Venom Immunotherapy: From Proteins to Product to Patient Protection

 , ,
, ,
Abstract
:1. Introduction
- The route of administration of VIT is subcutaneous (s.c.) only, whereas in AIT aeroallergens can be administered s.c. but also sublingually.
- Standard VIT involves a 100 µg bee or wasp maintenance dosage, whereas uniform dosing recommendations do not exist for AIT (i.e., inhalant allergens) between comparable products.
- Aqueous and depot adjuvant formulations of VIT are marketed throughout Europe, whereas aqueous extracts for s.c. AIT with inhalant allergens are rarely used in Europe.
- Contraindications against VIT differ from those of AIT against inhalant allergens; consider the life-threatening nature of HVA [20].
- Success rates of VIT are substantially higher compared to AIT with inhalant allergens. However, this involves longer treatment courses of VIT (5 years standard VIT versus 3 years standard AIT with inhalant allergens) and high-dose VIT.
- Measuring success rates in VIT and AIT with inhalant allergens (e.g., primary outcome parameters in controlled clinical trials) differs substantially.
- About 30% of patients treated for HVA require life-long VIT. This group includes patients suffering from co-morbidities such as mastocytosis, patients with severe initial systemic sting reactions, patients who have had systemic adverse events during VIT, but also individuals with high risks of future bee and/or wasp stings, such as beekeepers or workers in confectioneries [21,22,23,24,25].
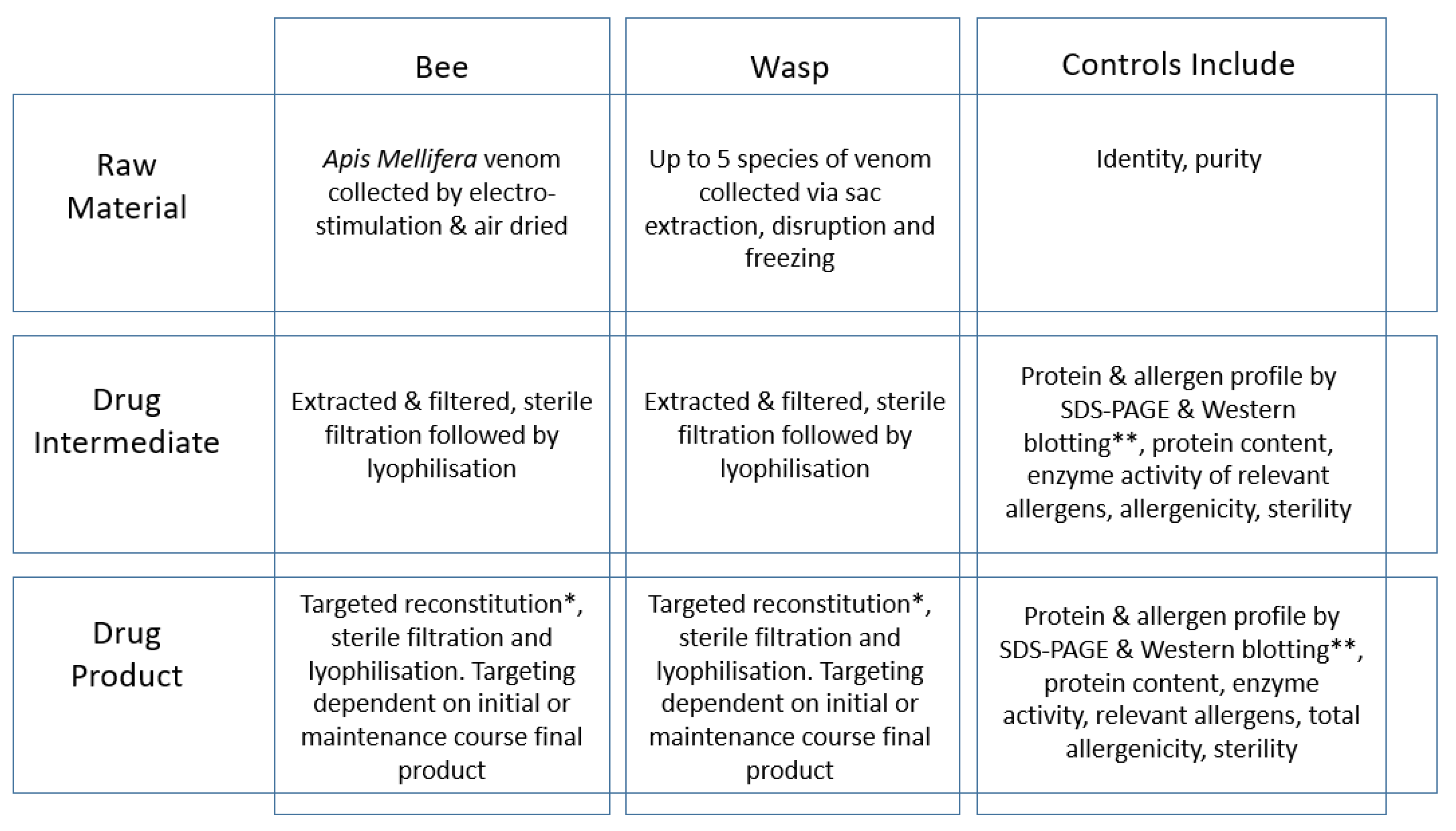
2. Production of Venom Extracts
3. The Molecular Story
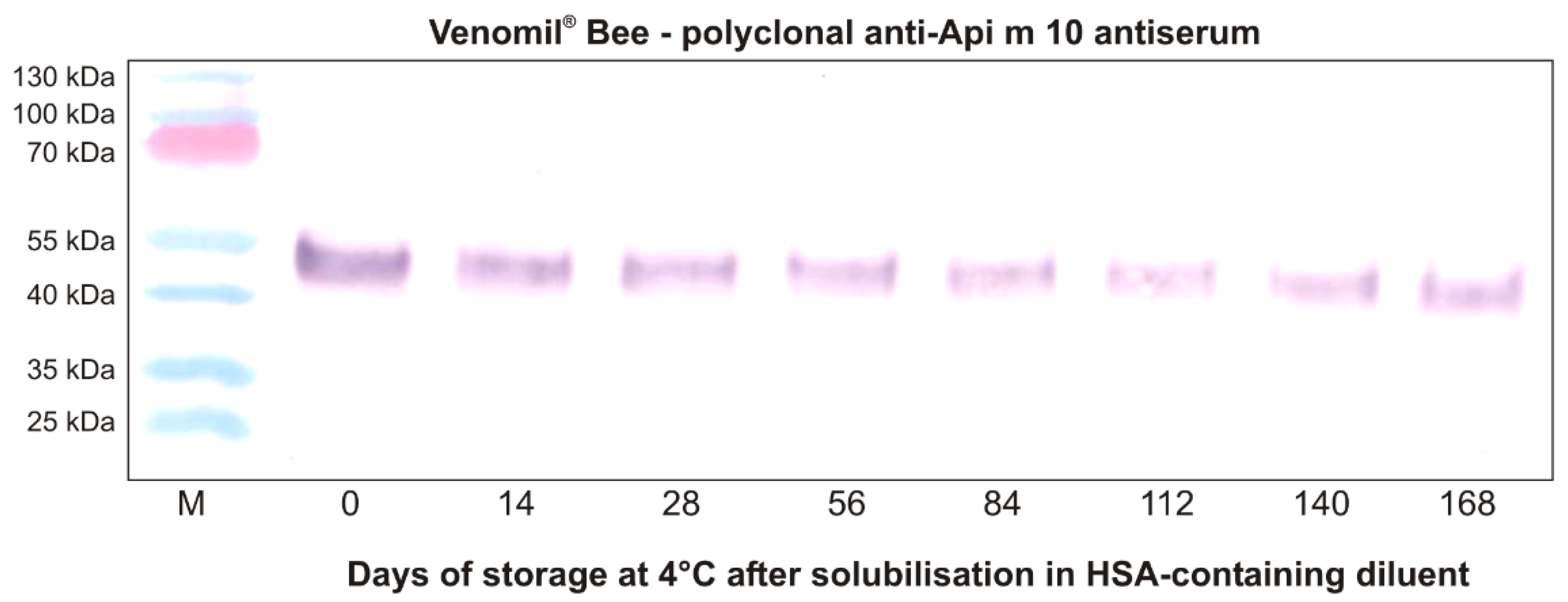
Api m 10 Stability in Venomil® and Its Potential Role for Therapeutic Success
4. Clinical Experience with Venomil®
4.1. Safety
4.2. Efficacy
5. The Choice of Depot for Long Term VIT: Considerations
6. Conclusions and Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blank, S.; Pehlivanli, S.; Methe, H.; Schmidt-Weber, C.B.; Biedermann, T.; Horny, H.P.; Kristensen, T.; Amar, Y.; Köberle, M.; Brockow, K.; et al. Fatal anaphylaxis following a hornet sting in a yellow jacket venom-sensitized patient with undetected monoclonal mast cell activation syndrome and without previous history of a systemic sting reaction. J. Allergy Clin. Immunol. Pract. 2020, 8, 401–403.e2. [Google Scholar] [CrossRef] [PubMed]
- Koschel, D. Impaired quality of life in patients with insect venom allergy. Allergo J. Int. 2017, 26, 88–92. [Google Scholar] [CrossRef]
- Manmohan, M.; Müller, S.; Myriam Rauber, M.; Koberne, F.; Reisch, H.; Koster, J.; Böhm, R.; Messelken, M.; Fischer, M.; Jakob, T. Current state of follow-up care for patients with Hymenoptera venom anaphylaxis in southwest Germany: Major impact of early information. Allergo J. Int. 2018, 27, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antolín-Amérigo, D.; Ruiz-León, B.; Vega-Castro, A.; de la Hoz Caballer, B. Natural history of systemic reactions and risk factors in children and adults with Hymenoptera venom allergy. Allergo J. 2020, 29, 26–33. [Google Scholar] [CrossRef]
