Origin and Characterization of Extracellular Vesicles Present in the Spider Venom of Ornithoctonus hainana

 , , and
, , and
Abstract
:1. Introduction
2. Results
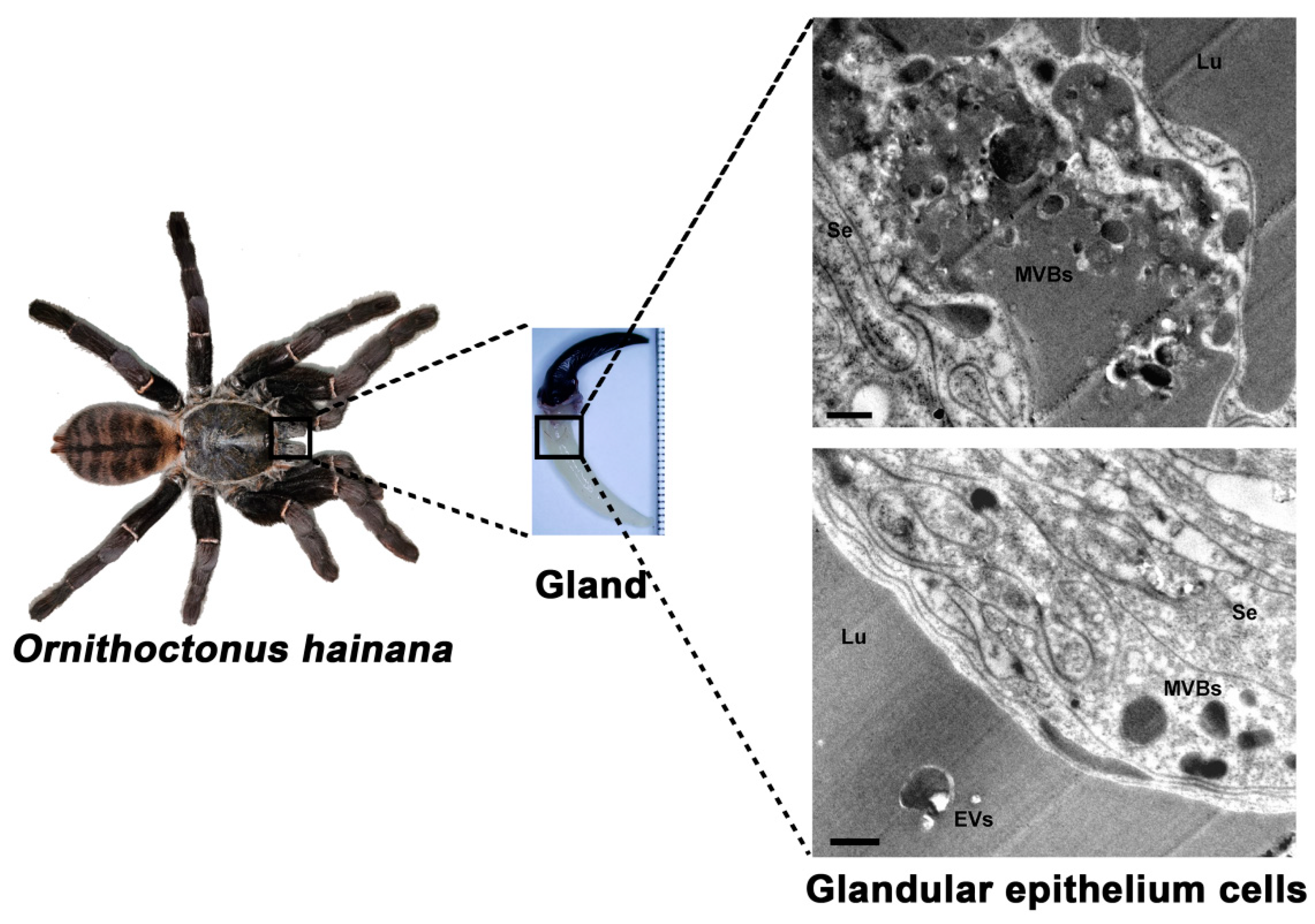
2.1. Ultrastructural Analysis of the Glandular Epithelium Cells of the Ornithoctonus hainana Venom Gland by TEM
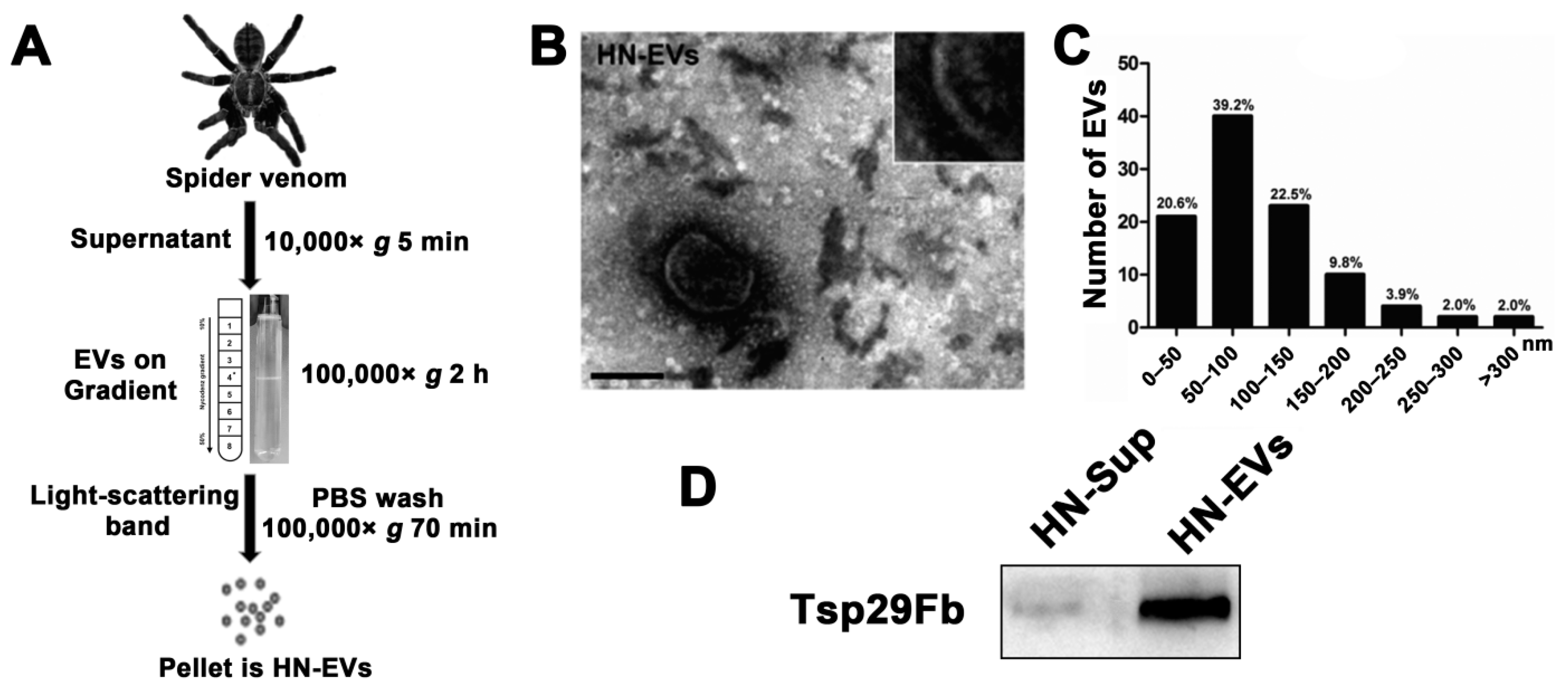
2.2. Isolation and Characterization of Secreted EVs from the Venom of Ornithoctonus hainana
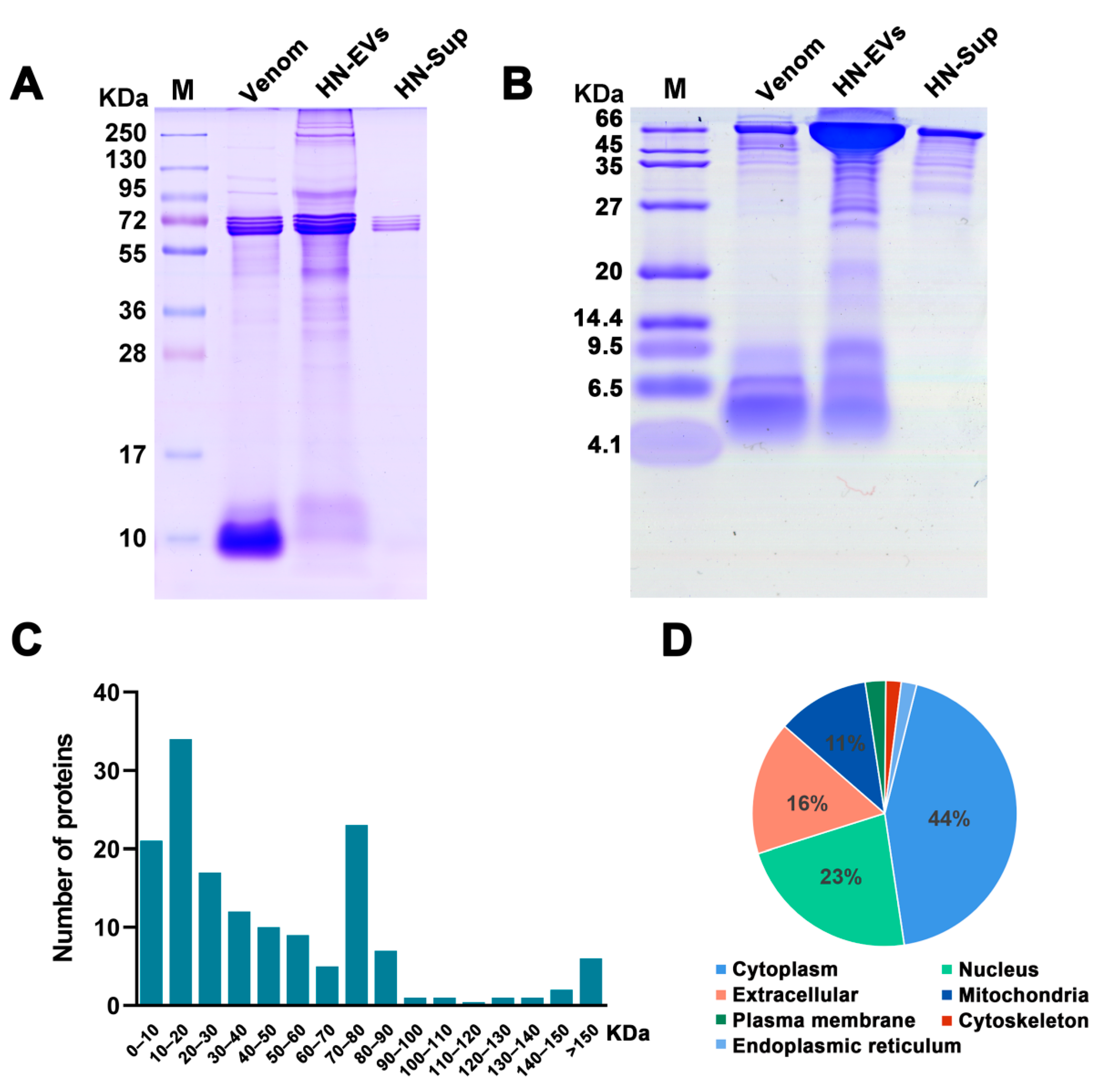
2.3. Analysis and Identification of HN-EV Proteins
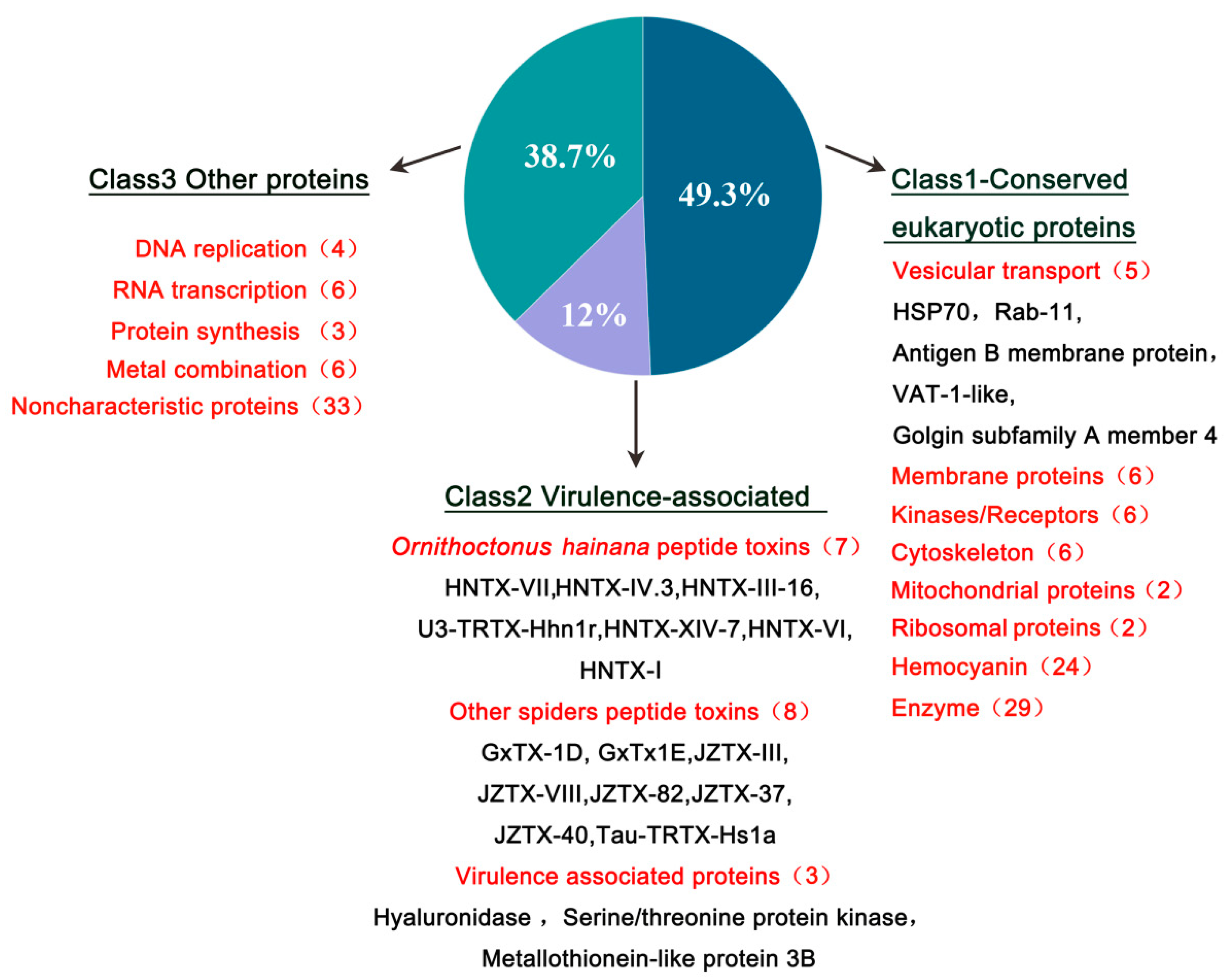
2.4. Functional Classification of HN-EV Proteins
2.5. Validation of Hyaluronidase Activity
2.6. HN-EVs Inhibit Viability of HUVECs
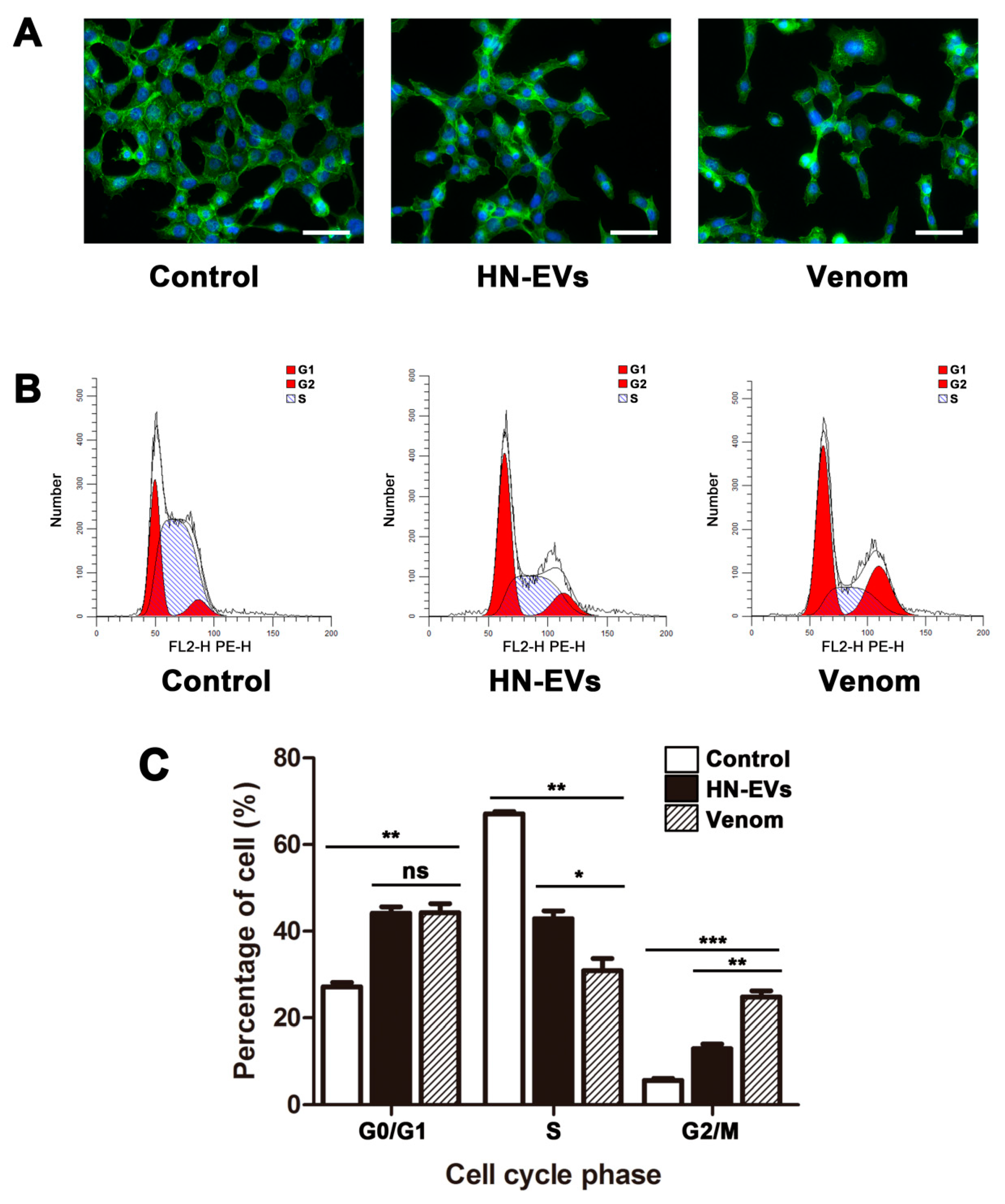
2.7. The Influence of HN-EVs on the Cell Cytoskeleton and Cell Cycle of HUVECs
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sample Preparation
5.2. Purification of EVs in Spider Venom
5.3. Transmission Electron Microscopy
5.4. SDS-PAGE and Western Blotting Assay
5.5. Sample Preparation and Digestion
5.6. MS/MS Analysis of Venom HN-EV Proteins
5.7. Database Search and Bioinformatic Analysis
5.8. Hyaluronic Acid Hydrolysis Assay
5.9. HN-EV Uptake Assay
5.10. Cell Viability Assays
5.11. F-Actin Organizational Changes
5.12. Cell Cycle Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dimitrov, D.; Lopardo, L.; Giribet, G.; Arnedo, M.A.; Alvarez-Padilla, F.; Hormiga, G. Tangled in a sparse spider web: Single origin of orb weavers and their spinning work unravelled by denser taxonomic sampling. Proc. Biol. Sci. 2012, 279, 1341–1350. [Google Scholar] [CrossRef] [PubMed]
- Rash, L.D.; Hodgson, W.C. Pharmacology and biochemistry of spider venoms. Toxicon 2002, 40, 225–254. [Google Scholar] [CrossRef]
- Langenegger, N.; Nentwig, W.; Kuhn-Nentwig, L. Spider Venom: Components, Modes of Action, and Novel Strategies in Transcriptomic and Proteomic Analyses. Toxins 2019, 11, 611. [Google Scholar] [CrossRef] [Green Version]
- Escoubas, P.; Diochot, S.; Corzo, G. Structure and pharmacology of spider venom neurotoxins. Biochimie 2000, 82, 893–907. [Google Scholar] [CrossRef]
