BmK86-P1, a New Degradation Peptide with Desirable Thermostability and Kv1.2 Channel-Specific Activity from Traditional Chinese Scorpion Medicinal Material

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
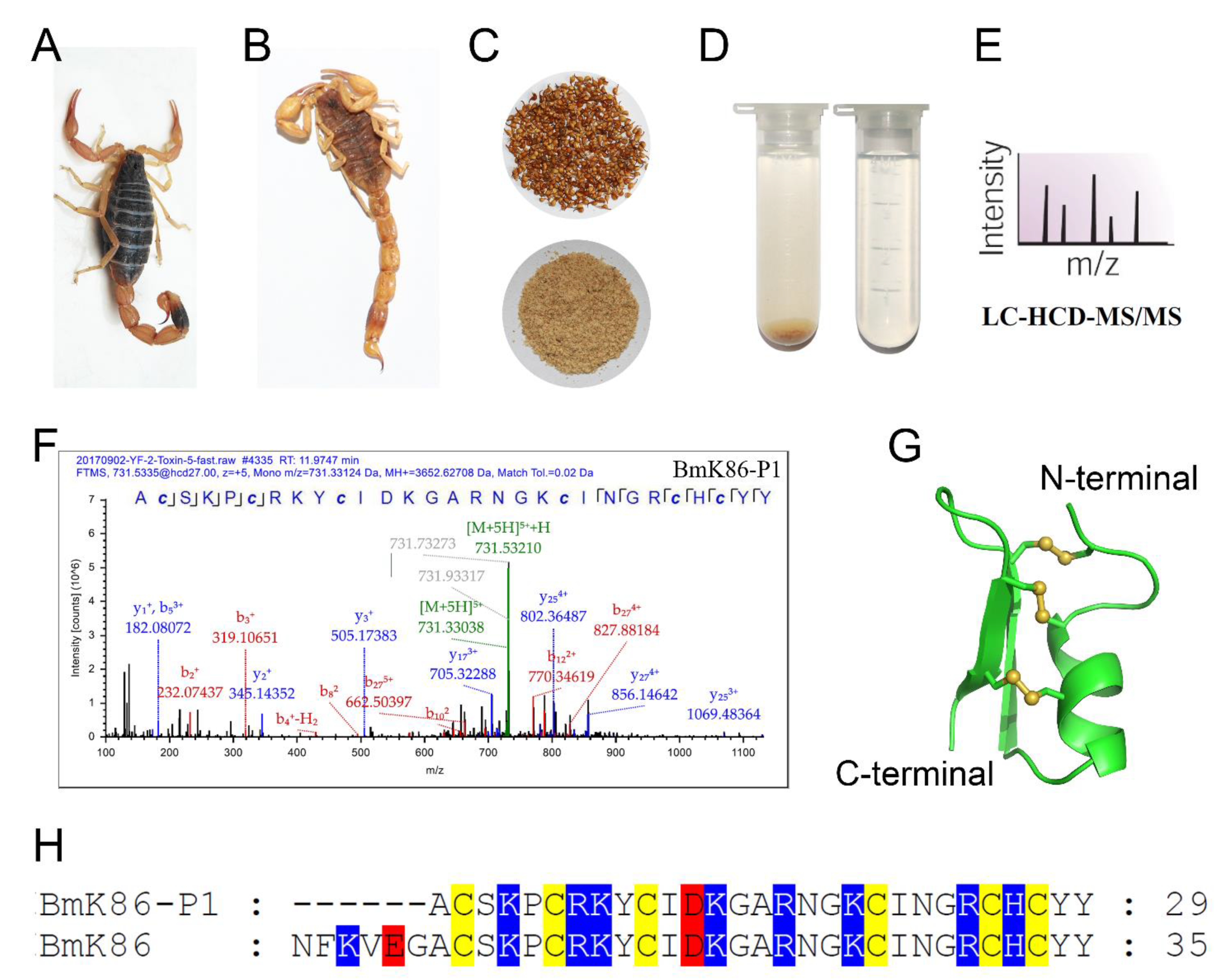
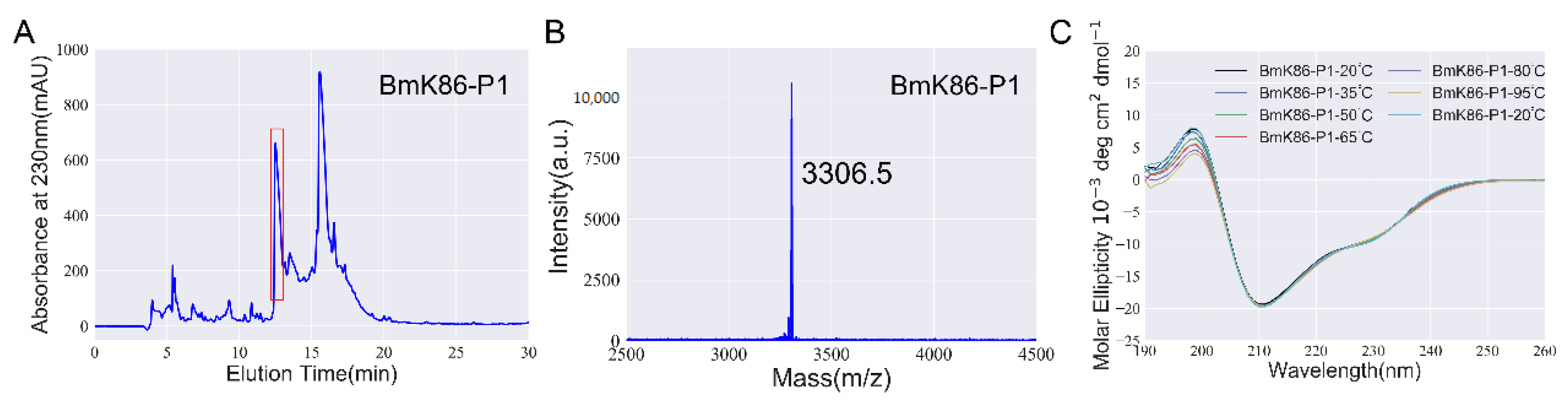
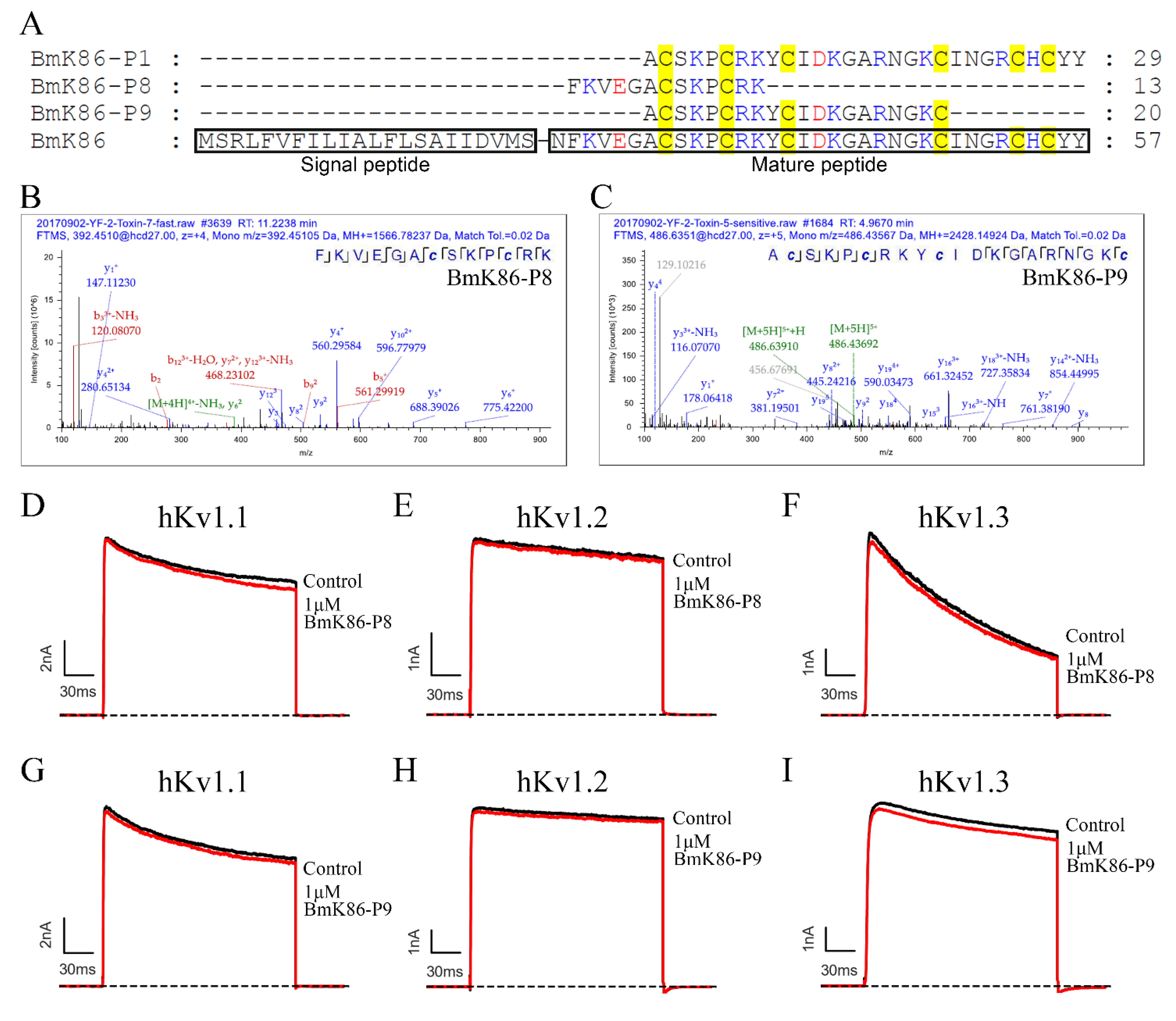
2.1. Identification of a New Peptide BmK86-P1 from Scorpion Medicinal Material
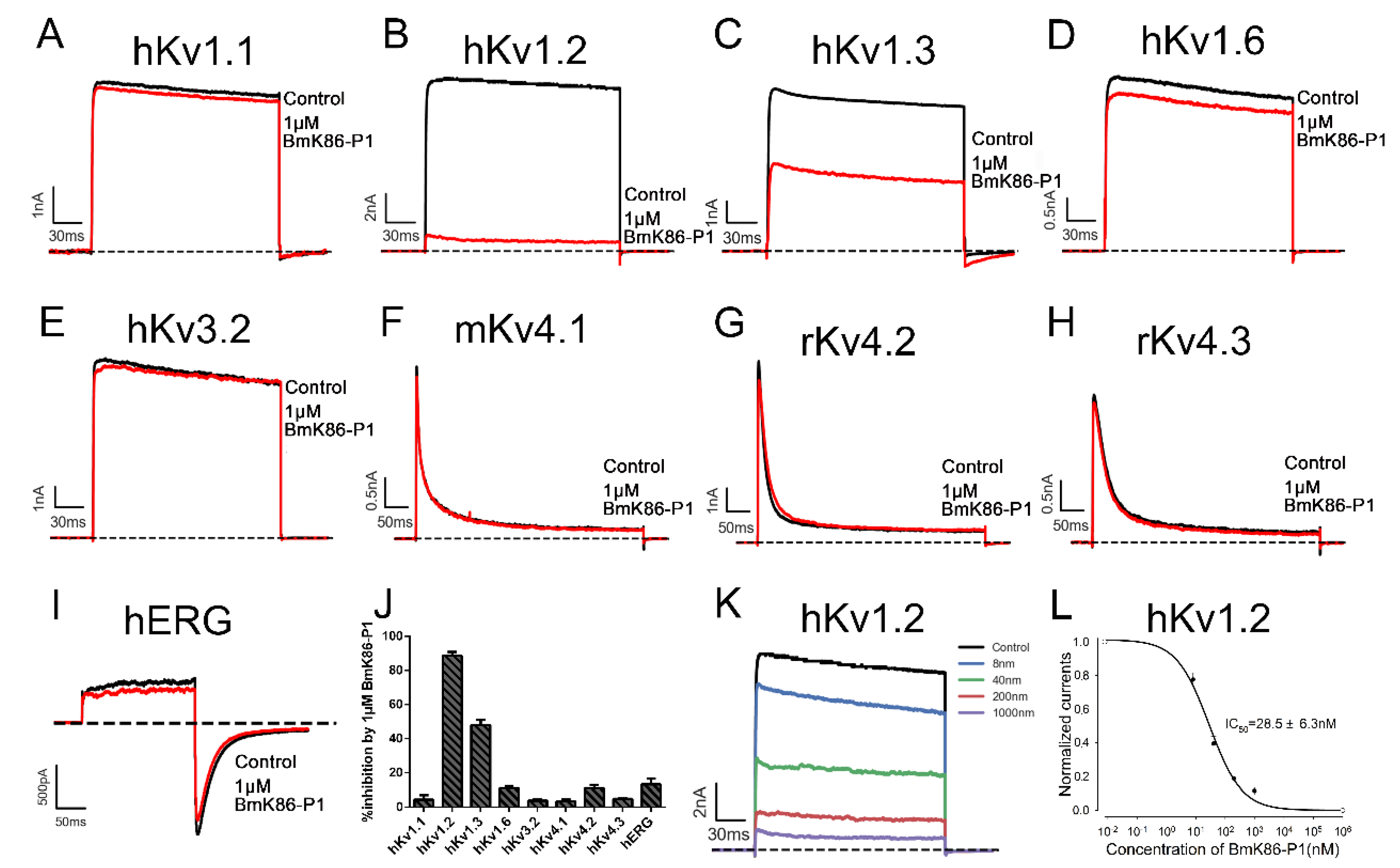
2.2. Electrophysiological Activities of BmK86-P1
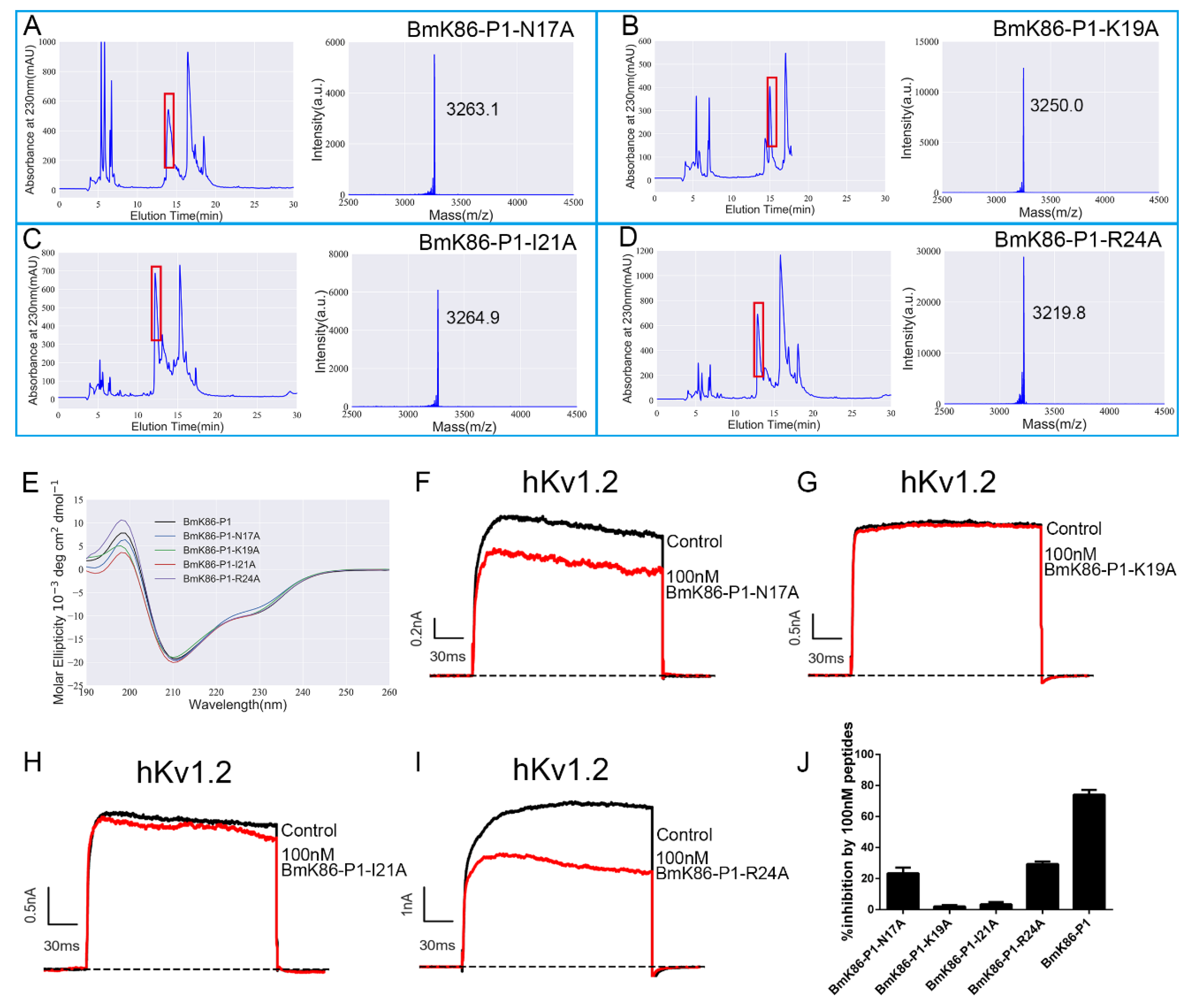
2.3. Functional Residues in the Channel Pore Region Responsible for BmK86-P1 Binding
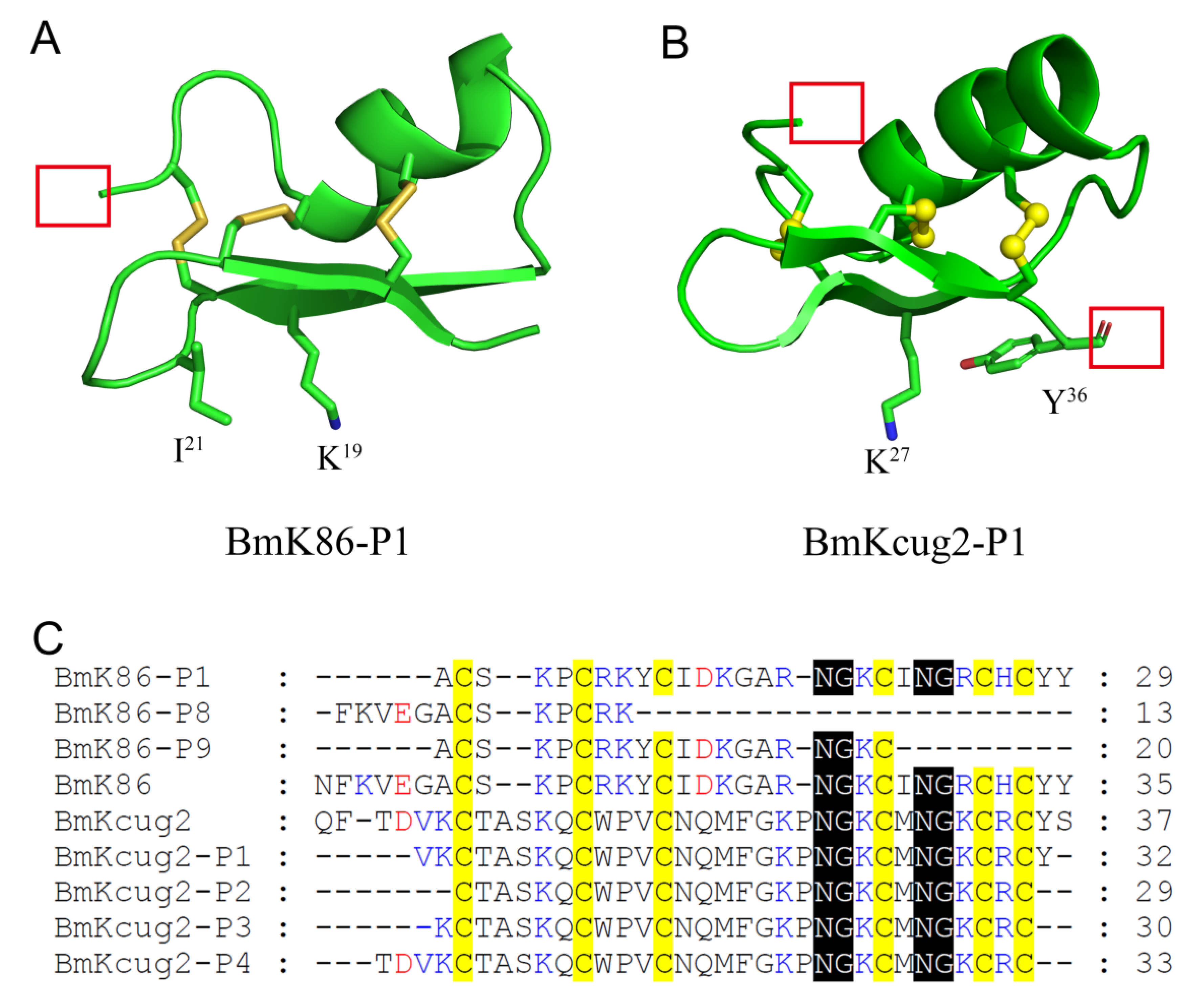
2.4. Functional Sites of BmK86-P1 Identified by Alanine-Scanning Mutagenesis
2.5. The Absence of the Conserved Cysteine Residue Pattern and Key Residue Deletion in Two New Analogs of BmK86 Results in the Loss of Inhibitory Activities to Potassium Channels
3. Discussion
4. Materials and Methods
4.1. LC-MS/MS Analysis
4.2. Construction of Expression Vector
4.3. Expression and Purification of Peptides
4.4. Potassium Channels and Cell Culture and Transfection
4.5. Circular Dichroism (CD) Spectroscopy