- Blank, S.; Haemmerle, S.; Jaeger, T.; Russkamp, D.; Ring, J.; Schmidt-Weber, C.B.; Ollert, M. Prevalence of Hymenoptera venom allergy and sensitization in the population-representative German KORA cohort. Allergo J. Int. 2019, 28, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Schiener, M.; Graessel, A.; Ollert, M.; Schmidt-Weber, C.B.; Blank, S. Allergen-specific immunotherapy of Hymenoptera venom allergy—Also a matter of diagnosis. Hum. Vaccines Immunother. 2017, 13, 2467–2481. [Google Scholar] [CrossRef] [Green Version]
- Klimek, L.; Brehler, R.; Hamelmann, E.; Kopp, M.; Ring, J.; Treudler, R.; Jakob, T.; Worm, M.; Pfaar, O. Evolution of subcutaneous allergen immunotherapy (part 1): From first developments to mechanism-driven therapy concepts. Allergo J. Int. 2019, 28, 78–95. [Google Scholar] [CrossRef] [Green Version]
- Klimek, L.; Brehler, R.; Hamelmann, E.; Kopp, M.; Ring, J.; Treudler, R.; Jakob, T.; Worm, M.; Pfaar, O. Development of subcutaneous allergen immunotherapy (part 2): Preventive aspects and innovations. Allergo J. Int. 2019, 28, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Blank, S.; Bazon, M.L.; Grosch, J.; Schmidt-Weber, C.B.; Brochetto-Braga, M.R.; Bilo, M.B.; Jakob, T. Antigen 5 Allergens of Hymenoptera Venoms and Their Role in Diagnosis and Therapy of Venom Allergy. Curr. Allergy Asthma Rep. 2020, 20, 58. [Google Scholar] [CrossRef]
- Brehler, R. Insects and arachnids as elicitors of toxic and allergic reactions in Germany. Allergo J. Int. 2017, 26, 129–136. [Google Scholar] [CrossRef]
- Mauss, V.; Ruëff, F. Identification of bee and wasp taxa relevant in systemic allergic reactions to Hymenoptera stings in Central Europe. Allergo J. Int. 2017, 26, 81–87. [Google Scholar] [CrossRef]
- Blank, S.; Seismann, H.; Michel, Y.; McIntyre, M.; Cifuentes, L.; Braren, I.; Grunwald, T.; Darsow, U.; Ring, J.; Bredehorst, R.; et al. Api m 10, a genuine A. mellifera venom allergen, is clinically relevant but underrepresented in therapeutic extracts. Allergy 2011, 66, 1322–1329. [Google Scholar] [CrossRef]
- Frick, M.; Fischer, J.; Helbling, A.; Rueff, F.; Wieczorek, D.; Ollert, M.; Pfutzner, W.; Muller, S.; Huss-Marp, J.; Dorn, B.; et al. Predominant Api m 10 sensitization as risk factor for treatment failure in honey bee venom immunotherapy. J. Allergy Clin. Immunol. 2016, 138, 1663–1671.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakob, T.; Rauber, M.M.; Perez-Riverol, A.; Spillner, E.; Blank, S. The Honeybee Venom Major Allergen Api m 10 (Icarapin) and Its Role in Diagnostics and Treatment of Hymenoptera Venom Allergy. Curr. Allergy Asthma Rep. 2020, 20, 48. [Google Scholar] [CrossRef] [PubMed]
