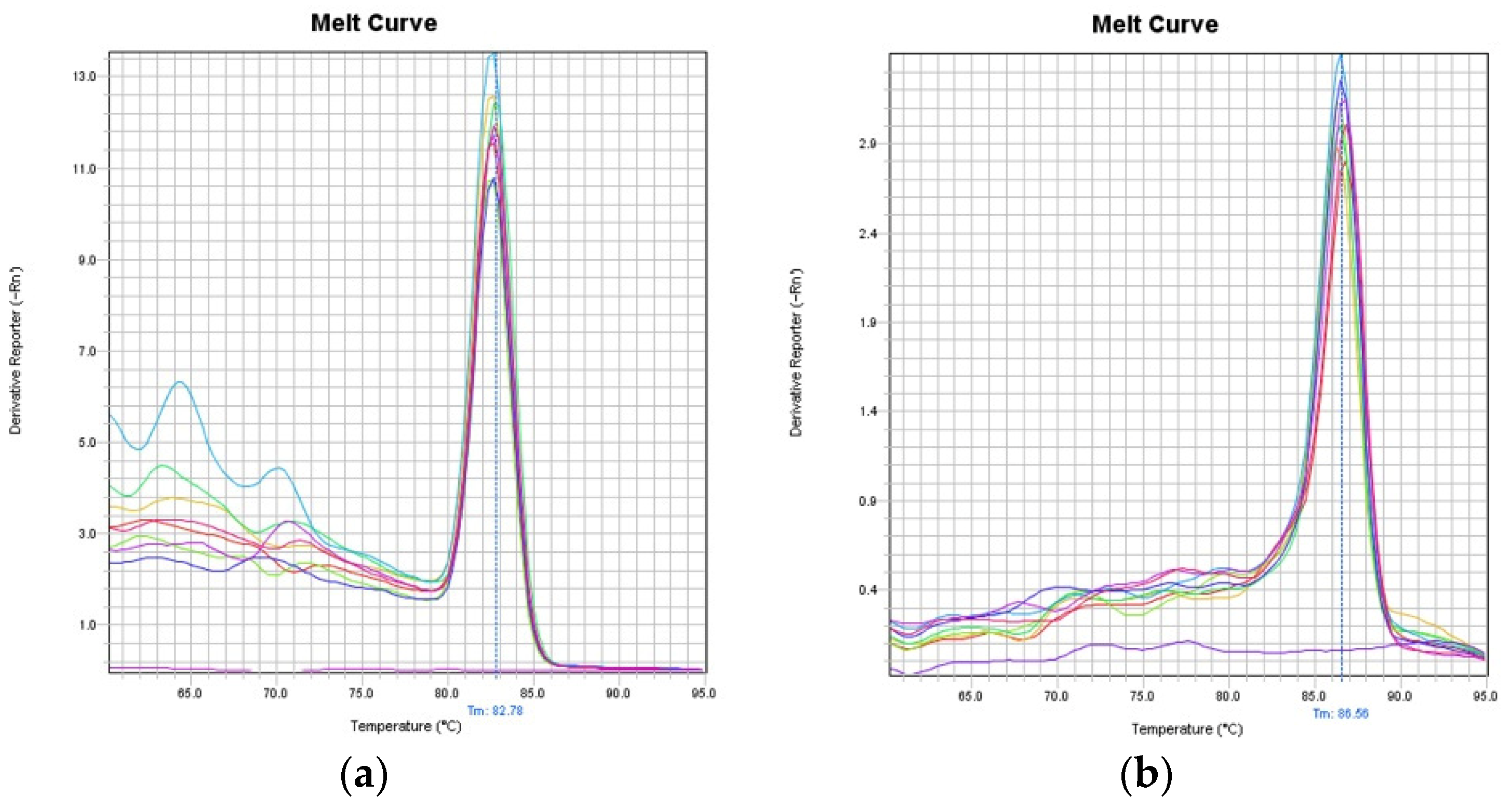
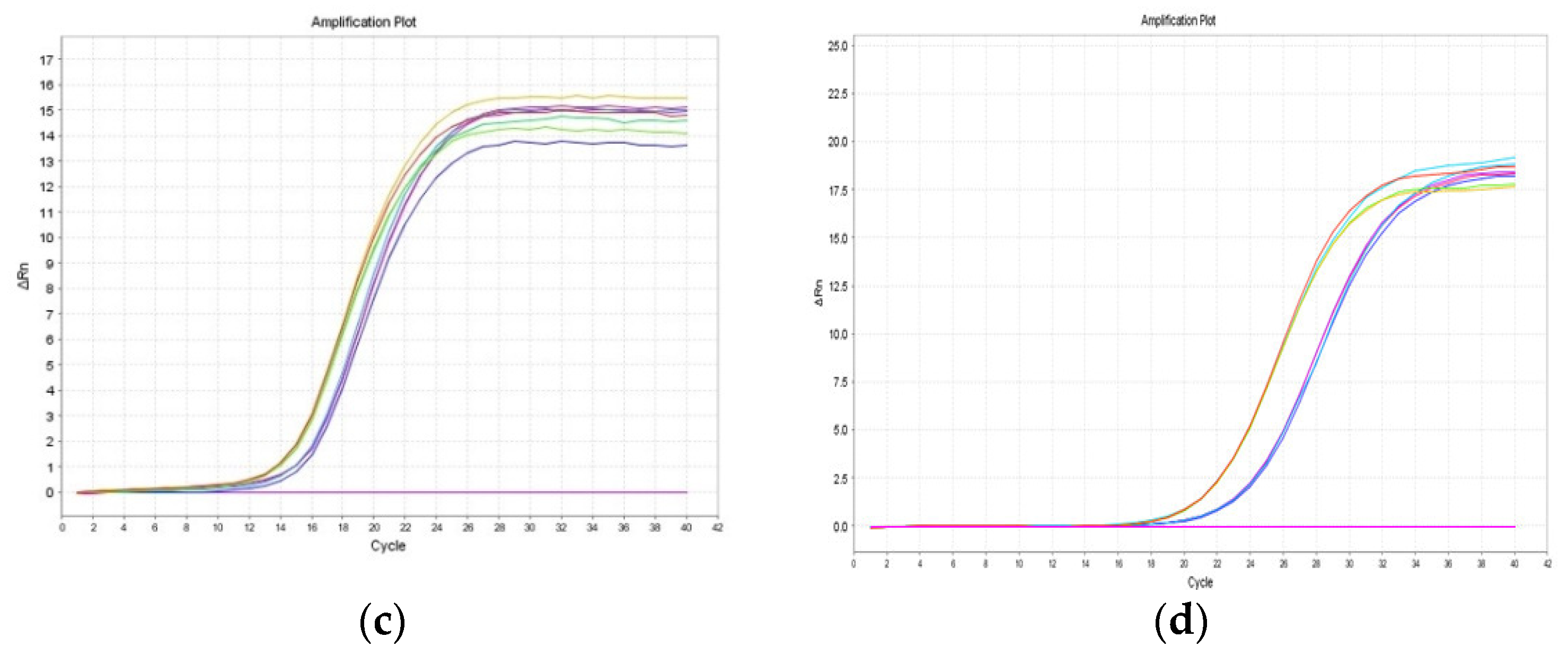
2.1. Validation of the LORD-Q PCR Assay in Common Carp Species
2.2. Mortality and Disorder Results
2.3. Lesion Rates by LORD-Q PCR Assay
3. Discussion
4. Materials and Methods
4.1. Mycotoxin Production and Analysis
4.2. Animal and Experimental Design
4.3. Sampling and DNS Isolation
4.4. LORD-Q PCR Assay
4.5. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, G.; Pettersson, H. Toxicological evaluation of trichothecenes in animal feed. Anim. Feed Sci. Technol. 2004, 114, 205–239. [Google Scholar] [CrossRef]
- Rodrigues, I.; Naehrer, K. Prevalence of mycotoxins in feedstuffs and feed surveyed worldwide in 2009 and 2010. Phytopathol. Mediterr. 2012, 51, 175–192. [Google Scholar]
- Matejova, I.; Faldyna, M.; Modra, H.; Blahova, J.; Palikova, M.; Markova, Z.; Franc, A.; Vicenova, M.; Vojtek, L.; Bartonkova, J.; et al. Effect of T-2 toxin-contaminated diet on common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2017, 60, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, C.; Kersten, S.; Burkhardt-Holm, P.; Valenta, H.; Dänick, S. Occurrence of Deoxynivalenol and Zearalenone in Commercial Fish Feed: An Initial Study. Toxins 2013, 5, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Ti, D.; Wang, N.; Deng, Z.; Zhan, Y.; Liu, W.; Hu, Y.; Liu, T.; Tan, L.; Li, Y.; et al. The protective effect of DL-Selenomethionine against T-2/HT-2 toxins-induced cytotoxicity and oxidative stress in broiler hepatocytes. Toxicol. In Vitro 2019, 54, 137–146. [Google Scholar] [CrossRef]
- Balogh, K.; Heincinger, M.; Fodor, J.; Mézes, M. Effect of long term feeding of T-2 and HT-2 toxin contaminated diet on the glutathione redox status and lipid peroxidation processes in common carp (Cyprinus carpio L.). Acta Biol. Szeged 2009, 53, 23–27. [Google Scholar] [CrossRef]
- Pelyhe, C.S.; Kövesi, B.; Zándoki, E.; Kovács, B.; Szabó-Fodor, J.; Mézes, M. Effect of 4-week feeding of deoxynivalenol or T-2-toxin-contaminated diet on lipid peroxidation and glutathione redox system in the hepatopancreas of common carp (Cyprinus carpio L.). Mycotoxin Res. 2016, 32, 77–83. [Google Scholar] [CrossRef]
