Variation in Rotenone and Deguelin Contents among Strains across Four Tephrosia Species and Their Activities against Aphids and Whiteflies

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
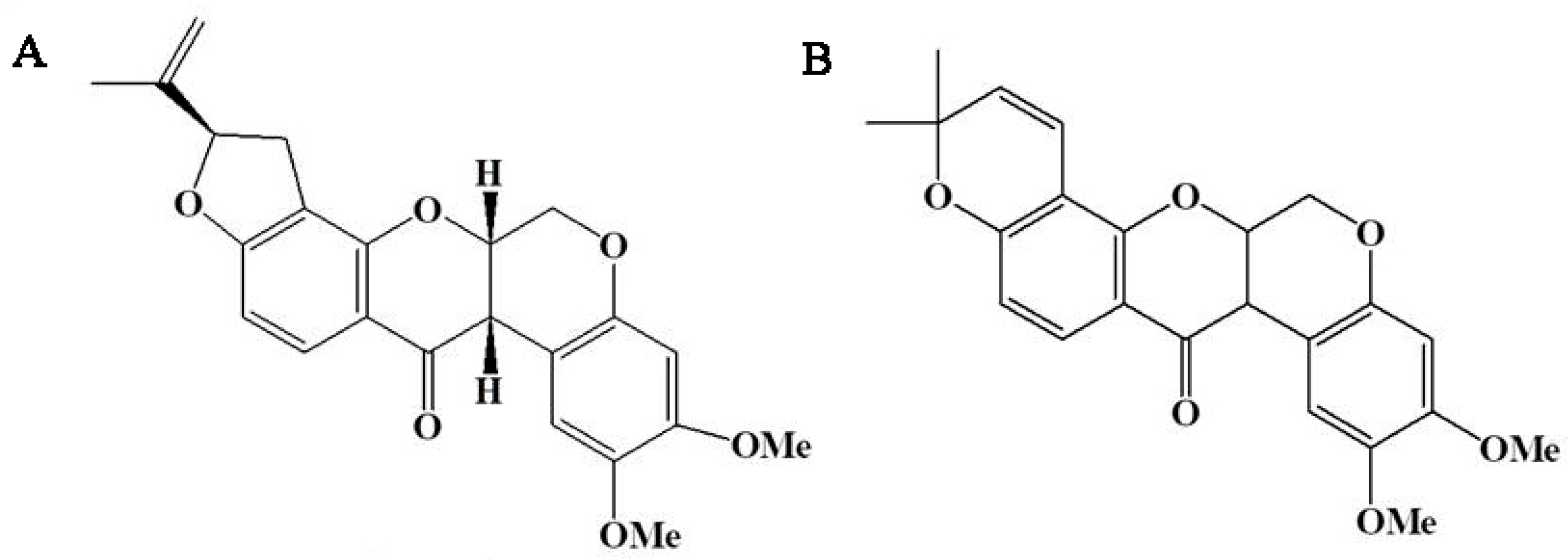
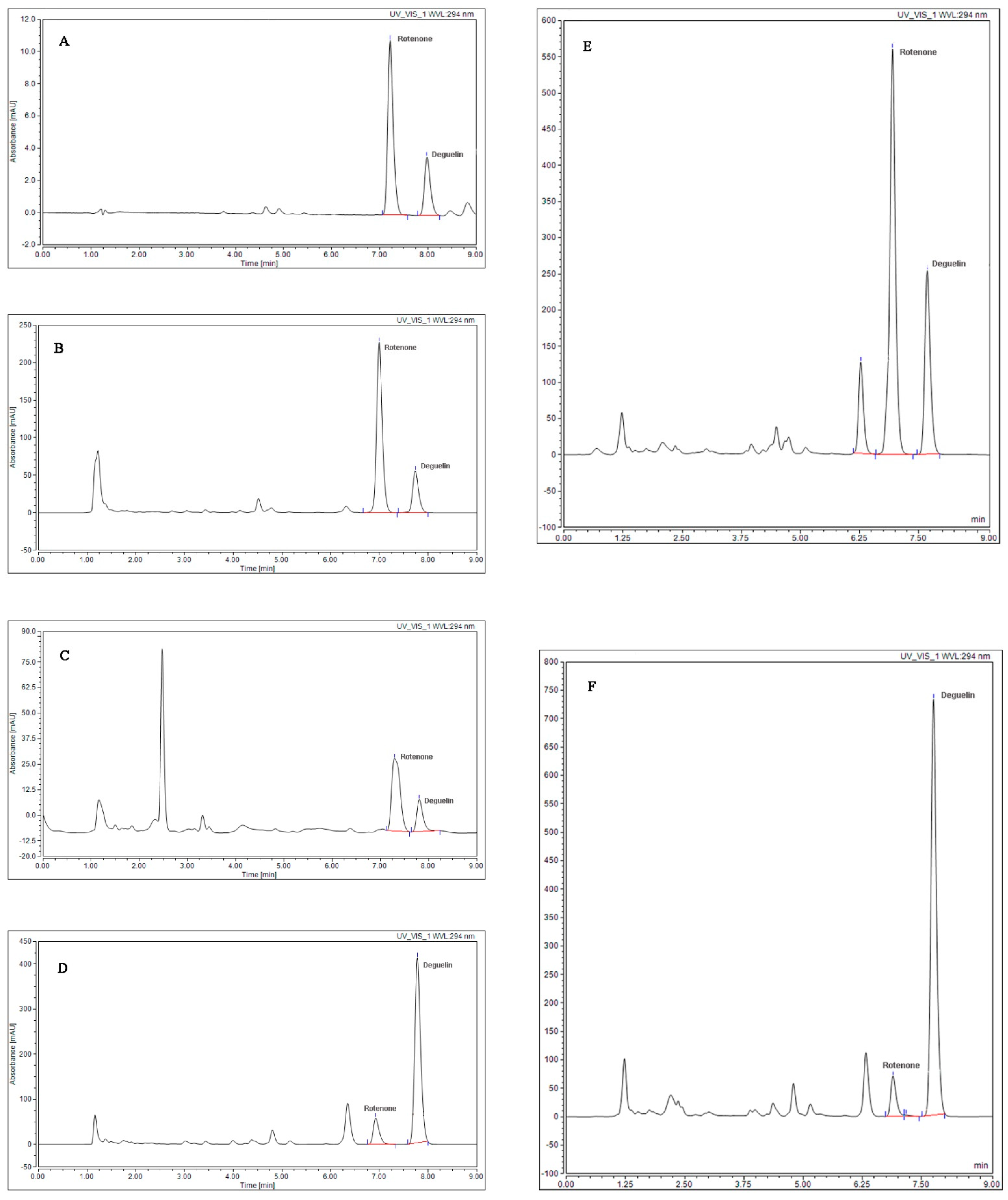
2.1. Rotenoid Contents in 13 Strains
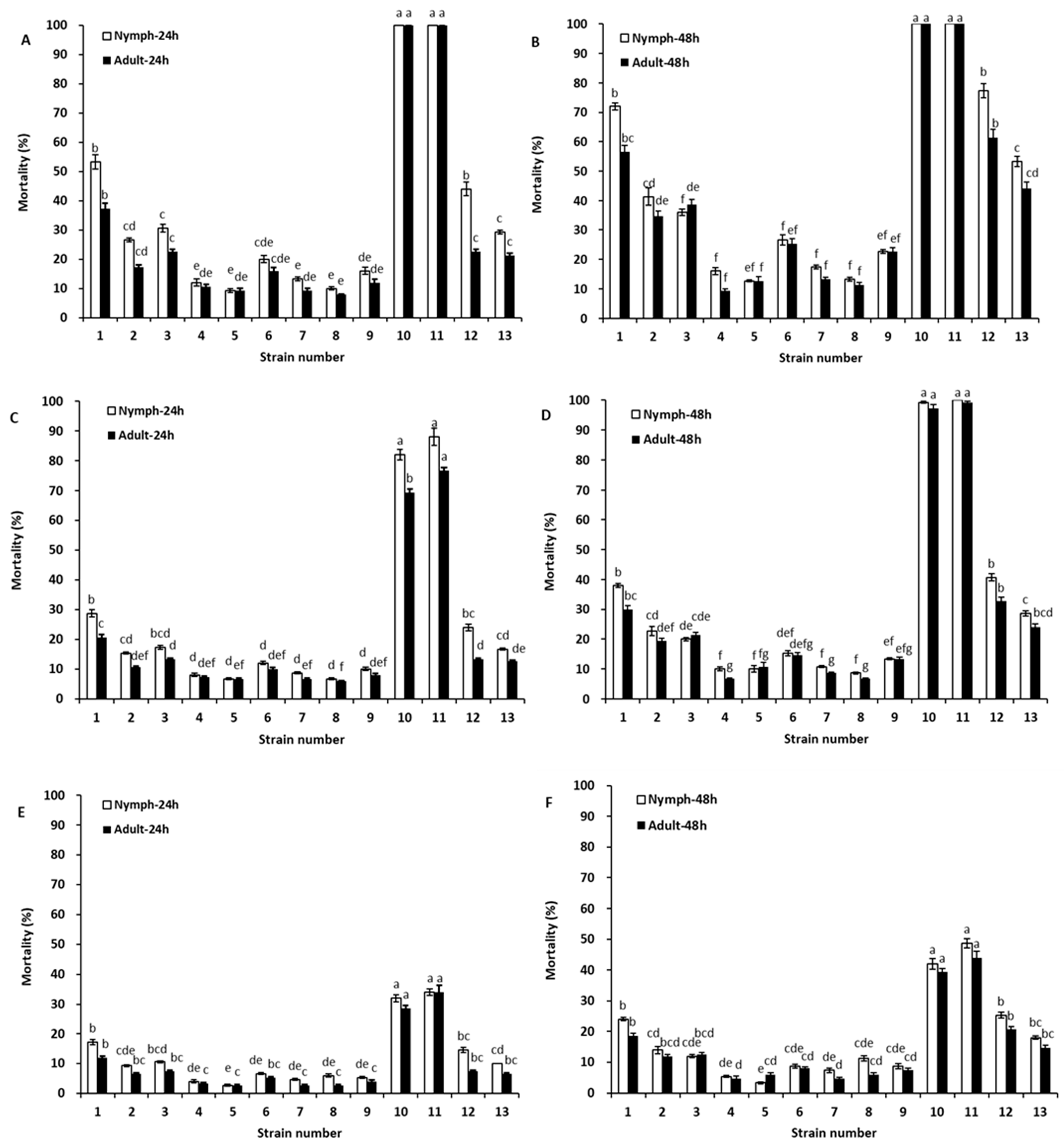
2.2. Toxicity of Rotenone and Deguelin Chemicals against Aphids and Whiteflies
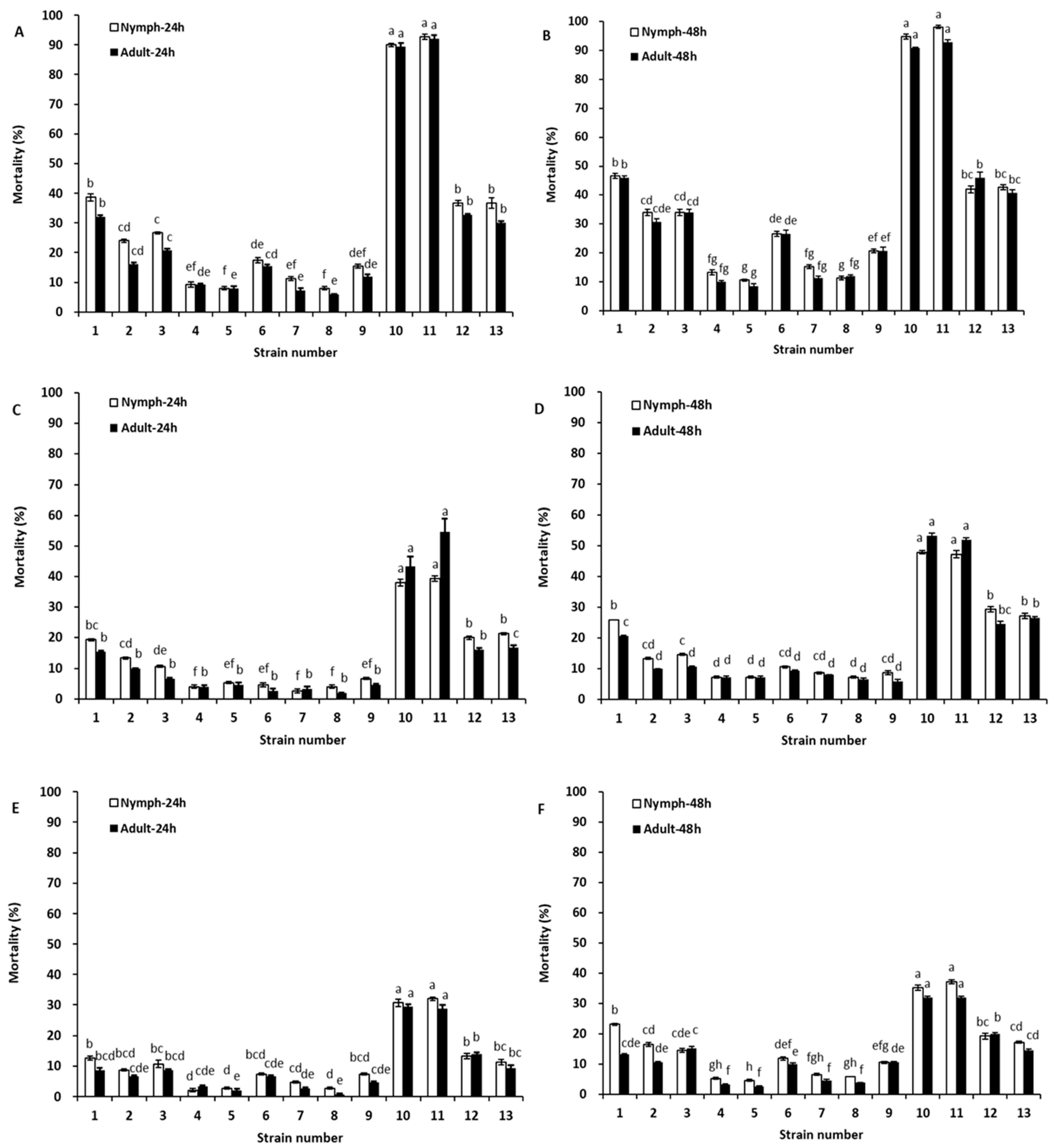
