Occurrence, Serotypes and Virulence Characteristics of Shiga-Toxin-Producing Escherichia coli Isolates from Goats on Communal Rangeland in South Africa

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. STEC Occurrence
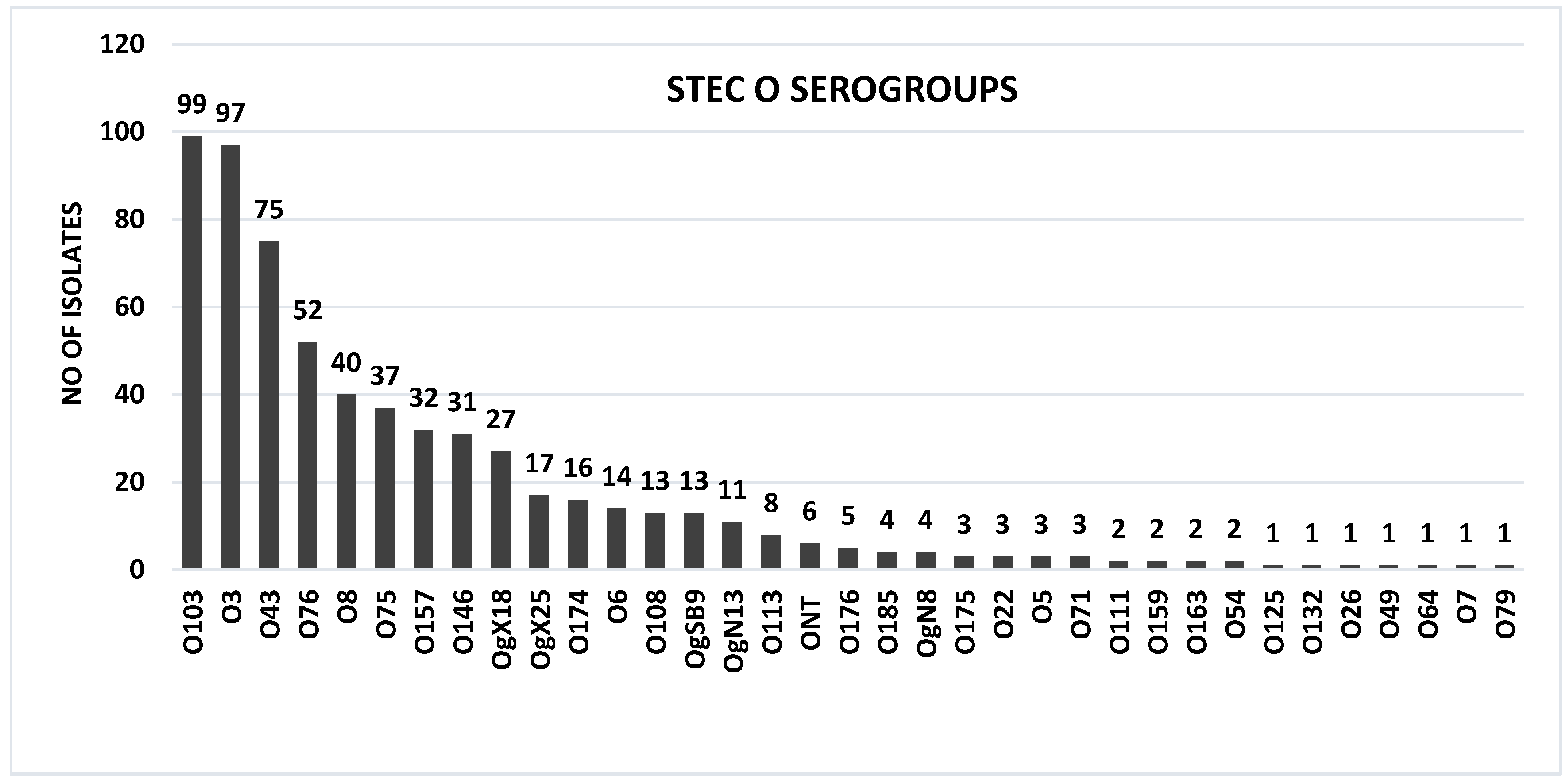
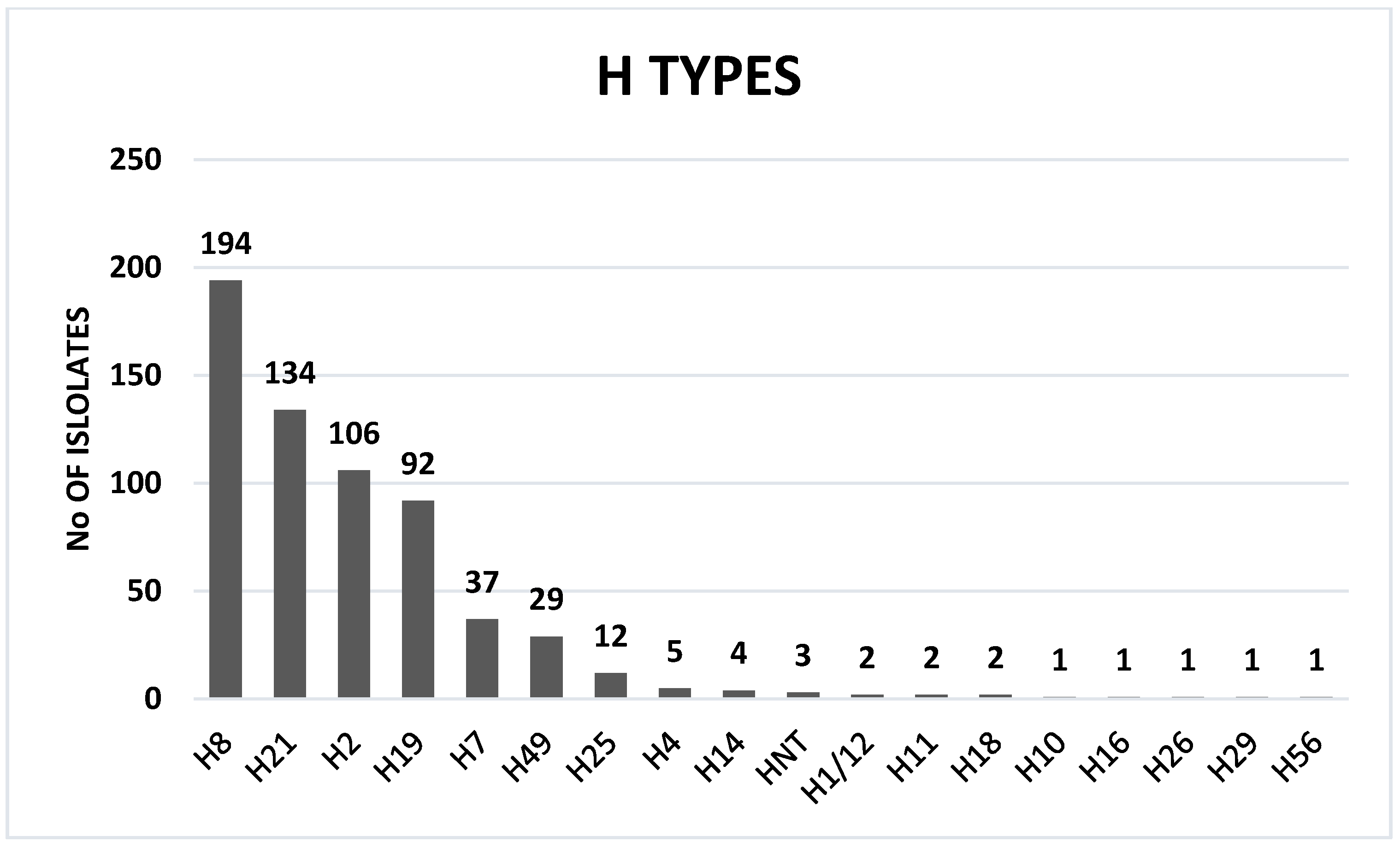
2.2. STEC O:H Serotypes
2.3. STEC Virulence Characteristics
3. Discussion
4. Conclusions
5. Materials and Methods
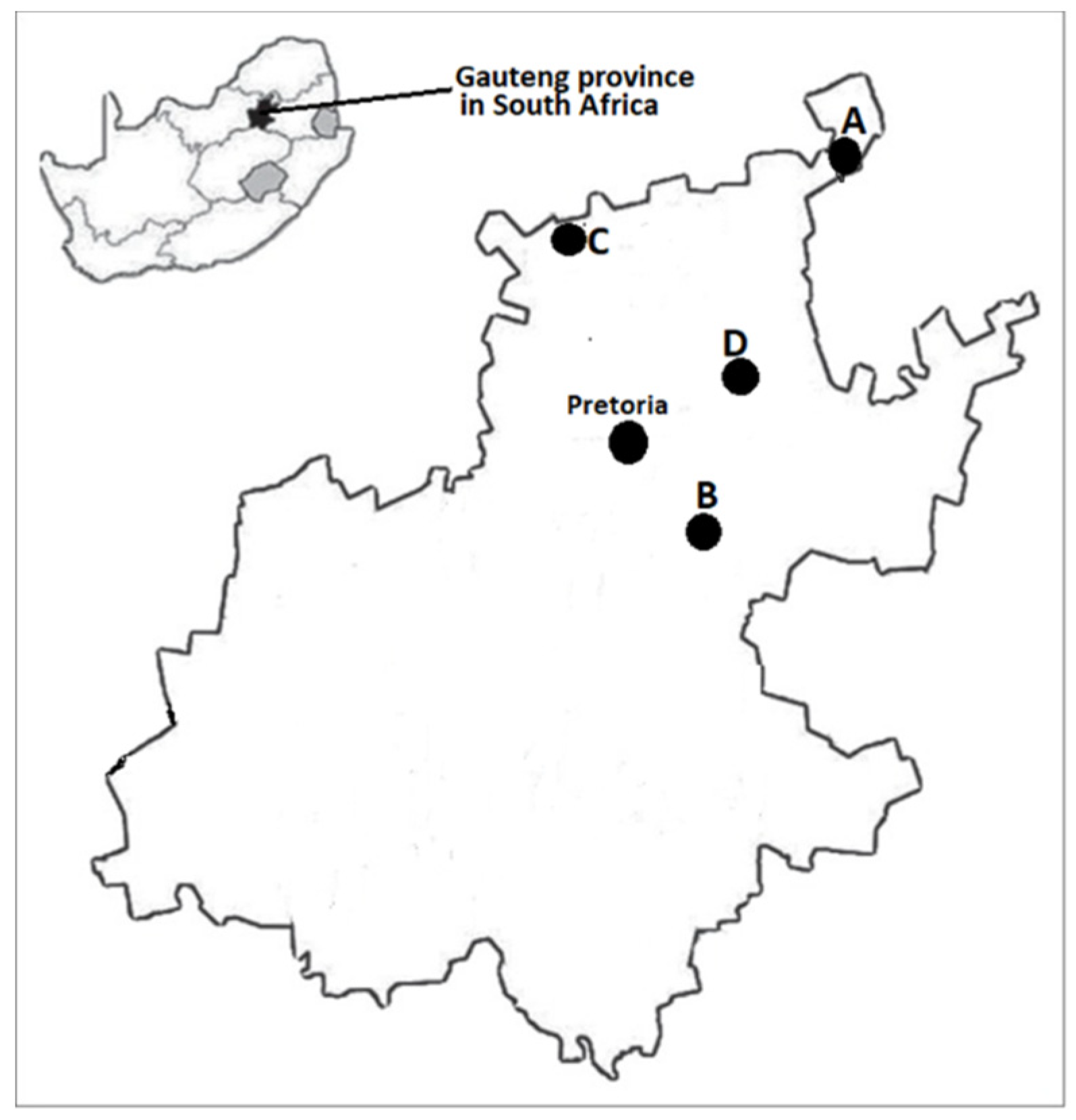
5.1. Study Population and Sample Collection
5.2. STEC Culture
5.3. DNA Extraction and STEC Screening
5.4. STEC Isolation and Identification
5.5. STEC Serotyping
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Döpfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T. World Health Organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar]
- Hussein, H.S. Prevalence and pathogenicity of Shiga toxin-producing Escherichia coli in beef cattle and their products. J. Anim. Sci. 2007, 85, E63–E72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persad, A.K.; Lejeune, J.T. Animal Reservoirs of Shiga Toxin-Producing Escherichia coli. Enterohemorrhagic Escherichia coli and Other Shiga Toxin-Producing E. coli.; American Society of Microbiology: Washington, DC, USA, 2015; pp. 211–230. [Google Scholar]
- La Ragione, R.M.; Best, A.; Woodward, M.J.; Wales, A.D. Escherichia coli O157:H7 colonization in small domestic ruminants. FEMS Microbiol. Rev. 2009, 33, 394–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, S.C.; Burgess, C.M.; Fanning, S.; Duffy, G. An Overview of Shiga-toxin producing Escherichia coli carriage and prevalence in the ovine meat production chain. Foodborne Pathog. Dis. 2021, 18, 147–168. [Google Scholar] [CrossRef] [PubMed]
- Isaäcson, M.; Canter, P.; Effler, P.; Arntzen, L.; Bomans, P.; Heenan, R. Haemorrhagic colitis epidemic in Africa. Lancet 1993, 341, 961. [Google Scholar] [CrossRef]