- Saez, N.J.; Senff, S.; Jensen, J.E.; Er, S.Y.; Herzig, V.; Rash, L.D.; King, G.F. Spider-venom peptides as therapeutics. Toxins 2010, 2, 2851–2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, G.F.; Hardy, M.C. Spider-venom peptides: Structure, pharmacology, and potential for control of insect pests. Annu. Rev. Entomol. 2013, 58, 475–496. [Google Scholar] [CrossRef] [PubMed]
- Coakley, G.; Maizels, R.M.; Buck, A.H. Exosomes and Other Extracellular Vesicles: The New Communicators in Parasite Infections. Trends Parasitol. 2015, 31, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Ofir-Birin, Y.; Regev-Rudzki, N. Extracellular vesicles in parasite survival. Science 2019, 363, 817–818. [Google Scholar] [CrossRef]
- Maas, S.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [Green Version]
- Eichenberger, R.M.; Sotillo, J.; Loukas, A. Immunobiology of parasitic worm extracellular vesicles. Immunol. Cell Biol. 2018, 96, 704–713. [Google Scholar] [CrossRef]
- Tulkens, J.; De Wever, O.; Hendrix, A. Analyzing bacterial extracellular vesicles in human body fluids by orthogonal biophysical separation and biochemical characterization. Nat. Protoc. 2020, 15, 40–67. [Google Scholar] [CrossRef]
- Zwarycz, A.S.; Livingstone, P.G.; Whitworth, D.E. Within-species variation in OMV cargo proteins: The Myxococcus xanthus OMV pan-proteome. Mol. Omics 2020, 16, 387–397. [Google Scholar] [CrossRef]
- Tassetto, M.; Kunitomi, M.; Andino, R. Circulating Immune Cells Mediate a Systemic RNAi-Based Adaptive Antiviral Response in Drosophila. Cell 2017, 169, 314–325.e13. [Google Scholar] [CrossRef] [Green Version]
- Vora, A.; Zhou, W.; Londono-Renteria, B.; Woodson, M.; Sherman, M.B.; Colpitts, T.M.; Neelakanta, G.; Sultana, H. Arthropod EVs mediate dengue virus transmission through interaction with a tetraspanin domain containing glycoprotein Tsp29Fb. Proc. Natl. Acad. Sci. USA 2018, 115, E6604–E6613. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Woodson, M.; Neupane, B.; Bai, F.; Sherman, M.B.; Choi, K.H.; Neelakanta, G.; Sultana, H. Exosomes serve as novel modes of tick-borne flavivirus transmission from arthropod to human cells and facilitates dissemination of viral RNA and proteins to the vertebrate neuronal cells. PLoS Pathog. 