Biocontrol Agents Reduce Progression and Mycotoxin Production of Fusarium graminearum in Spikelets and Straws of Wheat

 , , , and
, , , and
Abstract
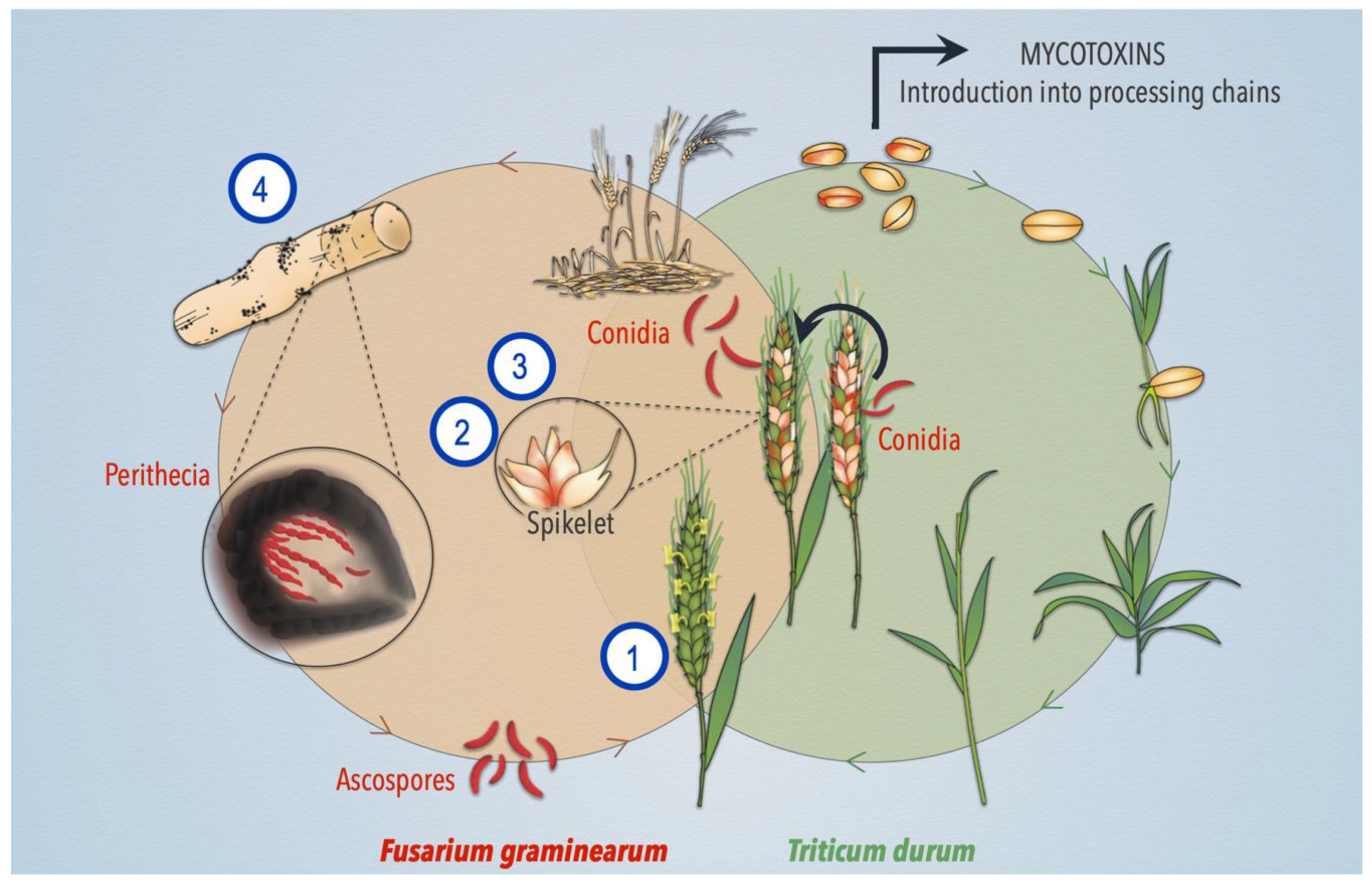
:1. Introduction
2. Results
2.1. Wheat Spikelet Colonization by Microorganisms
2.2. Inhibition of F. graminearum External Colonization during Antagonist Assays in Spikelet
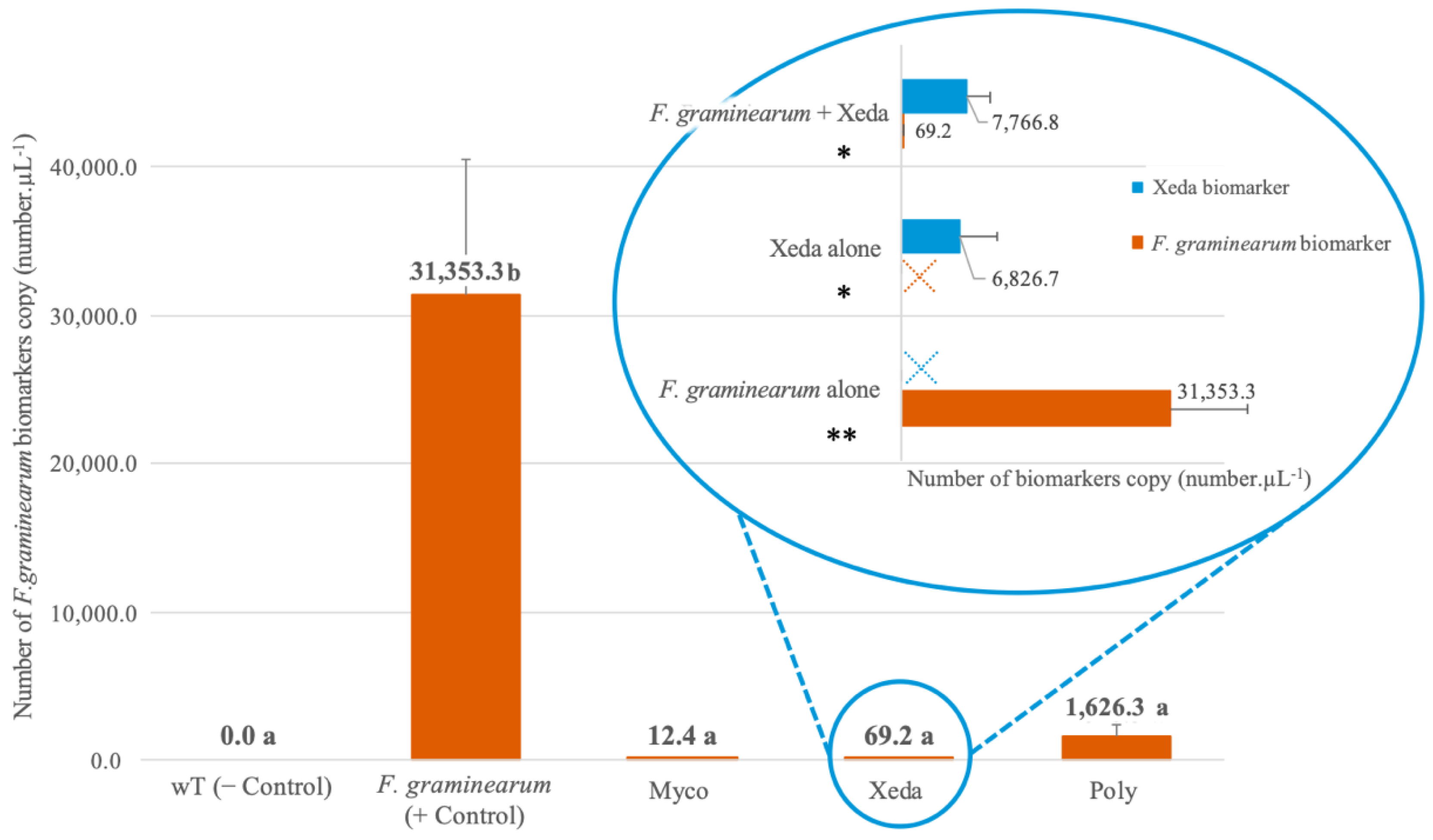
2.3. Quantification of F. graminearum Inhibition and Xeda–Pathogen Interaction in Internal Wheat Spikelet Tissues
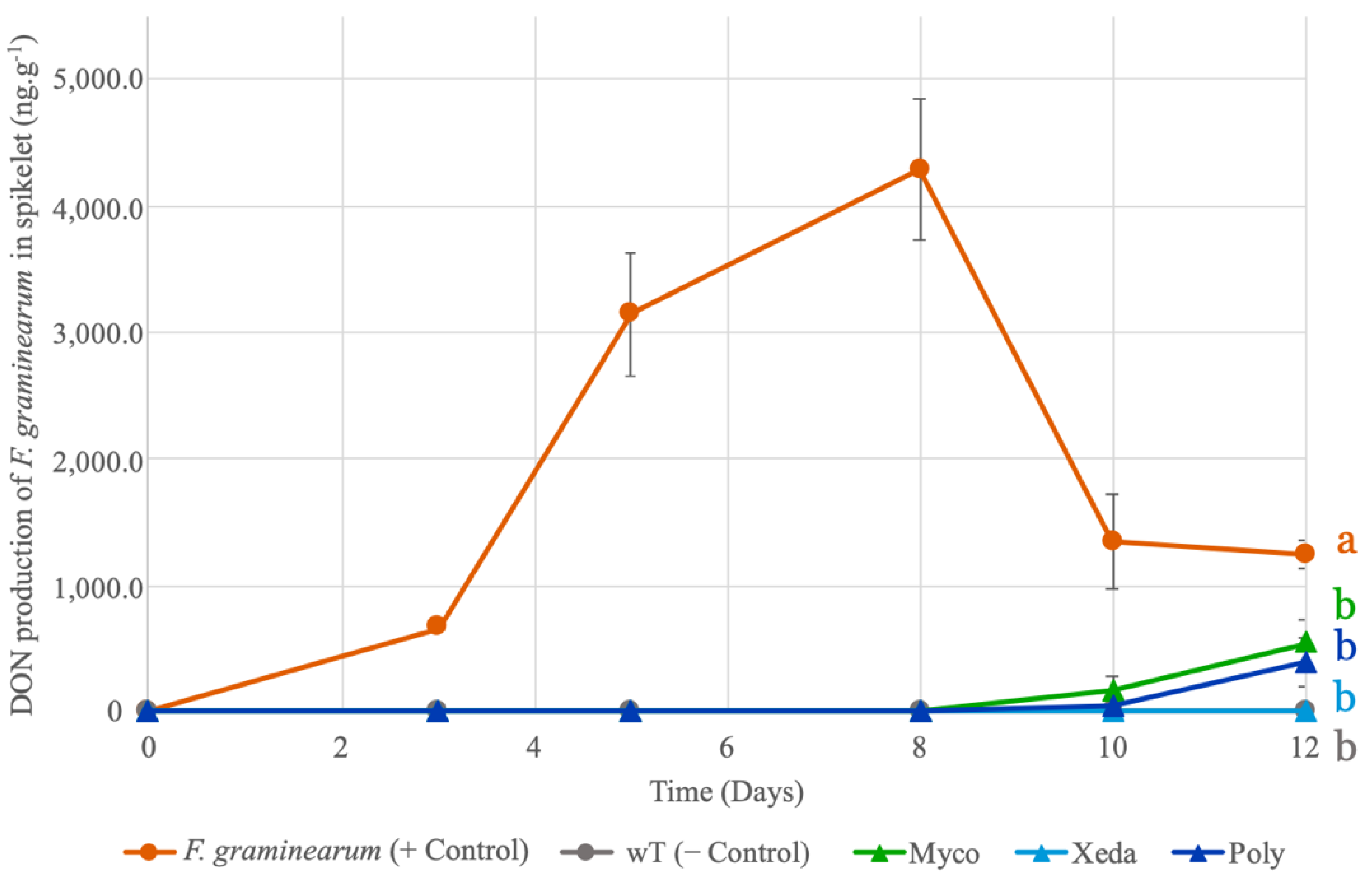
2.4. Inhibition of DON Production by F. graminearum during Antagonist Assay in Spikelet
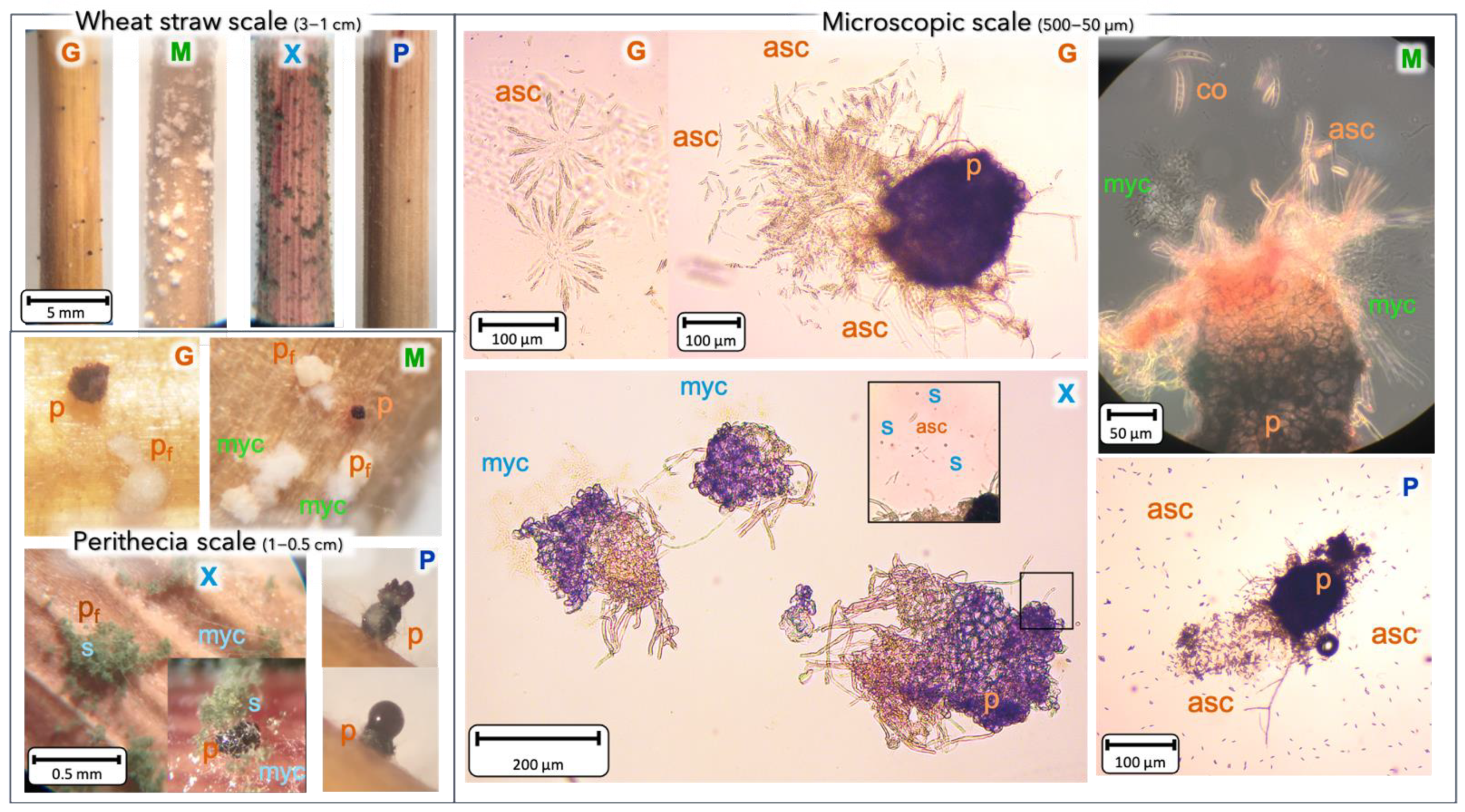
2.5. BCAs Impact on Perithecia Synthesis and Quantity on Wheat Straw
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Micro-Organisms and Culture Condition
5.2. Antagonistic Activity of BCAs on Wheat Spikelets
5.2.1. Inoculation and Visual Growth Assessment of Wheat Spikelet
5.2.2. Quantification of BCAs-Pathogens Interaction in Wheat Spikelet Tissues by ddPCR
5.2.3. DON, 15-AcDON and 3-AcDON Extraction and Quantification on Wheat Spikelet
5.2.4. Antagonistic Activity on Wheat Straw and F. graminearum Perithecia Production
5.2.5. Data Treatment and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium Toxins in Cereals: Occurrence, Legislation, Factors Promoting the Appearance and Their Management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Goswami, R.S.; Kistler, H. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarch, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From prevention in the field to detoxification by adsorption or transformation. Food Addit. Contam. Part A 2011, 28, 1590–1609. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Kabir, E.; Jahan, S.A. Exposure to pesticides and the associated human health effects. Sci. Total Environ. 2017, 575, 525–535. [Google Scholar] [CrossRef]