- Pereira Santos, M.C.; Lourenco, T.; Pereira Barbosa, M.; Branco Ferreira, M. Evolution of Api m10 Specific IgE and IgG4 After One Year of Bee Venom Immunotherapy. Eur. Ann. Allergy Clin. Immunol. 2020, 52, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Kohler, J.; Blank, S.; Muller, S.; Bantleon, F.; Frick, M.; Huss-Marp, J.; Lidholm, J.; Spillner, E.; Jakob, T. Component resolution reveals additional major allergens in patients with honeybee venom allergy. J. Allergy Clin. Immunol. 2014, 133, 1383–1389.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arzt, L.; Bokanovic, D.; Schrautzer, C.; Schwarz, I.; Laipold, K.; Aberer, W.; Sturm, G.J. Questionable diagnostic benefit of the commercially available panel of bee venom components. Allergy 2017, 72, 1419–1422. [Google Scholar] [CrossRef]
- Bilò, M.B.; Ollert, M.; Blank, S. The role of component-resolved diagnosis in Hymenoptera venom allergy. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 614–622. [Google Scholar] [CrossRef]
- Blank, S.; Etzold, S.; Darsow, U.; Schiener, M.; Eberlein, B.; Russkamp, D.; Wolf, S.; Graessel, A.; Biedermann, T.; Ollert, M.; et al. Component-resolved evaluation of the content of major allergens in therapeutic extracts for specific immunotherapy of honeybee venom allergy. Hum. Vaccines Immunother. 2017, 13, 2482–2489. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.; Ruëff, F. Dealing with absolute and relative contraindications to specific immunotherapy using Hymenoptera venoms. Allergo J. Int. 2017, 26, 122–128. [Google Scholar] [CrossRef]
- Sturm, G.J.; Varga, E.M.; Roberts, G.; Mosbech, H.; Bilò, M.B.; Akdis, C.A.; Antolín-Amérigo, D.; Cichocka-Jarosz, E.; Gawlik, R.; Jakob, T.; et al. EAACI guidelines on allergen immunotherapy: Hymenoptera venom allergy. Allergy 2018, 73, 744–764. [Google Scholar] [CrossRef] [Green Version]
- González-de-Olano, D.; Álvarez-Twose, I.; Vega-Castro, A. Venom immunotherapy in patients with mastocytosis. Allergo J. Int. 2020, 29, 67–72. [Google Scholar] [CrossRef]
- Ruëff, F.; Mastnik, S.; Oppel, E.M. Mast cell diseases in patients with insect venom allergy: Implications for diagnosis and therapy. Allergo J. Int. 2017, 26, 137–145. [Google Scholar] [CrossRef]
- Bonadonna, P.; Boni, E.; Zanotti, R. Mastocytosis as a risk factor for insect venom allergy. Allergo J. Int. 2020, 29, 73–78. [Google Scholar] [CrossRef]
- Dinkel, G.; Pietschke, K.; Belge, K.; Yazdi, A.; Fischer, J. Bee venom allergy: A recent trend among amateur beekeepers. Allergo J. Int. 2017, 26, 119–121. [Google Scholar] [CrossRef]
- Plunkett, G.; Jacobson, R.S.; Golden, D.B.K. Hymenoptera venoms used to produce allergen extracts. Ann. Allergy Asthma Immunol. 2017, 118, 649–654. [Google Scholar] [CrossRef]
- Hoffman, D.R. Allergens in hymenoptera venom XIII: Isolation and purification ofprotein components from three species of vespid venoms. J. Allergy Clin. Immunol. 1985, 75, 599–605. [Google Scholar] [CrossRef]
- Hoffman, D.R. Allergens in Hymenoptera venom XXV: The amino acid sequences of antigen 5 molecules and the structural basis of antigen cross-reactivity. J. Allergy Clin. Immunol. 1993, 92, 707–716. [Google Scholar] [CrossRef]