- Lynch, M.F.; Tauxe, R.V.; Hedberg, C.W. The growing burden of foodborne outbreaks due to contaminated fresh produce: Risks and opportunities. Epidemiol. Infect. 2009, 137, 307–315. [Google Scholar] [CrossRef]
- Nichols, M.C.; Gacek, P.; Phan, Q.; Gambino-Shirley, K.J.; Gollarza, L.M.; Schroeder, M.N.; Mercante, A.; Mullins, J.; Blackstock, A.; Laughlin, M.E.; et al. Agritourism and kidding season: A large outbreak of human Shiga toxin-producing Escherichia coli O157 (STEC O157) infections linked to a goat dairy farm—Connecticut, 2016. Front. Vet. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Schlager, S.; Lepuschitz, S.; Ruppitsch, W.; Ableitner, O.; Pietzka, A.; Neubauer, S.; Stoeger, A.; Lassnig, H.; Mikula, C.; Springer, B. Petting zoos as sources of Shiga toxin-producing Escherichia coli (STEC) infections. Int. J. Med. Microbiol. 2018, 308, 927–932. [Google Scholar] [CrossRef]
- Bettelheim, K.A.; Goldwater, P.N. A Compendium of Shigatoxigenic Escherichia coli: Origins, bacteriological and clinical data on the serogroups, serotypes and untypeable strains of E. coli reported between 1980 and 2017, excluding O157: H7. Adv. Microbiol. 2019, 9, 905–909. [Google Scholar] [CrossRef] [Green Version]
- Beutin, L.; Fach, P. Detection of Shiga toxin-producing Escherichia coli from nonhuman sources and strain typing. Microbiol. Spectr. 2014, 2, 263–285. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Scientific Opinion on VTEC-seropathotype and scientific criteria regarding pathogenicity assessment. EFSA J. 2013, 11, 3138. [Google Scholar]