2018, 14, e1006764. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, S.M.; Fernandes, W.; Sant’Anna, S.S.; Yamanouye, N. Microvesicles in the venom of Crotalus durissus terrificus (Serpentes, Viperidae). Toxicon 2007, 49, 106–110. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kanai-Azuma, M.; Akimoto, Y.; Kawakami, H.; Yanoshita, R. Exosome-like vesicles in Gloydius blomhoffii blomhoffii venom. Toxicon 2008, 51, 984–993. [Google Scholar] [CrossRef]
- Souza-Imberg, A.; Carneiro, S.M.; Giannotti, K.C.; Sant’Anna, S.S.; Yamanouye, N. Origin and characterization of small membranous vesicles present in the venom of Crotalus durissus terrificus. Toxicon 2017, 136, 27–33. [Google Scholar] [CrossRef]
- Heavner, M.E.; Ramroop, J.; Gueguen, G.; Ramrattan, G.; Dolios, G.; Scarpati, M.; Kwiat, J.; Bhattacharya, S.; Wang, R.; Singh, S.; et al. Novel Organelles with Elements of Bacterial and Eukaryotic Secretion Systems Weaponize Parasites of Drosophila. Curr. Biol. 2017, 27, 2869–2877.e6. [Google Scholar] [CrossRef]
- Wan, B.; Goguet, E.; Ravallec, M.; Pierre, O.; Lemauf, S.; Volkoff, A.N.; Gatti, J.L.; Poirié, M. Venom Atypical Extracellular Vesicles as Interspecies Vehicles of Virulence Factors Involved in Host Specificity: The Case of a Drosophila Parasitoid Wasp. Front. Immunol. 2019, 10, 1688. [Google Scholar] [CrossRef] [Green Version]
- Ramroop, J.R.; Heavner, M.E.; Razzak, Z.H.; Govind, S. A parasitoid wasp of Drosophila employs preemptive and reactive strategies to deplete its host’s blood cells. PLoS Pathog. 2021, 17, e1009615. [Google Scholar] [CrossRef] [PubMed]
- Carregari, V.C.; Rosa-Fernandes, L.; Baldasso, P.; Bydlowski, S.P.; Marangoni, S.; Larsen, M.R.; Palmisano, G. Snake Venom Extracellular vesicles (SVEVs) reveal wide molecular and functional proteome diversity. Sci. Rep. 2018, 8, 12067. [Google Scholar] [CrossRef] [PubMed]
- dos Santo, V.L.; Franco, C.R.; Viggiano, R.L.; da Silveira, R.B.; Cantão, M.P.; Mangili, O.C.; Veiga, S.S.; Gremski, W. Structural and ultrastructural description of the venom gland of Loxosceles intermedia (brown spider). Toxicon 2000, 38, 265–285. [Google Scholar] [CrossRef]