- Pietsch, C.; Michel, C.; Kersten, S.; Valenta, H.; Dänick, S.; Schulz, C.; Kloas, W.; Burkhardt-Holm, P. In vivo effects of deoxynivalenol (DON) on innate immune responses of carp (Cyprinus carpio L.). Food Chem. Toxicol. 2014, 68, 44–52. [Google Scholar] [CrossRef]
- Pietsch, C.; Schulz, C.; Rovira, P.; Kloas, W.; Burkhardt-Holm, P. Organ damage and hepatic lipid accumulation in carp (Cyprinus carpio L.) after feed-borne exposure to the mycotoxin, deoxynivalenol (DON). Toxins 2014, 6, 756–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Faragi, J.K. The Efficacy of Prebiotic (β-Glucan) as a Feed Additive against Toxicity of Aflatoxin B1 in Common Carp, Cyprinus Carpio L. J. Aquac. Res. Dev. 2014, 5, 240. [Google Scholar] [CrossRef] [Green Version]
- Lehle, S.; Hildebrand, D.G.; Merz, B.; Malak, P.N.; Becker, M.S.; Schmezer, P.; Essmann, F.; Schulze-Osthoff, K.; Rothfuss, O. LORD-Q: A long-run real-time PCR-based DNA-damage quantification method for nuclear and mitochondrial genome analysis. Nucleic Acids Res. 2013, 42, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wagener, S.; Volker, T.; De Spirt, S.; Ernst, H.; Stahl, W. 3,3′-Dihydroxyisorenieratene and isorenieratene prevent UV-induced DNA damage in human skin fibroblasts. Free Radic. Biol. Med. 2012, 53, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Dannenmann, B.; Lehle, S.; Lorscheid, S.; Huber, S.M.; Essmann, F.; Schulz-Osthoff, K. Simultaneous quantification of DNA-damage and mitochondrial copy number by Long-run DNA-damage quantification (LORD-Q). Oncotarget 2017, 8, 112417–112425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothfuss, O.; Gasser, T.; Patenge, N. Analysis of differential DNA damage in the mitochondrial genome employing a semi-long run real-time PCR approach. Nucleic Acids Res. 2010, 38, e24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Coffmann, J.A. Simple and fast quantification of DNA damage by real-time PCR, and its application to nuclear and mitochondrial DNA from multiple tissues of aging zebrafish. BMC Res. Notes 2017, 10, 269. [Google Scholar] [CrossRef] [Green Version]