- Riley, L.W.; Remis, R.S.; Helgerson, S.D.; McGee, H.B.; Wells, J.G.; Davis, B.R.; Hebert, R.J.; Olcott, E.S.; Johnson, L.M.; Hargrett, N.T.; et al. Hemorrhagic colitis associated with a rare Escherichia coli serotype N. Engl. J. Med. 1983, 308, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.T.; Sowers, E.G.; Wells, J.G.; Greene, K.D.; Griffin, P.M.; Hoekstra, R.M.; Strockbine, N.A. Non-O157 Shiga toxin–producing Escherichia coli infections in the United States, 1983–2002. J. Infect. Dis. 2005, 192, 1422–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, J.M.; Wilson, M.E.; Johnson, K.E.; Thorpe, C.M.; Sears, C.L. The emerging clinical importance of non-O157 Shiga toxin-producing Escherichia coli. Clin. Infect. Dis. 2006, 43, 1587–1595. [Google Scholar] [CrossRef]
- Karama, M.; Cenci-Goga, B.T.; Malahlela, M.; Smith, A.M.; Keddy, K.H.; El-Ashram, S.; Kabiru, L.M.; Kalake, A. Virulence characteristics and antimicrobial resistance profiles of shiga toxin-producing Escherichia coli isolates from humans in South Africa: 2006–2013. Toxins 2019, 11, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, A.D.; Newland, J.W.; Miller, S.F.; Holmes, R.K.; Smith, H.W.; Formal, S.B. Shiga-like toxin-converting phages from Escherichia coli strains that cause hemorrhagic colitis or infantile diarrhea. Science 1984, 226, 694–696. [Google Scholar] [CrossRef]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [Green Version]
- Scotland, S.; Smith, H.; Willshaw, G.; Rowe, B. Vero cytotoxin production in strain of Escherichia coli is determined by genes carried on bacteriophage. Lancet 1983, 322, 216. [Google Scholar] [CrossRef]
- Strockbine, N.A.; Jackson, M.; Sung, L.; Holmes, R.; O’Brien, A.D. Cloning and sequencing of the genes for Shiga toxin from Shigella dysenteriae type 1. J. Bacteriol. 1988, 170, 1116–1122. [Google Scholar] [CrossRef] [Green Version]