- Malli, H.; Kuhn-Nentwig, L.; Imboden, H.; Moon, M.J.; Wyler, T. Immunocytochemical localization and secretion process of the toxin CSTX-1 in the venom gland of the wandering spider Cupiennius salei (Araneae: Ctenidae). Cell Tissue Res. 2000, 299, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Yigit, N.; Guven, T. Functional structure of Agelena labyrinthica’s (Araneae:Agelenidae) venom gland and electrophoresis of venom. Toxicon 2006, 47, 58–67. [Google Scholar] [CrossRef]
- Rocha-e-Silva, T.A.A.; Collares-Buzato, C.B.; Cruz-Höfling, M.A.; Hyslop, S. Venom apparatus of the Brazilian tarantula Vitalius dubius Mello-Leitão 1923 (Theraphosidae). Cell Tissue Res. 2009, 335, 617–629. [Google Scholar] [CrossRef]
- Schmidtberg, H.; von Reumont, B.M.; Lemke, S.; Vilcinskas, A.; Lüddecke, T. Morphological Analysis Reveals a Compartmentalized Duct in the Venom Apparatus of the Wasp Spider (Argiope bruennichi). Toxins 2021, 13, 270. [Google Scholar] [CrossRef]
- Kowal, J.; Tkach, M.; Théry, C. Biogenesis and secretion of exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Xun, C.; Ge, L.; Tang, F.; Wang, L.; Zhuo, Y.; Long, L.; Qi, J.; Hu, L.; Duan, D.; Chen, P.; et al. Insight into the proteomic profiling of exosomes secreted by human OM-MSCs reveals a new potential therapy. Biomed. Pharm. 2020, 131, 110584. [Google Scholar] [CrossRef]
- Xun, C.; Ge, L.; Tang, F.; Wang, L.; Zhuo, Y.; Long, L.; Qi, J.; Hu, L.; Duan, D.; Chen, P.; et al. Molecular diversification of peptide toxins from the tarantula Haplopelma hainanum (Ornithoctonus hainana) venom based on transcriptomic, peptidomic, and genomic analyses. J. Proteome Res. 2010, 9, 2550–2564. [Google Scholar]
- Zhang, Y.Y.; Huang, Y.; He, Q.Z.; Luo, J.; Zhu, L.; Lu, S.S.; Liu, J.Y.; Huang, P.F.; Zeng, X.Z.; Liang, S.P. Structural and Functional Diversity of Peptide Toxins from Tarantula Haplopelma hainanum (Ornithoctonus hainana) Venom Revealed by Transcriptomic, Peptidomic, and Patch Clamp Approaches. J. Biol. Chem. 2015, 290, 14192–14207. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.C.; Long, R.W.; Wu, Y.Q.; Guo, Y.B.; Liu, D.L.; Peng, L.; Li, D.Q.; Yang, D.W.; Xu, X.; Liu, F.X.; et al. Identification and characterization of toxins in the venom gland of the Chinese bird spider, Haplopelma hainanum, by transcriptomic analysis. Insect Sci. 2016, 23, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Tahir, F.; Wang, J.C.; Woodson, M.; Sherman, M.B.; Karim, S.; Neelakanta, G.; Sultana, H. Discovery of Exosomes from Tick Saliva and Salivary Glands Reveals Therapeutic Roles for CXCL12 and IL-8 in Wound Healing at the Tick-Human Skin Interface. Front. Cell Dev. Biol. 2020, 8, 554. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.H.; Yao, L.L.; Liu, C.; Li, X.D. Regulation of Myosin-5b by Rab11a and the Rab11 family interacting protein 2. Biosci. Rep. 2019, 39, BSR20181252. [Google Scholar] [CrossRef] [Green Version]
- Liao, Z.; Cao, J.; Li, S.; Yan, X.; Hu, W.; He, Q.; Chen, J.; Tang, J.; Xie, J.; Liang, S. Proteomic and peptidomic analysis of the venom from Chinese tarantula Chilobrachys jingzhao. Proteomics 2007, 7, 1892–1907. [Google Scholar] [CrossRef]
- Rodríguez-Rios, L.; Díaz-Peña, L.F.; Lazcano-Pérez, F.; Arreguín-Espinosa, R.; Rojas-Molina, A.; García-Arredondo, A. Hyaluronidase-like enzymes are a frequent component of venoms from theraphosid spiders. Toxicon 2017, 136, 34–43. [Google Scholar] [CrossRef]
- Chaves-Moreira, D.; Matsubara, F.H.; Schemczssen-Graeff, Z.; De Bona, E.; Heidemann, V.R.; Guerra-Duarte, C.; Gremski, L.H.; Chávez-Olórtegui, C.; Senff-Ribeiro, A.; Chaim, O.M.; et al. Brown Spider (Loxosceles) Venom Toxins as Potential Biotools for the Development of Novel Therapeutics. Toxins 2019, 11, 355. [Google Scholar] [CrossRef] [Green Version]
- Khamtorn, P.; Rungsa, P.; Jangpromma, N.; Klaynongsruang, S.; Daduang, J.; Tessiri, T.; Daduang, S. Partial proteomic analysis of brown widow spider (Latrodectus geometricus) venom to determine the biological activities. Toxicon X 2020, 8, 100062. [Google Scholar] [CrossRef]
- Di Nicola, M.R.; Pontara, A.; Kass, G.; Kramer, N.I.; Avella, I.; Pampena, R.; Mercuri, S.R.; Dorne, J.; Paolino, G. Vipers of Major clinical relevance in Europe: Taxonomy, venom composition, toxicology and clinical management of human bites. Toxicology 2021, 453, 152724. [Google Scholar] [CrossRef]