- Kainz, P.; Schmiedlechner, A.; Strack, H.B. Specificity-Enhanced Hot-Start PCR: Addition of Double-Stranded DNA Fragments Adapted to the Annealing Temperature. Biotechniques 2018, 28, 2. [Google Scholar] [CrossRef] [Green Version]
- Tubbs, A.; Nussenzweig, A. Endogenous DNA damage as a source of genomic instability in cancer. Cell 2017, 168, 644–656. [Google Scholar] [CrossRef] [Green Version]
- Holton, N.W.; Ebenstein, Y.; Gassman, N.R. Broad spectrum detection of DNA damage by Repair Assisted Damage Detection (RADD). DNA Repair 2018, 66–67, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Anson, R.M.; Mason, P.A.; Bohr, V.A. Gene-specific and mitochondrial repair of oxidative DNA damage. Methods Mol. Biol. 2006, 314, 155–181. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Hou, R.; Hong, X.; Yan, S.; Zha, J. Organophosphate flame retardants (OPFRs) induce genotoxicity in vivo: A survey on apoptosis, DNA methylation, DNA oxidative damage, liver metabolites, and transcriptomics. Environ. Int. 2019, 130, 104914. [Google Scholar] [CrossRef] [PubMed]
- Mondal, N.K.; Bhattacharya, P.; Ray, M.R. Assessment of DNA damage by comet assay and fast halo assay in buccal epithelial cells of Indian women chronically exposed to biomass smoke. Int. J. Hyg. Environ. Health 2011, 214, 311–318. [Google Scholar] [CrossRef]
- Mathew, S.T.; Johansson, P.; Gao, Y.; Fasth, A.; Ek, T.; Hammarsten, O. A flow cytometry assay that measures cellular sensitivity to DNA-damaging agents, customized for clinical routine laboratories. Clin. Biochem. 2016, 49, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Didenko, V.V. (Ed.) DNA Damage Detection In Situ, Ex Vivo, and In Vitro; Springer Protocols, Methods in Molecular Biology; Humna Press: New York, NY, USA, 2011. [Google Scholar]
- Edwards, J.G. Quantification of mitochondrial DNA (mtDNA) damage and error rates by real-time QPCR. Mitochondrion 2009, 9, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Moss, M.O. Mycotoxin review—1. Aspergillus and Penicillium. Mycologist 2002, 16, 116–119. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; Smet, D.D.; De Saeger, S.; Zhang, S. T-2 Toxin, a Trichothecene Mycotoxin: Review of Toxicity, Metabolism, and Analytical Methods. J. Agric. Food Chem. 2011, 59, 3441–3453. [Google Scholar] [CrossRef]