- Torres, A.M.; Palacios, S.; Yerkovich, N.; Palazzini, J.M.; Battilani, P.; Leslie, J.; Logrieco, A.; Chulze, S. Fusarium head blight and mycotoxins in wheat: Prevention and control strategies across the food chain. World Mycotoxin J. 2019, 12, 333–355. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2017, 63, 39–59. [Google Scholar] [CrossRef] [Green Version]
- van der Lee, T.; Zhang, H.; van Diepeningen, A.; Waalwijk, C. Biogeography of Fusarium graminearum species complex and chemotypes: A review. Food Addit. Contam. Part A 2014, 32, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Llorens, A.; Mateo, R.; Hinojo, M.; Valle-Algarra, F.; Jiménez, M. Influence of environmental factors on the biosynthesis of type B trichothecenes by isolates of Fusarium spp. from Spanish crops. Int. J. Food Microbiol. 2004, 94, 43–54. [Google Scholar] [CrossRef]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Boedi, S.; Berger, H.; Sieber, C.; Münsterkötter, M.; Maloku, I.; Warth, B.; Sulyok, M.; Lemmens, M.; Schuhmacher, R.; Güldener, U.; et al. Comparison of Fusarium graminearum Transcriptomes on Living or Dead Wheat Differentiates Substrate-Responsive and Defense-Responsive Genes. Front. Microbiol. 2016, 7, 1113. [Google Scholar] [CrossRef] [PubMed]
- Bujold, I.; Paulitz, T.; Carisse, O. Effect of Microsphaeropsis sp. on the Production of Perithecia and Ascospores of Gibberella zeae. Plant Dis. 2001, 85, 977–984. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, B.K.; Bakker, M.G. Bacterial endophyte antagonism toward a fungal pathogenin vitrodoes not predict protection in live plant tissue. FEMS Microbiol. Ecol. 2018, 95, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Profizi, C.; Baillieul, F. Screening of wheat endophytes as biological control agents against Fusarium head blight using two different in vitro tests. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Beck, B.; Spanoghe, P.; Moens, M.; Brusselman, E.; Temmerman, F.; Pollet, S.; Nuyttens, D. Improving the biocontrol potential of Steinernema feltiae against Delia radicum through dosage, application technique and timing. Pest Manag. Sci. 2013, 70, 841–851. [Google Scholar] [CrossRef]
- Rojas, E.C.; Jensen, B.; Jørgensen, H.J.L.; Latz, M.A.C.; Esteban, P.; Ding, Y.; Collinge, D.B. Selection of fungal endophytes with biocontrol potential against Fusarium head blight in wheat. Biol. Control 2020, 144, 104222. [Google Scholar] [CrossRef]
- Pellan, L.; Durand, N.; Martinez, V.; Fontana, A.; Schorr-Galindo, S.; Strub, C. Commercial Biocontrol Agents Reveal Contrasting Comportments Against Two Mycotoxigenic Fungi in Cereals: Fusarium Graminearum and Fusarium Verticillioides. Toxins 2020, 12, 152. [Google Scholar] [CrossRef] [Green Version]
- Xue, A.G.; Chen, Y.; Voldeng, H.D.; Fedak, G.; Savard, M.E.; Längle, T.; Zhang, J.; Harman, G.E. Concentration and cultivar effects on efficacy of CLO-1 biofungicide in controlling Fusarium head blight of wheat. Biol. Control 2014, 73, 2–7. [Google Scholar] [CrossRef]
- Siou, D.; Gelisse, S.; Laval, V.; Repincay, C.; Canalès, R.; Suffert, F.; Lannou, C. Effect of wheat spike infection timing on fusarium head blight development and mycotoxin accumulation. Plant Pathol. 2013, 63, 390–399. [Google Scholar] [CrossRef]