- Biner, O.; Trachsel, C.; Moser, A.; Kopp, L.; Langenegger, N.; Kämpfer, U.; von Ballmoos, C.; Nentwig, W.; Schürch, S.; Schaller, J.; et al. Isolation, N-glycosylations and Function of a Hyaluronidase-Like Enzyme from the Venom of the Spider Cupiennius salei. PLoS ONE 2015, 10, e0143963. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; Im, H.; Castro, C.M.; Breakefield, X.; Weissleder, R.; Lee, H. New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 2018, 118, 1917–1950. [Google Scholar] [CrossRef]
- Nyffeler, M.; Knörnschild, M. Bat predation by spiders. PLoS ONE 2013, 8, e58120. [Google Scholar] [CrossRef] [Green Version]
- Pineda, S.S.; Chin, Y.K.; Undheim, E.; Senff, S.; Mobli, M.; Dauly, C.; Escoubas, P.; Nicholson, G.M.; Kaas, Q.; Guo, S.; et al. Structural venomics reveals evolution of a complex venom by duplication and diversification of an ancient peptide-encoding gene. Proc. Natl. Acad. Sci. USA 2020, 117, 11399–11408. [Google Scholar] [CrossRef]
- Kuhn-Nentwig, L.; Stöcklin, R.; Nentwig, W. Venom Composition and Strategies in Spiders. Adv. Insect Phys. 2011, 40, 1–86. [Google Scholar]
- Pekár, S.; Bočánek, O.; Michálek, O.; Petráková, L.; Haddad, C.R.; Šedo, O.; Zdráhal, Z. Venom gland size and venom complexity-essential trophic adaptations of venomous predators: A case study using spiders. Mol. Ecol. 2018, 27, 4257–4269. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L.; Langenegger, N.; Heller, M.; Koua, D.; Nentwig, W. The Dual Prey-Inactivation Strategy of Spiders-In-Depth Venomic Analysis of Cupiennius salei. Toxins 2019, 11, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizki, R.M.; Rizki, T.M. Parasitoid virus-like particles destroy Drosophila cellular immunity. Proc. Natl. Acad. Sci. USA 1990, 87, 8388–8392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Xiao, Y.; Hu, W.; Xie, J.; Bosmans, F.; Tytgat, J.; Liang, S. Function and solution structure of hainantoxin-I, a novel insect sodium channel inhibitor from the Chinese bird spider Selenocosmia hainana. FEBS Lett. 2003, 555, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Klint, J.K.; Chin, Y.K.; Mobli, M. Rational Engineering Defines a Molecular Switch That Is Essential for Activity of Spider-Venom Peptides against the Analgesics Target NaV1.7. Mol. Pharm. 