4.6. Electrophysiological Recordings
4.7. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chinese Pharmacopoeia Comission. Chinese Pharmacopoeia (Part I); China Medical Science and Technology Press: Beijing, China, 2020; p. 143.
- Yang, F.; Wang, D.; Tong, Y.; Qin, C.; Yang, L.; Yu, F.; Huang, X.; Liu, S.; Cao, Z.; Guo, L.; et al. Thermostable potassium channel-inhibiting neurotoxins in processed scorpion medicinal material revealed by proteomic analysis: Implications of its pharmaceutical basis in traditional Chinese medicine. J. Proteom. 2019, 206, 103435. [Google Scholar] [CrossRef]
- Qin, C.; Wan, X.; Li, S.; Yang, F.; Yang, L.; Zuo, Z.; Cao, Z.; Chen, Z.; Wu, Y. Different pharmacological properties between scorpion toxin BmKcug2 and its degraded analogs highlight the diversity of K+ channel blockers from thermally processed scorpions. Int. J. Biol. Macromol. 2021, 178, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Syrbe, S.; Hedrich, U.B.S.; Riesch, E.; Djemie, T.; Muller, S.; Moller, R.S.; Maher, B.; Hernandez-Hernandez, L.; Synofzik, M.; Caglayan, H.S.; et al. De novo loss- or gain-of-function mutations in KCNA2 cause epileptic encephalopathy. Nat. Genet. 