- Foroud, N.A.; Pordel, R.; Goyal, R.K.; Ryabova, D.; Eranthodi, A.; Chatterton, S.; Kovalchuk, I. Chemical Activation of the Ethylene Signaling Pathway Promotes Fusarium graminearum Resistance in Detached Wheat Heads. Phytopathology 2019, 109, 796–803. [Google Scholar] [CrossRef]
- Kheiri, A.; Jorf, S.A.M.; Malihipour, A. Infection process and wheat response to Fusarium head blight caused by Fusarium graminearum. Eur. J. Plant Pathol. 2018, 153, 489–502. [Google Scholar] [CrossRef]
- Vargas, W.A.; Mandawe, J.C.; Kenerley, C.M. Plant-Derived Sucrose Is a Key Element in the Symbiotic Association between Trichoderma virens and Maize Plants. Plant Physiol. 2009, 151, 792–808. [Google Scholar] [CrossRef] [Green Version]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Váňová, M.; Tvarůžek, L.; Hrabalová, H. Reaction of winter wheat varieties to Fusarium graminearum and F. culmorum in field infection trials and the efficacy of fungicides. Plant Prot. Sci. 2001, 37, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhou, M.; Zhang, Z.; Ren, L.; Du, L.; Zhang, B.; Xu, H.; Xin, Z. Expression of a radish defensin in transgenic wheat confers increased resistance to Fusarium graminearum and Rhizoctonia cerealis. Funct. Integr. Genom. 2011, 11, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Bekalu, Z.E.; Madsen, C.K.; Dionisio, G.; Holme, I.B.; Jørgensen, L.N.; Fomsgaard, I.S.; Brinch-Pedersen, H. Overexpression of Nepenthesin HvNEP-1 in Barley Endosperm Reduces Fusarium Head Blight and Mycotoxin Accumulation. Agronomy 2020, 10, 203. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Aguirre, I.; Rački, N.; Dreo, T.; Ravnikar, M. Droplet Digital PCR for Absolute Quantifi cation of Pathogens. Plant Pathol. 2015, 1302, 331–347. [Google Scholar]