- Wicher, K.; Reisman, R.E.; Wypych, J.; Elliott, W.; Steger, R.; Mathews, R.S.; Arbesman, C.E. Comparison of the venom immunogenicity of various species of yellow jackets (genus Vespula). J. Allergy Clin. Immunol. 1980, 66, 244–249. [Google Scholar] [CrossRef]
- Jakob, T.; Rafei-Shamsabadi, D.; Spillner, E.; Muller, S. Diagnostics in Hymenoptera venom allergy: Current concepts and developments with special focus on molecular allergy diagnostics. Allergo J. Int. 2017, 26, 93–105. [Google Scholar] [CrossRef]
- Blank, S.; Bilò, M.B.; Grosch, J.; Schmidt-Weber, C.B.; Ollert, M.; Jakob, T. Marker allergens in Hymenoptera venom allergy—Characteristics and potential use in precision medicine. Allergo J. Int. 2021, 30, 26–38. [Google Scholar] [CrossRef]
- Ollert, M.; Blank, S. Anaphylaxis to insect venom allergens: Role of molecular diagnostics. Curr. Allergy Asthma Rep. 2015, 15, 26. [Google Scholar] [CrossRef] [Green Version]
- Hemmer, W. Cross-reactivity to honeybee and wasp venom. Hautarzt 2008, 59, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Matuszewska, E.; Matysiak, J.; Breborowicz, A.; Olejniczak, K.; Kycler, Z.; Kokot, Z.J.; Matysiak, J. Proteomic features characterization of Hymenoptera venom allergy. Allergy Asthma Clin. Immunol. 2019, 15, 77. [Google Scholar] [CrossRef]
- Ewan, P.W. Venom allergy. BMJ 1998, 316, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Peiren, N.; de Graaf, D.C.; Brunain, M.; Bridts, C.H.; Ebo, D.G.; Stevens, W.J.; Jacobs, F.J. Molecular cloning and expression of icarapin, a novel IgE-binding bee venom protein. FEBS Lett. 2006, 580, 4895–4899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, M.; Fierlbeck, G.; King, T.; Valent, P.; Bühring, H.J. Individual hymenoptera venom compounds induce upregulation of the basophil activation marker ectonucleotide pyrophosphatase/phosphodiesterase 3 (CD203c) in sensitized patients. Int. Arch. Allergy Immunol. 2002, 129, 160–168. [Google Scholar] [CrossRef]
- Grunwald, T.; Bockisch, B.; Spillner, E.; Ring, J.; Bredehorst, R.; Ollert, M.W. Molecular cloning and expression in insect cells of honeybee venom allergen acid phosphatase (Api m 3). J. Allergy Clin. Immunol. 2006, 117, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Spillner, E.; Blank, S.; Jakob, T. Hymenoptera allergens: From venom to “venome”. Front. Immunol. 2014, 5, 77. [Google Scholar] [CrossRef] [Green Version]
- Spiric, J.; Paulus, K.; Schwaben, L.; Vieths, S.; Junker, A.; Mahler, V.; Reuter, A. Complete compositional analysis of honeybee venom therapeutic products by mass spectrometry. Confirmation of all known Api m allergens in one blow. In EAACI 2018 Poster 1680; Paul Ehrlich Institute: Langen, Germany, 2018. [Google Scholar]
- Paulus, K.E.; Spiric, J.; Junker, A.; Schwaben, L.; Lidholm, J.; Vieths, S.; Mahler, V. Api m 10 can be detected qualitatively by mass spectrometry in all products for allergen immunotherapy for honeybee venom allergy. In EAACI 2018 Poster 1677; Paul Ehrlich Institute: Langen, Germany; Thermo Fisher Scientific: Uppsala, Sweden, 2018. [Google Scholar]