- Anater, A.; Manyes, L.; Meca, G.; Ferrer, E.; Bittencourt, L.F.; Pimpão, C.T.; Font, G. Mycotoxins and their consequences in aquaculture: A review. Aquaculture 2016, 451, 1–10. [Google Scholar] [CrossRef]
- Panigrahi, S. Bioassay of mycotoxins using terrestrial and aquatic, animal and plant species. Food Chem. Toxicol. 1993, 31, 767–790. [Google Scholar] [CrossRef]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef]
- Mostrom, M. Trichothecenes and zearalenone. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 753–763. [Google Scholar] [CrossRef]
- Li, P.; Su, R.; Yin, R.; Lai, D.; Wang, M.; Liu, Y.; Zhou, L. Detoxification of mycotoxins through biotransformation. Toxins 2020, 12, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dizdaroglu, M. Oxidatively induced DNA damage: Mechanisms, repair and disease. Cancer Lett. 2012, 327, 26–47. [Google Scholar] [CrossRef] [PubMed]
- Jena, N.R. DNA damage by reactive species: Mechanisms, mutation and repair. J. Biosci. 2012, 37, 503–517. [Google Scholar] [CrossRef] [PubMed]
- García-Medinan, S.; Núñez-Betancourt, J.A.; García-Medina, A.L.; Galar-Martínez, L.; Neri-Cruz, N.; Islas-Flores, H.; Gómez-Oliván, L.M. The relationship of cytotoxic and genotoxic damage with blood aluminum levels and oxidative stress induced by this metal in common carp (Cyprinus carpio) erythrocytes. Ecotoxicol. Environ. Saf. 2013, 96, 191–197. [Google Scholar] [CrossRef]
- Yang, L.; Tu, D.; Zhai, Z.; Cui, J. Cytotoxicity and apoptosis induced by mixed mycotoxins (T-2 and HT-2 toxin) on primary hepatocytes of broilers in vitro. Toxicon 2017, 129, 1–10. [Google Scholar] [CrossRef]
- Fodor, J.; Németh, M.; Kametler, L.; Pósa, R.; Kovács, M.; Horn, P. Novel methods of Fusarium toxins’ production for toxicological experiments. Acta Agrar. Kaposváriensis 2006, 10, 277–285. [Google Scholar]
- Pussemier, L.; Piérard, J.Y.; Anselme, M.; Tangni, E.K.; Motte, J.C.; Larondelle, Y. Development and application of analytical methods for the determination of mycotoxins in organic and conventional wheat. Food Addit. Contam. 2006, 23, 1208–1218. [Google Scholar] [CrossRef]