2015, 47, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Corbett, M.A.; Bellows, S.T.; Li, M.; Carroll, R.; Micallef, S.; Carvill, G.L.; Myers, C.T.; Howell, K.B.; Maljevic, S.; Lerche, H.; et al. Dominant KCNA2 mutation causes episodic ataxia and pharmacoresponsive epilepsy. Neurology 2016, 87, 1975–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, G.; Guo, J.; Zhang, Z.; Zhang, S.; Zhu, Y.; Cheng, J.; Yu, L.; Ji, Y.; Tao, J. Kv1.3 Channel as a Key Therapeutic Target for Neuroinflammatory Diseases: State of the Art and Beyond. Front. Neurosci. 2019, 13, 1393. [Google Scholar] [CrossRef] [Green Version]
- Gubič, S.; Hendrickx, L.A.; Toplak, Z.; Sterle, M.; Peigneur, S.; Tomašič, T.; Pardo, L.A.; Tytgat, J.; Zega, A.; Mašič, L.P. Discovery of K V 1.3 ion channel inhibitors: Medicinal chemistry approaches and challenges. Med. Res. Rev. 2021, 41, 2423–2473. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.Z.; Cao, Z.J.; Li, W.X.; Sabatier, J.M.; Wu, Y.L. Treating autoimmune disorders with venom-derived peptides. Expert Opin. Biol. Ther. 2017, 17, 1065–1075. [Google Scholar] [CrossRef]