- Van Vaerenbergh, M.; De Smet, L.; Rafei-Shamsabadi, D.; Blank, S.; Spillner, E.; Ebo, D.G.; Devreese, B.; Jakob, T.; de Graaf, D.C. IgE recognition of chimeric isoforms of the honeybee (Apis mellifera) venom allergen Api m 10 evaluated by protein array technology. Mol. Immunol. 2015, 63, 449–455. [Google Scholar] [CrossRef]
- Rauber, M.M.; Roßbach, A.; Jung, A.; Müller, S.; Möbs, C.; Pfützner, W.; Miehe, M.; Spillner, E.; Jakob, T. The honey bee venom allergen Api m 10 displays one major IgE epitope, Api m 10(160-174). Allergy 2020, 75, 1756–1759. [Google Scholar] [CrossRef] [Green Version]
- Nataf, P.; Guinnepain, M.T.; Herman, D. Rush venom immunotherapy: A 3-day programme for hymenoptera sting allergy. Clin. Exp. Allergy 1984, 14, 269–275. [Google Scholar] [CrossRef]
- Birnbaum, J.; Charpin, D.; Vervloet, D. Rapid Hymenoptera venom immunotherapy: Comparative safety of three protocols. Clin. Exp. Allergy 1993, 23, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Gleich, G.; Yunginger, J.; Lichtenstein, L.W.; Valentine, M.D.; Grant, A.; Stier, R.A. Therapy for Hypersensitivity to Hymenoptera Stings. In Report to US Food & Drug Administration under BB-IND 1292; Data on File; June 1982.
- Becker, M.; Steiß, J.O. Poster: Safety of modified ultra-rush venom immunotherapy with lyophilized extracts in children—A retrospective and prospective analysis. In Allergy; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Ruëff, F.; Przybilla, B. Venom immunotherapy: Adverse reactions and treatment failure. Curr. Opin. Allergy Clin. Immunol. 2004, 4, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-H.; Landeck, L.; Stefaniak, R.; Zuberbier, T.; Worm, M. Verträglichkeit der spezifischen Wespengift-Immuntherapie nach einem Rush- oder Ultra-Rush-Protokoll. Allergo J. 2005, 14, 482–486. [Google Scholar]
- Döring, H.F.; Tüttenberg, H.W. Insektengifthyposensibilisierung beim niedergelassenen Dermatologen. Der. Dermatol. 1994, 42, 1–4. [Google Scholar]
- Jung, A.; Schill, W.-B. Verlaufsparameter zur HyposensibiIisierung bei Hymenopterengiftallergie. Z. Haulkr. 2002, 77, 241–244. [Google Scholar]
- Lohse, K.; Rabe, U. Poster—Evaluation of a „super-rush dose regimen” for yearly therapy control and boostering after completed insect venom immunotherapy with bee/wasp. In Proceedings of the World Allergy Congress, Munich, Germany, 26 June–1 July 2005. [Google Scholar]
- Munstedt, K.; Wrobel, D.; Kalder, M. Efficacy of venom immunotherapy in beekeepers. J. Investig. Allergol. Clin. Immunol. 2010, 20, 58–62. [Google Scholar] [PubMed]
- Lee, H.; Zuberbier, T.; Worm, M. CD203c-Expression auf basophilen Granulozyten im Verlauf der spezifischen Immuntherapie bei Wespengiftallergikern. <Poster Lee Basophilen Aktivierung Erfurt 08.pdf>. Allergie-Kongress 2008. Data on File. Erfurt 2008.