2.3. Toxicity of Tephrosia Leaf Extracts to Aphis gossypii and Bemisia tabaci
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials
5.2. Sample Preparation for Evaluation of Rotenone and Deguelin Contents in Acetonitrile Extracts
5.3. Standard Calibration
5.4. Toxicity of Rotenone, Deguelin, and Leaf Extracts of Tephroisa Plants against Aphis gossypii
5.5. Toxicity of Rotenone, Deguelin, and Leaf Extracts of Tephroisa Plants against Bemisia tabaci
5.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, P.; Qin, D.; Chen, J.; Zhang, Z. Plants in the Genus Tephrosia: Valuable Resources for Botanical Insecticides. Insects 2020, 11, 721. [Google Scholar] [CrossRef] [PubMed]
- Touqeer, S.; Saeed, M.A.; Ajaib, M. A review on the phytochemistry and pharmacology of genus Tephrosia. Phytopharmacology 2013, 4, 598–637. [Google Scholar]
- Nayak, P. Botanical pesticides: An insecticide from plant derivatives. Biot. Res. Today 2020, 2, 727–730. [Google Scholar]
- Cabras, P.; Caboni, P.; Cabras, M.; Angioni, A.; Russo, M. Rotenone Residues on Olives and in Olive Oil. J. Agric. Food Chem. 2002, 50, 2576–2580. [Google Scholar] [CrossRef]
- Manda, R.R.; Addanki, V.A.; Srivastava, S. Microbial bio-pesticides and botanicals as an alternative to synthetic pesticides in the sustainable agricultural production. Plant Cell Biotechnol. Mol. Biol. 2020, 21, 31–48. [Google Scholar]
- Baker, B. Rotenone Use in Organic Farming. Hygeia Anal 2017. Available online: https://hygeia-analytics.com/2017/01/04/rotenone-use-in-organic-farming/ (accessed on 20 September 2020).