- Selvaraj, V.; Maheshwari, Y.; Hajeri, S.; Yokomi, R. Droplet Digital PCR for Absolute Quantification of Plant Pathogens. In Plant Biotechnology: Progress in Genomic Era; Springer: Singapore, 2019; pp. 583–595. [Google Scholar] [CrossRef]
- Brown, N.A.; Evans, J.; Mead, A.; Hammond-Kosack, K.E. A spatial temporal analysis of the Fusarium graminearum transcriptome during symptomless and symptomatic wheat infection. Mol. Plant Pathol. 2017, 18, 1295–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatesh, N.; Keller, N.P. Mycotoxins in Conversation with Bacteria and Fungi. Front. Microbiol. 2019, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, J.; Yang, N.; Wen, Z.; Sun, X.; Chai, Y.; Ma, Z. Wheat microbiome bacteria can reduce virulence of a plant pathogenic fungus by altering histone acetylation. Nat. Commun. 2018, 9, 3429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyprianou, M. Commission Regulation (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- Pellan, L.; Dieye, C.; Durand, N.; Fontana, A.; Strub, C.; Schorr-Galindo, S. Biocontrol Agents: Toolbox for the Screening of Weapons against Mycotoxigenic Fusarium. J. Fungi 2021, 7, 446. [Google Scholar] [CrossRef]
- Gilbert, J.; Fernando, W.G.D. Epidemiology and biological control of Gibberella zeae/Fusarium graminearum. Can. J. Plant Pathol. 2004, 26, 464–472. [Google Scholar] [CrossRef]
- Inch, S.; Walker, D.J.; Gilbert, J.; Daayf, F.; Fernando, W.G.D.; Piercey-Normore, M. The development of a model to predict the potential efficacy of Trichoderma harzianumisolates on perithecial production of Gibberella zeaebased on secondary metabolite production. Can. J. Plant Pathol. 2011, 33, 337–346. [Google Scholar] [CrossRef]