- Baenkler, H.W.; Meusser-Storm, S.; Eger, G. Continuous immunotherapy for hymenoptera venom allergy using six month intervals. Allergol. Immunopathol. 2005, 33, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, U.; Helbling, A.; Berchtold, E. Immunotherapy with honeybee venom and yellow jacket venom is different regarding efficacy and safety. J. Allergy Clin. Immunol. 1992, 89, 529–535. [Google Scholar] [CrossRef]
- Ruëff, F.; Vos, B.; Oude Elberink, J.; Bender, A.; Chatelain, R.; Dugas-Breit, S.; Horny, H.P.; Küchenhoff, H.; Linhardt, A.; Mastnik, S.; et al. Predictors of clinical effectiveness of Hymenoptera venom immunotherapy. Clin. Exp. Allergy 2014, 44, 736–746. [Google Scholar] [CrossRef]
- Frew, A.J.; Pfaar, O. Placebo effects in allergen immunotherapy: An experts’ opinion. Allergo J. Int. 2018, 27, 162–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roesch, A.; Boerzsoenyi, J.; Babilas, P.; Landthaler, M.; Szeimies, R.M. Outcome survey of insect venom allergic patients with venom immunotherapy in a rural population. J. Dtsch. Dermatol. Ges. 2008, 6, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Stoevesandt, J.; Trautmann, A. Lessons from times of shortage: Interchangeability of venom preparations and dosing protocols. Allergy 2019, 74, 1392–1395. [Google Scholar] [CrossRef] [PubMed]
- Heddle, R.; Smith, A.; Woodman, R.; Hissaria, P.; Petrovsky, N. Randomized controlled trial demonstrating the benefits of delta inulin adjuvanted immunotherapy in patients with bee venom allergy. J. Allergy Clin. Immunol. 2019, 144, 504–513.e16. [Google Scholar] [CrossRef] [PubMed]
- Mosbech, H.; Müller, U. Side-effects of insect venom immunotherapy: Results from an EAACI multicenter study. European Academy of Allergology and Clinical Immunology. Allergy 2000, 55, 1005–1010. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Jarolim, E.; Bachmann, M.; Bonini, S.; Jacobsen, L.; Jutel, M.; Klimek, L.; Mahler, V.; Mosges, R.; Moingeon, P.; RE, O.H.; et al. State-of-the-art in marketed adjuvants and formulations in Allergen Immunotherapy: A position paper of the European Academy of Allergy and Clinical Immunology (EAACI). Allergy 2020, 75, 746–760. [Google Scholar] [CrossRef]
- McDougall, S.A.; Heath, M.D.; Kramer, M.F.; Skinner, M.A. Analysis of aluminium in rat following administration of allergen immunotherapy using either aluminium or microcrystalline-tyrosine-based adjuvants. Bioanalysis 2016, 8, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Kramer, M.F.; Heath, M.D. Aluminium in allergen-specific subcutaneous immunotherapy--a German perspective. Vaccine 2014, 32, 4140–4148. [Google Scholar] [CrossRef] [Green Version]
- PEI. Available online: https://www.pei.de/DE/newsroom/veroffentlichungen-arzneimittel/sicherheitsinformationen-human/2014/ablage2014/2014-01-21-sicherheitsbewertung-von-aluminium-in-therapieallergenen.html (accessed on 10 May 2021).
- Weisser, K.; Stubler, S.; Matheis, W.; Huisinga, W. Towards toxicokinetic modelling of aluminium exposure from adjuvants in medicinal products. Regul. Toxicol. Pharmacol. 2017, 88, 310–321. [Google Scholar] [CrossRef]
- Ruëff, F.; Wolf, H.; Schnitker, J.; Ring, J.; Przybilla, B. Specific immunotherapy in honeybee venom allergy: A comparative study using aqueous and aluminium hydroxide adsorbed preparations. Allergy 2004, 59, 589–595. [Google Scholar] [CrossRef]
- Pfaar, O.; Bachert, C.; Bufe, A.; Buhl, R.; Ebner, C.; Eng, P.; Friedrichs, F.; Fuchs, T.; Hamelmann, E.; Hartwig-Bade, D.; et al. Guideline on allergen-specific immunotherapy in IgE-mediated allergic diseases: S2k Guideline of the German Society for Allergology and Clinical Immunology (DGAKI), the Society for Pediatric Allergy and Environmental Medicine (GPA), the Medical Association of German Allergologists (AeDA), the Austrian Society for Allergy and Immunology (ÖGAI), the Swiss Society for Allergy and Immunology (SGAI), the German Society of Dermatology (DDG), the German Society of Oto- Rhino-Laryngology, Head and Neck Surgery (DGHNO-KHC), the German Society of Pediatrics and Adolescent Medicine (DGKJ), the Society for Pediatric Pneumology (GPP), the German Respiratory Society (DGP), the German Association of ENT Surgeons (BV-HNO), the Professional Federation of Paediatricians and Youth Doctors (BVKJ), the Federal Association of Pulmonologists (BDP) and the German Dermatologists Association (BVDD). Allergo J. Int. 2014, 23, 282–319. [Google Scholar] [CrossRef]
- Heath, M.D.; Mohsen, M.O.; de Kam, P.J.; Carreno Velazquez, T.L.; Hewings, S.J.; Kramer, M.F.; Kundig, T.M.; Bachmann, M.F.; Skinner, M.A. Shaping Modern Vaccines: Adjuvant Systems Using MicroCrystalline Tyrosine (MCT). Front. Immunol. 2020, 11, 594911. [Google Scholar] [CrossRef]