- Stevenson, P.C.; Belmain, S.R. Pesticidal Plants in African Agriculture: Local Uses and Global Perspectives. Outlooks Pest Manag. 2016, 27, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Irvine, J.E.; Freyre, R.H. Source Materials for Rotenone, Occurrence of Rotenoids in Some Species of the Genus Tephrosia. J. Agric. Food Chem. 1959, 7, 106–107. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides in the twenty-first century—Fulfilling their promise? Annu. Rev. Entomol. 2020, 65, 233–249. [Google Scholar] [CrossRef] [Green Version]
- Kocdor, M.A.; Cengiz, H.; Ates, H.; Kocdor, H. Inhibition of Cancer Stem-Like Phenotype by Curcumin and Deguelin in CAL-62 Anaplastic Thyroid Cancer Cells. Anti-Cancer Agents Med. Chem. 2019, 19, 1887–1898. [Google Scholar] [CrossRef]
- Mikami, A.Y.; Ventura, M.U.; Andrei, C.C. Brown stink bug mortality by seed extracts of Tephrosia Vogelii containing deguelin and tephrosin. Braz. Arch. Biol. Technol. 2018, 61. [Google Scholar] [CrossRef]
- Roark, R.C. Tephrosia as an Insecticide—A Review of the Literature; Roark, R.C., Ed.; U.S. Department of Agriculture, Bureau of Entomology and Plant Quarantine, Division of Insecticide Investigations, E-402: Washington, DC, USA, 1937; pp. 1–165. [Google Scholar]
- Delfel, N.E.; Tallent, W.H.; Carlson, D.G.; Wolff, I.A. Distribution of rotenone and deguelin in Tephrosia vogelii and separation of rotenoid-rich fractions. J. Agric. Food Chem. 1970, 18, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Mkindi, A.G.; Tembo, Y.; Mbega, E.R.; Medvecky, B.; Kendal-Smith, A.; Farrell, I.W.; Ndakidemi, P.A.; Belmain, S.R.; Stevenson, P.C. Phytochemical Analysis of Tephrosia vogelii across East Africa Reveals Three Chemotypes that Influence Its Use as a Pesticidal Plant. Plants 2019, 8, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, N. Rotenone—A review of its toxicity and use for fisheries management. Sci. Conserv. 2003, 211, 1–40. [Google Scholar]
- Tuli, H.S.; Mittal, S.; Loka, M.; Aggarwal, V.; Aggarwal, D.; Masurkar, A.; Kaur, G.; Varol, M.; Sak, K.; Kumar, M.; et al. Deguelin targets multiple oncogenic signaling pathways to combat human malignancies. Pharmacol. Res. 2021, 166, 105487. [Google Scholar] [CrossRef]
- Belmain, S.R.; Amoah, B.A.; Nyirenda, S.P.; Kamanula, J.F.; Stevenson, P.C. Highly Variable Insect Control Efficacy of Tephrosia vogelii Chemotypes. J. Agric. Food Chem. 2012, 60, 10055–10063. [Google Scholar] [CrossRef]
- Gaskins, M.H.; White, G.A.; Martin, F.W.; Delfel, N.E.; Ruppel, E.G.; Barnes, D.K. Tephrosia vogelii: A Source of Rotenoids for Insecticidal and Piscicidal Use; Technical Bulletins 171886; United States Department of Agriculture, Economic Research Service: Washington, DC, USA, 1972.