- Trebstein, A.; Seefelder, W.; Lauber, U.; Humpf, H.U. Determination of T-2 and HT-2 toxins in cereals including oats after immunoaffinity cleanup by liquid chromatography and fluorescence detection. J. Agric. Food Chem. 2008, 56, 4968–4975. [Google Scholar] [CrossRef]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [Green Version]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 26 July 2013).




{kind=link}
{kind=link}
{kind=link}
| Control | DON | T-2 | |
|---|---|---|---|
| Week 1 | 0 | 1 | 1 |
| Week 2 | 0 | 3 | 3 |
| Week 3 | 2 | 4 | 4 |
| Mortality rate % | 5.5 | 16.7 | 27.8 |
| Liver necrosis | 0% | 100% | 100% |
| Intestine inflammation | 0% | 100% | 100% |
| Week | Control | T-2 | DON | ||
|---|---|---|---|---|---|
| Mean | Mean | S.D. | Mean | S.D. | |
| 1. | 0 A | 1.33 bB | 0.22 | 1.87 bC | 0.42 |
| 2. | 0 A | 0.74 aB | 0.06 | 1.47 abC | 0.43 |
| 3. | 0 A | 0.68 aA | 0.05 | 0.82 aA | 0.27 |
| Primer Name | Sequence (5′ to 3′) | Amplicon Size (bp) | Position on the mt Genome (bp) | PCR Efficiency % | R2 |
|---|---|---|---|---|---|
| F2-R4.R | AAGCACGGATCAGACGAACA | 156 | 5887–6042 | 94 | 0.99 |
| F2-R4.F | CACGCAGGAGCATCAGTAGA | ||||
| F3-R2.R | TAGGCTGGATAATAGGGTTGC | 3436 | 647–4082 | 86 | 0.98 |
| F3-R2.F | CCGTTCAACCTCACCACTTCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabó, R.T.; Kovács-Weber, M.; Balogh, K.M.; Mézes, M.; Kovács, B. Changes of DNA Damage Effect of T-2 or Deoxynivalenol Toxins during Three Weeks Exposure in Common Carp (Cyprinus carpio L.) Revealed by LORD-Q PCR. Toxins 2021, 13, 576. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080576
Szabó RT, Kovács-Weber M, Balogh KM, Mézes M, Kovács B. Changes of DNA Damage Effect of T-2 or Deoxynivalenol Toxins during Three Weeks Exposure in Common Carp (Cyprinus carpio L.) Revealed by LORD-Q PCR. Toxins. 2021; 13(8):576. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080576
Chicago/Turabian StyleSzabó, Rubina Tünde, Mária Kovács-Weber, Krisztián Milán Balogh, Miklós Mézes, and Balázs Kovács. 2021. "Changes of DNA Damage Effect of T-2 or Deoxynivalenol Toxins during Three Weeks Exposure in Common Carp (Cyprinus carpio L.) Revealed by LORD-Q PCR" Toxins 13, no. 8: 576. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080576