- Leuthard, D.S.; Duda, A.; Freiberger, S.N.; Weiss, S.; Dommann, I.; Fenini, G.; Contassot, E.; Kramer, M.F.; Skinner, M.A.; Kundig, T.M.; et al. Microcrystalline Tyrosine and Aluminum as Adjuvants in Allergen-Specific Immunotherapy Protect from IgE-Mediated Reactivity in Mouse Models and Act Independently of Inflammasome and TLR Signaling. J. Immunol. 2018, 200, 3151–3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shardlow, E.; Exley, C. The size of micro-crystalline tyrosine (MCT®) influences its recognition and uptake by THP-1 macrophages in vitro. RSC Adv. 2019, 9, 24505–24518. [Google Scholar] [CrossRef] [Green Version]
- Mohsen, M.O.; Heath, M.D.; Cabral-Miranda, G.; Lipp, C.; Zeltins, A.; Sande, M.; Stein, J.V.; Riether, C.; Roesti, E.; Zha, L.; et al. Vaccination with nanoparticles combined with micro-adjuvants protects against cancer. J. Immunother. Cancer 2019, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, L.; Duschl, A.; Himly, M. Nanotechnology-Based Vaccines for Allergen-Specific Immunotherapy: Potentials and Challenges of Conventional and Novel Adjuvants under Research. Vaccines 2020, 8, 237. [Google Scholar] [CrossRef]
- Mahler, V.; Bonertz, A.; Ruoff, C.; Hartenstein, D.; Mentzer, D.; Kaul, S.; Vieths, S. What we learned from TAO—10 years of German therapy allergen ordinance. Allergo J. Int. 2019, 28, 330–337. [Google Scholar] [CrossRef]
- Bilò, M.B.; Martini, M.; Corsi, A.; Tontini, C.; Antonicelli, L. Venom immunotherapy in Europe and the United States. Allergo J. Int. 2020, 29, 29–37. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Company | Venoms Covered under Marketing Authorization (MA) | Country in Which Market Authorization Is Held |
|---|---|---|
| Bayer | All Hymenoptera venom products | US |
| Stallergenes | Honeybee and Yellow jacket | France |
| Allergy Therapeutics * | All Hymenoptera venom products | Germany |
| Biochemical Name | Venom | Allergen Nomenclature | MW | Potential N-Glycosylation | Allergenicity |
|---|---|---|---|---|---|
| Phospholipases | |||||
| A2 | Honeybee | Api m 1 | 16 kDa | 1 | An in-vitro study using flow-cytometry analysis, showed an increase of basophil CD203c in response to Api m 1 in 9 of 13 patients with bee allergy. Specific IgE levels to Api m 1 increased in 4 patients allergic to bee venom (quantified by fluorescence immunoassay (UniCAP®); 5 patients tested) [37]. The sensitization rate to Api m 1 is reported to range from 57–97% in honeybee venom-allergic patients in different cohorts [31]. PDB structure: 1POC Uniprot: P00630 |
| A1B | Wasp | Ves v 1 | 34 kDa | 0 | In the same in-vitro study as above, 17 patients with allergy of wasp venom presented upregulation of CD203c basophil response when stimulated with purified Ves v 1 [37]. IgE sensitization to Ves v 1 varies from 39–66% as per different populations of wasp allergic patients and is reported to increase in wasp/honeybee double-sensitized patients [31]. UniProt: P49369 |
| Hyaluronidases | |||||
| Honeybee | Api m 2 | 44 kDa | 3 | An in-vitro study demonstrated that all 13 patients with bee allergy responded to purified Api m 2 by increasing levels of basophil activation marker CD203c. Specific IgE levels were increased in 5 patients that reacted with Api m 2 using RAST in 5 patients allergic to bee venom (39 patients tested) [37]. Sensitization rates range from 28–60% in different study populations (28–55 in honeybee venom-allergic patients and 45–60% in honeybee/wasp venom double-sensitized patients) [31]. PDB structure: 1FCQ UniProt: Q08169 | |
| Wasp | Ves v 2 | 38 kDa | 4 | Of 35 patients allergic to wasp venom, 26 patients presented upregulation of CD203c basophil response when stimulated with purified Ves v 2 [37]. Around 10–15% of patients with wasp allergy are estimated to have IgE antibodies against Ves v 2 and peptide-specific cross-reactivity with Api m 2 [31]. PDB structure: 2ATM Uniprot: P49370 & Q5D7H4 | |
| Acid phosphatases | Honeybee | Api m 3 | 43–49 kDa | 2 | Recombinant allergen Api m 3 showed immunoreactivity to specific IgE antibodies in pooled serum by Western blot and 37% in individual sera by ELISA in honeybee venom-sensitized patients [38]. The sensitization rate is reported to range from 28–63% in different studies of honeybee venom-allergic patients [31]. Uniprot: Q4TUB9 |
| Dipeptidyl peptidase IV | Honyebee | Api m 5 | 100 kDa | 6 | IgE reactivity to Api m 5 was detected in 58.3% of 144 honeybee venom allergy patients [16]. The sensitization rate is reported to range from 16–70% in different studies of honeybee venom-allergic patients [31]. Uniprot: B2D0J4 |