- Krasavtsev, O.A. Frost hardening of woody plants at temperatures below zero. In Cellular Injury and Resistance in Freezing Organisms, Proceedings of the International Conference on Low Temperature Science, Sapporo, Japan, 14–19 1966; Asahina, E., Ed.; Bunyeido Printing Co.: Sapporo, Japan, 1966; pp. 131–141. [Google Scholar]
- Burke, M.J.; Gusta, L.V.; Quamme, H.A.; Weiser, C.J.; Li, P.H. Freezing and Injury in Plants. Annu. Rev. Plant Physiol. 1976, 27, 507–528. [Google Scholar] [CrossRef]
- Sakai, A.; Larcher, W. Frost Survival of Plants: Responses and Adaptation to Freezing Stress; Springer: Berlin/Heidelberg, Germany, 2012; ISBN 3642717454. [Google Scholar]
- Ambroise, V.; Legay, S.; Guerriero, G.; Hausman, J.-F.; Cuypers, A.; Sergeant, K. The Roots of Plant Frost Hardiness and Tolerance. Plant Cell Physiol. 2019, 61, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Irvine, J.E.; Freyre, R.H. Effect of Planting Time and Photoperiod on Tephrosia vogelii. Agron. J. 1966, 58, 49–51. [Google Scholar] [CrossRef]
- Walia, S.; Kumar, D.; Koul, O. Botanical biopesticides in pest management: Potential and constraints. In Biopesticides in Environment and Food Security: Issues and Strategies; Koul, O., Dhaliwal, G.S., Khokhar, S., Ram, S., Eds.; Scientific Publishers (India): Jodhpur, India, 2012; pp. 182–211. [Google Scholar]
- Mwangi, F.N. Evaluation of Botanical Pesticides and Coloured Sticky Insect Traps for Management of Insect Pests (Thrips, Whiteflies and Aphids) in French Beans (Phaseolus Vulgaris L.). Master’s Thesis, University of Nairobi, Nairobi, Kenya, 2015. [Google Scholar]
- Kayange, C.D.M.; Njera, D.; Nyirenda, S.P.; Mwamlima, L. Effectiveness of Tephrosia vogelii and Tephrosia candida Extracts against Common Bean Aphid (Aphis fabae) in Malawi. Adv. Agric. 2019, 2019, 6704834. [Google Scholar] [CrossRef] [Green Version]
- Said, A.H.; Solhaug, A.; Sandvik, M.; Msuya, F.E.; Kyewalyanga, M.S.; Mmochi, A.J.; Lyche, J.L.; Hurem, S. Isolation of the Tephrosia vogelii extract and rotenoids and their toxicity in the RTgill-W1 trout cell line and in zebrafish embryos. Toxicon 2020, 183, 51–60. [Google Scholar] [CrossRef]
- Kerebba, N.; Oyedeji, A.O.; Byamukama, R.; Kuria, S. Pesticidal activity of Tithonia diversifolia (Hemsl.) A. Gray and Tephrosia vogelii (Hook f.); phytochemical isolation and characterization: A review. S. Afr. J. Bot. 2019, 121, 366–376. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Species | Origin | Content (% in m/m) | |||
|---|---|---|---|---|---|---|
| November | February | May | August | |||
| 1 | Tephrosia adunca | Venezuela (PI 308578) | 0.0215 ± 0.0003 aDE z | 0.2014 ± 0.0163 aC | 0.1502 ± 0.0179 aB | 0.1375 ± 0.0004 aC |
| 2 | Tephrosia adunca | Venezuela (PI 308579) | 0.0195 ± 0.0009 aE | 0.0461 ± 0.0080 bDE | 0.0356 ± 0.0035 bC | 0.0293 ± 0.0002 bJ |