2015, 88, 1002–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.L.; Yi, S.; Liang, S.P. Mechanism of action of two insect toxins huwentoxin-III and hainantoxin-VI on voltage-gated sodium channels. J. Zhejiang Univ. Sci. B 2010, 11, 451–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Dai, J.; Chen, Z.; Hu, W.; Xiao, Y.; Liang, S. Isolation and characterization of hainantoxin-IV, a novel antagonist of tetrodotoxin-sensitive sodium channels from the Chinese bird spider Selenocosmia hainana. Cell. Mol. Life Sci. 2003, 60, 972–978. [Google Scholar] [CrossRef]
- Wullschleger, B.; Kuhn-Nentwig, L.; Tromp, J.; Kämpfer, U.; Schaller, J.; Schürch, S.; Nentwig, W. CSTX-13, a highly synergistically acting two-chain neurotoxic enhancer in the venom of the spider Cupiennius salei (Ctenidae). Proc. Natl. Acad. Sci. USA 2004, 101, 11251–11256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clémençon, B.; Kuhn-Nentwig, L.; Langenegger, N.; Kopp, L.; Peigneur, S.; Tytgat, J.; Nentwig, W.; Lüscher, B.P. Neurotoxin Merging: A Strategy Deployed by the Venom of the Spider Cupiennius salei to Potentiate Toxicity on Insects. Toxins 2020, 12, 250. [Google Scholar] [CrossRef] [PubMed]
- Elsharkasy, O.M.; Nordin, J.Z.; Hagey, D.W.; de Jong, O.G.; Schiffelers, R.M.; Andaloussi, S.E.; Vader, P. Extracellular vesicles as drug delivery systems: Why and how. Adv. Drug Deliv. Rev. 2020, 159, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Hu, Z.; Zhou, X.; Chen, J.; Tang, C.; Xiao, Z.; Ying, D.; Liu, Z.; Liang, S. The venom of the spider Selenocosmia jiafu contains various neurotoxins acting on voltage-gated ion channels in rat dorsal root ganglion neurons. Toxins 2014, 6, 988–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Accession | Gene | Description |
|---|---|---|---|
| Vesicular transport associated proteins | A0A4Y2J3E9 | RAB11FIP3 | Rab11 family-interacting protein 3 |
| A0A4Y2IWS4 | HSP70B2_4 | Heat shock protein 70 B2 | |