- Mao, X.; Cao, Z.; Yin, S.; Ma, Y.; Wu, Y.; Li, W. Cloning and characterization of BmK86, a novel K+ -channel blocker from scorpion venom. Biochem. Biophys. Res. Commun. 2007, 360, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Lee, A.; Campbell, E.; Mackinnon, R. Structure of a pore-blocking toxin in complex with a eukaryotic voltage-dependent K(+) channel. eLife 2013, 2, e00594. [Google Scholar] [CrossRef]
- Zhao, Y.H.; Chen, Z.Y.; Cao, Z.J.; Li, W.X.; Wu, Y. Diverse Structural Features of Potassium Channels Characterized by Scorpion Toxins as Molecular Probes. Molecules 2019, 24, 2045. [Google Scholar] [CrossRef] [Green Version]
- Gilquin, B.; Braud, S.; Eriksson, M.A.L.; Roux, B.; Bailey, T.D.; Priest, B.T.; Garcia, M.L.; Ménez, A.; Gasparini, S. A Variable Residue in the Pore of Kv1 Channels Is Critical for the High Affinity of Blockers from Sea Anemones and Scorpions. J. Biol. Chem. 2005, 280, 27093–27102. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.J.; Jiang, L.; Yi, H.; Han, S.; Yang, D.W.; Liu, M.L.; Liu, H.; Cao, Z.J.; Wu, Y.L.; Li, W.X. Different Residues in Channel Turret Determining the Selectivity of ADWX-1 Inhibitor Peptide between Kv1.1 and Kv1.3 Channels. J. Proteome Res. 2008, 7, 4890–4897. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Xie, Z.; Zhang, Q.; Li, Y.; Yang, F.; Chen, Z.; Li, W.; Cao, Z.; Wu, Y. Scorpion Potassium Channel-blocking Defensin Highlights a Functional Link with Neurotoxin. J. Biol. Chem. 2016, 291, 7097–7106. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Yin, S.; Yi, H.; Mouhat, S.; Qiu, S.; Cao, Z.; Sabatier, J.-M.; Wu, Y.; Li, W. Protein−Protein Recognition Control by Modulating Electrostatic Interactions. J. Proteome Res. 2010, 9, 3118–3125. [Google Scholar] [CrossRef]
- Chen, Z.; Hu, Y.; Hong, J.; Hu, J.; Yang, W.; Xiang, F.; Yang, F.; Xie, Z.; Cao, Z.; Li, W.; et al. Toxin acidic residue evolutionary function-guided design of de novo peptide drugs for the immunotherapeutic target, the Kv1.3 channel. Sci. Rep. 2015, 5, 9881. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Z.; Chen, Z.; Cao, Z.; Li, W.; Wu, Y. Scorpion Toxin-potassium Channel Interaction Law and its Applications. Venoms Toxins 2021, 1, 15–26. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Y.; Liu, H.; Yu, W.; Yang, F.; Li, W.; Cao, Z.; Wu, Y. Mouse β-Defensin 3, A Defensin Inhibitor of Both Its Endogenous and Exogenous Potassium Channels. Molecules 2018, 23, 1489. [Google Scholar] [CrossRef] [Green Version]