| Icarapin variant 2 | Honeybee | Api m 10 | 50–55 kDa | 2 | IgE reactivity presented in 50% of 84 honeybee venom-sensitized patients [12]. The sensitization rate varies from 35–75% in honeybee venom-allergic patients [31]. Uniprot: Q1HHN7 |
| Antigen 5 | Wasp | Ves v 5 | 23 kDa | 0 | Of 26 patients allergic to wasp venom and 8 patients allergic to bee and wasp venom (24 tested patients), 27 showed upregulation of CD203c expression in basophils in response to Ves v 5 [37]. The sensitization rate of Ves v 5 is reported to range from 82–98% in wasp venom-allergic patients [31]. PDB structure: 1QNX UniProt: Q05110 |
| PDB: Protein Data Bank; UniProt: Universal Protein Resource (comprehensive database of protein sequence). | |||||
| Publication | Species | Class | Patients | Implied ADR Rates | Implied Efficacy Rate | Notes | Reference |
|---|---|---|---|---|---|---|---|
| Döring et al., 1994 | Honeybee + Wasp | Retrospective NIS | 612 | 100% local ADRs 3.9% ADRs grade I+ 0.9% ADRs grade II+ | 99.3% protection (100% HB, 99.0% Wsp) | Analysis of 14 years of patient data from private practice | [50] |
| Baenkler et al., 2005 | Honeybee + Wasp | Non-controlled IIT | 176 | N/A local ADRs 14.2% systemic ADRs 1.7% ADRs grade II+ | 89.3% complete protection (71.4% HB, 88.7% Wsp, 100% HB + Wsp) 98.7% partial protection (100% HB, 98.1% Wsp, 100% HB + Wsp) | Venomil and Reless; Off-label continuation course using 6-month-intervals after month 9. | [55] |
| Jung et al., 2002 | Honeybee + Wasp | NIS | 50 | N/A local ADRs 12.0% systemic ADRs 0% ADRs grade II+ | 73.7% complete protection (75.0% HB, 73.3% Wsp) 100% partial protection | Evaluation of biomarkers (sIgE, skin tests) for treatment control | [51] |
| Lee et al., 2005 | Wasp | NIS | 50 | 100% local ADRs 10% systemic ADRs 2% ADRs grade II+ | N/A | Tolerability study of rush- and ultra-rush posologies | [49] |
| Lohse et al., 2005 | Honeybee + Wasp | Non-controlled IIT | 36 | N/A | 100% protection | Evaluation of a “super rush dose regimen” as treatment control and possible booster after 3–5 years of VIT | [52] |
| Münstedt et al., 2010 | Honeybee | Retrospective NIS | 43 | N/A | 97.7% complete protection 100% partial protection | Survey and clinical follow-up of VIT-treated beekeepers; 80% Venomil patients, 20% Reless; high chance of selection bias | [53] |
| Lee et al., 2008 | Wasp | Non-controlled IIT | 11 | N/A | 100% protection | Basophil activation test as treatment control; sting challenge after 1 year | [54] |
| Roesch et al., 2008 | Honeybee + Wasp | Retrospective NIS | 137 | N/A | 80% complete protection 87.7+% partial protection | Survey of VIT-treated patients | [59] |
| Stoevesandt et al., 2019 | Honeybee + Wasp | Retrospective NIS | 44 | N/A local ADRs 6.8% systemic ADRs | N/A | Tolerability of Venomil vs. ALK lyophilisiert/depot SQ | [60] |
| Becker et al., 2020 | Honeybee + Wasp | Retrospective NIS | 114 | N/A local ADRs 1.8% systemic ADRs | 87% partial protection (85.7% HB, 88.2% Wsp) | Analysis of 16 years of patient data from one university pediatrics department | [47] |
| Bayer Inc. 1982 | Honeybee + Wasp | Prospective Open-Label Clinical Trial | 114 | N/A local ADRs 30% systemic ADRs | 88.7% complete protection 98.3% partial protection | Licensing study for Safety and Efficacy in the US; sting challenge after reaching maintenance dose | [46] |
| Nataf et al., 1984 | Honeybee + Wasp | Prospective Open-Label Clinical Trial | 52 | N/A local ADRs N/A systemic ADRs 1.9% ADRs grade II+ | 100% protection | Evaluation of four-day rush initiation treatment | [44] |
| Birnbaum et al., 1993 | Honeybee + Wasp | Prospective Open-Label Clinical Trial | 284 | 100% local ADRs 12.0% systemic ADRs | N/A | Evaluation of three different rapid initiation treatments | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feindor, M.; Heath, M.D.; Hewings, S.J.; Carreno Velazquez, T.L.; Blank, S.; Grosch, J.; Jakob, T.; Schmid-Grendelmeier, P.; Klimek, L.; Golden, D.B.K.; et al. Venom Immunotherapy: From Proteins to Product to Patient Protection. Toxins 2021, 13, 616. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090616
Feindor M, Heath MD, Hewings SJ, Carreno Velazquez TL, Blank S, Grosch J, Jakob T, Schmid-Grendelmeier P, Klimek L, Golden DBK, et al. Venom Immunotherapy: From Proteins to Product to Patient Protection. Toxins. 2021; 13(9):616. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090616
Chicago/Turabian StyleFeindor, Martin, Matthew D. Heath, Simon J. Hewings, Thalia L. Carreno Velazquez, Simon Blank, Johannes Grosch, Thilo Jakob, Peter Schmid-Grendelmeier, Ludger Klimek, David B. K. Golden, and et al. 2021. "Venom Immunotherapy: From Proteins to Product to Patient Protection" Toxins 13, no. 9: 616. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090616