| A0A4Y2H3W6 | Vat1l_2 | Synaptic vesicle membrane protein VAT-1-like | |
| A0A2P6KJS2 | NCL1_37780 | Golgin subfamily A member 4 | |
| A0A4Y2J0H4 | AVEN_242130_1 | Putative antigen B membrane protein | |
| Ornithoctonushainana peptide toxins | D2Y2C3 | HNTX-VII | U5-theraphotoxin-Hhn1a |
| D2Y2D7 | HNTX-IV.3 | Mu-theraphotoxin-Hhn1b 3 | |
| D2Y2I6 | HNTX-III-16 | Mu-theraphotoxin-Hhn2i | |
| P0CH72 | U3-TRTX-Hhn1r | U3-theraphotoxin-Hhn1r | |
| D2Y2E6 | HNTX-XIV-7 | U8-theraphotoxin-Hhn1f | |
| P0CH70 | HNTX-VI | U1-theraphotoxin-Hhn1a | |
| D2Y1X8 | HNTX-I | Mu-theraphotoxin-Hhn2b 3 | |
| Other spiders peptide toxins | P84835 | GxTx1E | Kappa-theraphotoxin-Pg1a |
| B1P1A6 | JZTX-VIII | U3-theraphotoxin-Cg1a | |
| P0CH43 | Tau-TRTX-Hs1a | Tau-theraphotoxin-Hs1a | |
| B1P1K1 | JZTX-82 | Secretory protein | |
| P84836 | GxTX-1D | Kappa-theraphotoxin-Pg1b | |
| B1P1D8 | JZTX-37 | U13-theraphotoxin-Cg1b | |
| B1P1G5 | JZTX-40 | U21-theraphotoxin-Cg1c | |
| P62520 | JZTX-III | Beta/kappa-theraphotoxin-Cg1a | |
| Virulence associated proteins | A0A2L2YBK4 | Hyal | Hyaluronidase |
| A0A2P6KXQ8 | NCL1_24342 | Serine/threonine protein kinase-like | |
| A0A4Y2HPX8 | AVEN_37080_1 | Metallothionein-like protein 3B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xun, C.; Wang, L.; Yang, H.; Xiao, Z.; Deng, M.; Xu, R.; Zhou, X.; Chen, P.; Liu, Z. Origin and Characterization of Extracellular Vesicles Present in the Spider Venom of Ornithoctonus hainana. Toxins 2021, 13, 579. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080579
Xun C, Wang L, Yang H, Xiao Z, Deng M, Xu R, Zhou X, Chen P, Liu Z. Origin and Characterization of Extracellular Vesicles Present in the Spider Venom of Ornithoctonus hainana. Toxins. 2021; 13(8):579. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080579
Chicago/Turabian StyleXun, Chengfeng, Lu Wang, Hailin Yang, Zixuan Xiao, Min Deng, Rongfang Xu, Xi Zhou, Ping Chen, and Zhonghua Liu. 2021. "Origin and Characterization of Extracellular Vesicles Present in the Spider Venom of Ornithoctonus hainana" Toxins 13, no. 8: 579. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080579