- Pena, S.D.J.; Coimbra, R.L.M. Ataxia and myoclonic epilepsy due to a heterozygous new mutation in KCNA2: Proposal for a new channelopathy. Clin. Genet. 2014, 87, e1–e3. [Google Scholar] [CrossRef]
- Helbig, K.L.; Hedrich, U.; Shinde, D.N.; Krey, I.; Teichmann, A.; Hentschel, J.; Schubert, J.; Chamberlin, A.C.; Huether, R.; Lu, H.; et al. A recurrent mutation in KCNA2 as a novel cause of hereditary spastic paraplegia and ataxia. Ann. Neurol. 2016, 80, 638–642. [Google Scholar] [CrossRef] [Green Version]
- Hundallah, K.; Alenizi, A.; Alhashem, A.; Tabarki, B. Severe early-onset epileptic encephalopathy due to mutations in the KCNA2 gene: Expansion of the genotypic and phenotypic spectrum. Eur. J. Paediatr. Neurol. 2016, 20, 657–660. [Google Scholar] [CrossRef]
- Wang, X.; Umetsu, Y.; Gao, B.; Ohki, S.; Zhu, S. Mesomartoxin, a new Kv1.2-selective scorpion toxin interacting with the channel selectivity filter. Biochem. Pharmacol. 