- Rabiey, M.; Ullah, I.; Shaw, L.J.; Shaw, M.W. Potential ecological effects of Piriformospora indica, a possible biocontrol agent, in UK agricultural systems. Biol. Control 2017, 104, 1–9. [Google Scholar] [CrossRef]
- Correa, O.S.; Montecchia, M.S.; Berti, M.F.; Ferrari, M.C.F.; Pucheu, N.L.; Kerber, N.L.; García, A.F. Bacillus amyloliquefaciens BNM122, a potential microbial biocontrol agent applied on soybean seeds, causes a minor impact on rhizosphere and soil microbial communities. Appl. Soil Ecol. 2009, 41, 185–194. [Google Scholar] [CrossRef]
- Edel-Hermann, V.; Brenot, S.; Gautheron, N.; Aime, S.; Alabouvette, C.; Steinberg, C. Ecological fitness of the biocontrol agent Fusarium oxysporum Fo47 in soil and its impact on the soil microbial communities. FEMS Microbiol. Ecol. 2009, 68, 37–45. [Google Scholar] [CrossRef]
- Leplat, J.; Friberg, H.; Abid, M.; Steinberg, C. Survival of Fusarium Graminearum, the Causal Agent of Fusarium Head Blight. A Review. Agron. Sustain. Dev. 2013, 33, 97–111. [Google Scholar] [CrossRef]
- Marín, P.; Jurado, M.; Magan, N.; Vázquez, C.; Teresa González-Jaén, M. Effect of Solute Stress and Temperature on Growth Rate and TRI5 Gene Expression Using Real Time RT-PCR in Fusarium Graminearum from Spanish Wheat. Int. J. Food Microbiol. 2010, 140, 169–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerin, D.; Pollastro, S.; Raguseo, C.; Angelini, R.M.D.M.; Faretra, F. A Ready-to-Use Single- and Duplex-TaqMan-QPCR Assay to Detect and Quantify the Biocontrol Agents Trichoderma Asperellum and Trichoderma Gamsii. Front. Microbiol. 2018, 9, 2073. [Google Scholar] [CrossRef] [PubMed]
- Manstretta, V.; Rossi, V. Effects of Temperature and Moisture on Development of Fusarium graminearum Perithecia in Maize Stalk Residues. Appl. Environ. Microbiol. 2016, 82, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Amplicon Size | Targeted Organisms | Source | |