| 3 | Tephrosia candida | Brazil (PI 337100) | 0.0257 ± 0.0001 bC | 0.1741 ± 0.0045 aCD | 0.1198 ± 0.0538 aB | 0.1012 ± 0.0043 aE |
| 4 | Tephrosia candida | South Africa (DLEG 990070) | 0.0422 ± 0.0008 aA | 0.1198 ± 0.0929 aCDE | 0.1172 ± 0.0073 aB | 0.0821 ± 0.0009 bF |
| 5 | Tephrosia candida | US, Florida (PI No. unknown) | 0.0089 ± 0.0004 cF | 0.1984 ± 0.0120 aC | 0.1202 ± 0.0066 aB | 0.0663 ± 0.0001 cG |
| 6 | Tephrosia grandiflora | US, California (DLEG 900611) | 0.0000 ± 0.0000 aH | 0.0188 ± 0.0101 aE | 0.0342 ± 0.0019 aC | 0.0370 ± 0.0004 aI |
| 7 | Tephrosia grandiflora | US, Arizona (DLEG 930029) | 0.0000 ± 0.0000 aH | 0.0043 ± 0.0022 aE | 0.0058 ± 0.0001 bC | 0.0012 ± 0.0001 bK |
| 8 | Tephrosia grandiflora | South Africa (DLEG 910177D) | 0.0000 ± 0.0000 aH | 0.0022 ± 0.0011 aE | 0.0062 ± 0.0001 bC | 0.0013 ± 0.0001 bK |
| 9 | Tephrosia vogelii | Congo (PI 213396) | 0.0030 ± 0.0002 dG | 0.0242 ± 0.0054 cE | 0.0264 ± 0.0011 cC | 0.0494 ± 0.0001 eH |
| 10 | Tephrosia vogelii | US, Puerto Rico (PI 257533) | 0.0229 ± 0.0012 bD | 0.7498 ± 0.0594 bB | 0.6658 ± 0.0455 aA | 0.8487 ± 0.0041 aA |
| 11 | Tephrosia vogelii | Kenya (PI 305346) | 0.0389 ± 0.0023 aB | 0.9713 ± 0.1060 aA | 0.7034 ± 0.0316 aA | 0.8110 ± 0.0007 bB |
| 12 | Tephrosia vogelii | Bolivia (PI 387870) | 0.0221 ± 0.0006 bD | 0.0141 ± 0.0089 cE | 0.1524 ± 0.0334 bB | 0.0847 ± 0.0037 dF |
| 13 | Tephrosia vogelii | US, Puerto Rico (PI 574535) | 0.0112 ± 0.0005 cF | 0.0108 ± 0.0071 cE | 0.0743 ± 0.0152 bcBC | 0.1285 ± 0.0017 cD |
| NO. | Species | Origin | Content (% in m/m) | |||
|---|---|---|---|---|---|---|
| November | February | May | August | |||
| 1 | Tephrosia adunca | Venezuela (PI 308578) | 0.0211 ± 0.0006 aE z | 0.1498 ± 0.0192 aC | 0.0901 ± 0.0084 bD | 0.1101 ± 0.0003 bF |
| 2 | Tephrosia adunca | Venezuela (PI 308579) | 0.0211 ± 0.0012 aE | 0.2230 ± 0.0149 aC | 0.1485 ± 0.0106 aD | 0.2957 ± 0.0012 aE |
| 3 | Tephrosia candida | Brazil (PI 337100) | 0.0000 ± 0.0000 bF | 0.0468 ± 0.0020 aC | 0.0158 ± 0.0024 bD | 0.0297 ± 0.0008 bH |
| 4 | Tephrosia candida | South Africa (DLEG 990070) | 0.0000 ± 0.0000 bF | 0.0163 ± 0.0082 aC | 0.0140 ± 0.0013 bD | 0.0045 ± 0.0045 cI |
| 5 | Tephrosia candida | US, Florida (PI No. unknown) | 0.0177 ± 0.0010 aE | 0.1444 ± 0.0662 aC | 0.1101 ± 0.0019 aD | 0.0777 ± 0.0090 aG |
| 6 | Tephrosia grandiflora | US, California (DLEG 900611) | 0.1062 ± 0.0021 aCD | 2.1064 ± 0.3909 aB | 1.1226 ± 0.0774 aC | 0.7939 ± 0.0157 aC |
| 7 | Tephrosia grandiflora | US, Arizona (DLEG 930029) | 0.0111 ± 0.0008 bEF | 0.1412 ± 0.0568 bC | 0.0107 ± 0.0011 bD | 0.0118 ± 0.0002 bHI |
| 8 | Tephrosia grandiflora | South Africa (DLEG 910177D) | 0.0000 ± 0.0000 cF | 0.0424 ± 0.0025 bC | 0.0218 ± 0.0037 bD | 0.0125 ± 0.0001 bHI |
| 9 | Tephrosia vogelii | Congo (PI 213396) | 0.0949 ± 0.0028 cD | 1.8263 ± 0.3812 cB | 0.9442 ± 0.0515 cC | 0.7809 ± 0.0086 cC |