2015, 93, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Liu, S.; Zhang, Y.; Qin, C.; Xu, L.; Li, W.; Cao, Z.; Li, W.; Wu, Y. Expression of recombinant α-toxin BmKM9 from scorpion Buthus martensii Karsch and its functional characterization on sodium channels. Peptides 2018, 99, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Hu, Y.; Yi, H.; Yin, S.; Han, S.; Hu, J.; Chen, Z.; Yang, W.; Cao, Z.; De Waard, M.; et al. Two Conserved Arginine Residues from the SK3 Potassium Channel Outer Vestibule Control Selectivity of Recognition by Scorpion Toxins. J. Biol. Chem. 2013, 288, 12544–12553. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Yi, H.; Yin, S.-J.; Chen, Z.-Y.; Liu, H.; Cao, Z.-J.; Wu, Y.-L.; Li, W.-X. Structural Basis of a Potent Peptide Inhibitor Designed for Kv1.3 Channel, a Therapeutic Target of Autoimmune Disease. J. Biol. Chem. 2008, 283, 19058–19065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Li, T.; Xu, L.; Yu, C.; Cao, Z.; Li, W.; Wu, Y. Kv1.3 potassium channel-blocking toxin Ctri9577, novel gating modifier of Kv4.3 potassium channel from the scorpion toxin family. Biochem. Biophys. Res. Commun. 2014, 444, 406–410. [Google Scholar] [CrossRef]