- Boerlin, P.; McEwen, S.A.; Boerlin-Petzold, F.; Wilson, J.B.; Johnson, R.P.; Gyles, C.L. Associations between virulence factors of Shiga toxin-producing Escherichia coli and disease in humans. J. Clin. Microbiol. 1999, 37, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Donohue-Rolfe, A.; Kondova, I.; Oswald, S.; Hutto, D.; Tzipori, S. Escherichia coli 0157: H7 strains that express Shiga toxin (Stx) 2 alone are more neurotropic for gnotobiotic piglets than are isotypes producing only Stx1 or both Stx1 and Stx2. J. Infect. Dis. 2000, 181, 1825–1829. [Google Scholar] [CrossRef] [Green Version]
- Ethelberg, S.; Olsen, K.E.P.; Scheutz, F.; Jensen, C.; Schiellerup, P.; Engberg, J.; Petersen, A.M.; Olesen, B.; Gerner-Smidt, P.; Mølbak, K. Virulence factors for hemolytic uremic syndrome, Denmark. Emerg. Infect. Dis. 2004, 10, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Ostroff, S.M.; Tarr, P.I.; Neill, M.A.; Lewis, J.H.; Hargrett-Bean, N.; Kobayashi, J.M. Toxin genotypes and plasmid profiles as determinants of systemic sequelae in Escherichia coli O157: H7 infections. J. Infect. Dis. 1989, 160, 994–998. [Google Scholar] [CrossRef] [PubMed]
- Persson, S.; Olsen, K.E.P.; Ethelberg, S.; Scheutz, F. Subtyping method for Escherichia coli Shiga toxin (Verocytotoxin) 2 variants and correlations to clinical manifestations. J. Clin. Microbiol. 2007, 45, 2020–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerse, A.E.; Yu, J.; Tll, B.D.; Kaper, J.B. A genetic locus of enteropathogenic Escherichia coli necessary for the production of attaching and effacing lesions on tissue culture cells. Proc. Natl. Acad. Sci. USA 1990, 87, 7839–7843. [Google Scholar] [CrossRef] [Green Version]