|---|---|---|---|---|---|
| Tri5 | F | GATCTGATGACTACCCTCAATTCCTT | 71 | F. graminearum | [40] |
| R | GCCATAGAGAAGCCCCAACAC | ||||
| RNA PolyB subII (rpb2) | F | GGAGGTCGTTGAGGAGTACGAA | 142 | Xeda (T. asperellum) | [41] |
| R | TTGCAGATAGGATTTACGACGAGT | ||||
| IS | Polarity | MRM | EC |
|---|---|---|---|
| U-(13C15)-DEOXYNIVALENOL | − | 370.3 > 59.0 | 35 |
| Polarity | MRM Q | EC MRM Q | MRM q | EC MRM q | R2 Calibration Curve | |
|---|---|---|---|---|---|---|
| DON | − | 355.0 > 59.0 | 35 | 355.0 > 265.1 | 35 | 0.9998 |
| 15-AcDON | + | 338.90 > 137.10 | −19 | 338.90 > 297.20 | −14 | 0.9988 |
| 3-AcDON | − | 397.20 > 59.00 | 25 | 397.20 > 307.10 | 16 | 0.9988 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pellan, L.; Dieye, C.A.T.; Durand, N.; Fontana, A.; Schorr-Galindo, S.; Strub, C. Biocontrol Agents Reduce Progression and Mycotoxin Production of Fusarium graminearum in Spikelets and Straws of Wheat. Toxins 2021, 13, 597. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090597
Pellan L, Dieye CAT, Durand N, Fontana A, Schorr-Galindo S, Strub C. Biocontrol Agents Reduce Progression and Mycotoxin Production of Fusarium graminearum in Spikelets and Straws of Wheat. Toxins. 2021; 13(9):597. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090597
Chicago/Turabian StylePellan, Lucile, Cheikh Ahmeth Tidiane Dieye, Noël Durand, Angélique Fontana, Sabine Schorr-Galindo, and Caroline Strub. 2021. "Biocontrol Agents Reduce Progression and Mycotoxin Production of Fusarium graminearum in Spikelets and Straws of Wheat" Toxins 13, no. 9: 597. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090597