- Hu, Y.-T.; Hu, J.; Li, T.; Wei, J.-J.; Feng, J.; Du, Y.-M.; Cao, Z.-J.; Li, W.-X.; Wu, Y.-L. Open conformation of hERG channel turrets revealed by a specific scorpion toxin BmKKx2. Cell Biosci. 2014, 4, 18. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, C.; Yang, X.; Zuo, Z.; Yang, L.; Yang, F.; Cao, Z.; Chen, Z.; Wu, Y. BmK86-P1, a New Degradation Peptide with Desirable Thermostability and Kv1.2 Channel-Specific Activity from Traditional Chinese Scorpion Medicinal Material. Toxins 2021, 13, 610. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090610
Qin C, Yang X, Zuo Z, Yang L, Yang F, Cao Z, Chen Z, Wu Y. BmK86-P1, a New Degradation Peptide with Desirable Thermostability and Kv1.2 Channel-Specific Activity from Traditional Chinese Scorpion Medicinal Material. Toxins. 2021; 13(9):610. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090610
Chicago/Turabian StyleQin, Chenhu, Xuhua Yang, Zheng Zuo, Liuting Yang, Fan Yang, Zhijian Cao, Zongyun Chen, and Yingliang Wu. 2021. "BmK86-P1, a New Degradation Peptide with Desirable Thermostability and Kv1.2 Channel-Specific Activity from Traditional Chinese Scorpion Medicinal Material" Toxins 13, no. 9: 610. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090610