| 10 | Tephrosia vogelii | US, Puerto Rico (PI 257533) | 0.1046 ± 0.0092 cCD | 2.5960 ± 0.2347 bcB | 1.0358 ± 0.2452 cC | 0.4178 ± 0.0004 dD |
| 11 | Tephrosia vogelii | Kenya (PI 305346) | 0.1129 ± 0.0023 cC | 2.4223 ± 0.2885 cB | 1.1009 ± 0.1177 cC | 0.4389 ± 0.0010 dD |
| 12 | Tephrosia vogelii | Bolivia (PI 387870) | 0.6222 ± 0.0107 aA | 4.0757 ± 0.1076 abA | 2.0828 ± 0.2709 bB | 1.3542 ± 0.0110 bB |
| 13 | Tephrosia vogelii | US, Puerto Rico (PI 574535) | 0.3102 ± 0.0102 bB | 4.4269 ± 0.9611 aA | 2.9027 ± 0.1128 aA | 2.0860 ± 0.0168 aA |
| Treatment | Target Organism | Time | LC50 (mg/L) | 95% Confidence Limit (mg/L) | Regression Equations (Y = m + bx) | R2 |
|---|---|---|---|---|---|---|
| Rotenone | Nymph | 24 h | 2.38 | 2.02–2.81 | Y = 3.9472 + 2.7919x | 0.9543 |
| 48 h | 2.06 | 1.74–2.43 | Y = 4.1754 + 2.6329x | 0.9428 | ||
| Adult | 24 h | 2.98 | 2.46–3.60 | Y = 3.7602 + 2.6177x | 0.9775 | |
| 48 h | 2.56 | 2.12–3.09 | Y = 4.0216 + 2.3956x | 0.9532 | ||
| Deguelin | Nymph | 24 h | 9.03 | 7.10–11.48 | Y = 2.5669 + 2.5462x | 0.9966 |
| 48 h | 7.75 | 5.99–10.03 | Y = 3.1992 + 2.0250x | 0.9880 | ||
| Adult | 24 h | 12.10 | 8.74–16.75 | Y = 2.6502 + 2.1701x | 0.9738 | |
| 48 h | 10.88 | 7.43–15.91 | Y = 3.3679 + 1.5746x | 0.9579 |
| Treatment | Target Organism | Time | LC50 (mg/L) | 95% Confidence Limit (mg/L) | Regression Equations (Y = m + bx) | R2 |
|---|---|---|---|---|---|---|
| Rotenone | Nymph | 24 h | 14.84 | 11.84–18.61 | Y = 2.9108 + 1.7835x | 0.9862 |
| 48 h | 12.41 | 9.97–15.44 | Y = 3.0114 + 1.8184x | 0.9733 | ||
| Adult | 24 h | 16.50 | 13.10–20.79 | Y = 2.8053 + 1.8027x | 0.9866 | |
| 48 h | 13.73 | 10.85–17.38 | Y = 3.0999 + 1.6702x | 0.9740 | ||
| Deguelin | Nymph | 24 h | 53.95 | 42.74–68.09 | Y = 1.8779 + 1.8027x | 0.9942 |
| 48 h | 37.94 | 30.79–46.74 | Y = 1.9612 + 1.9245x | 0.9868 | ||
| Adult | 24 h | 61.80 | 49.07–77.82 | Y = 1.4796 + 1.9656x | 0.9860 | |
| 48 h | 41.90 | 33.79–51.95 | Y = 1.9916 + 1.8545x | 0.9912 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Zhang, M.; Mellich, T.A.; Pearson, B.J.; Chen, J.; Zhang, Z. Variation in Rotenone and Deguelin Contents among Strains across Four Tephrosia Species and Their Activities against Aphids and Whiteflies. Toxins 2022, 14, 339. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050339
Zhang P, Zhang M, Mellich TA, Pearson BJ, Chen J, Zhang Z. Variation in Rotenone and Deguelin Contents among Strains across Four Tephrosia Species and Their Activities against Aphids and Whiteflies. Toxins. 2022; 14(5):339. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050339
Chicago/Turabian StyleZhang, Peiwen, Mengzi Zhang, Terri A. Mellich, Brian J. Pearson, Jianjun Chen, and Zhixiang Zhang. 2022. "Variation in Rotenone and Deguelin Contents among Strains across Four Tephrosia Species and Their Activities against Aphids and Whiteflies" Toxins 14, no. 5: 339. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050339