- McKee, M.L.; O’Brien, A.D. Truncated enterohemorrhagic Escherichia coli (EHEC) O157: H7 intimin (EaeA) fusion proteins promote adherence of EHEC strains to HEp-2 cells. Infect. Immun. 1996, 64, 2225–2233. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, T.K.; Jarvis, K.G.; Donnenberg, M.S.; Kaper, J.B. A genetic locus of enterocyte effacement conserved among diverse enterobacterial pathogens. Proc. Natl. Acad. Sci. USA 1995, 92, 1664–1668. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, T.K.; Kaper, J.B. A cloned pathogenicity island from enteropathogenic Escherichia coli confers the attaching and effacing phenotype on E. coli K-12. Mol. Microbiol. 1997, 23, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Pai, C.; Kelly, J.; Meyers, G. Experimental infection of infant rabbits with verotoxin-producing Escherichia coli. Infect. Immun. 1986, 51, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Coombes, B.K.; Wickham, M.E.; Mascarenhas, M.; Gruenheid, S.; Finlay, B.B.; Karmali, M.A. Molecular analysis as an aid to assess the public health risk of non-O157 shiga toxin-producing Escherichia coli strains. Appl. Environ. Microbiol. 2008, 74, 2153–2160. [Google Scholar] [CrossRef] [Green Version]
- Karmali, M.A.; Mascarenhas, M.; Shen, S.; Ziebell, K.; Johnson, S.; Reid-Smith, R.; Isaac-Renton, J.; Clark, C.; Rahn, K.; Kaper, J.B. Association of genomic O island 122 of Escherichia coli EDL 933 with Verocytotoxin-producing Escherichia coli seropathotypes that are linked to epidemic and/or serious disease. J. Clin. Microbiol. 2003, 41, 4930–4940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.; Beutin, L.; Karch, H. Molecular analysis of the plasmid-encoded hemolysin of Escherichia coli O157: H7 strain EDL 933. Infect. Immun. 1995, 63, 1055–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.; Hensel, M. Pathogenicity islands in bacterial pathogenesis. Clin. Microbiol. Rev. 2004, 17, 14–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.; Karch, H. Enterohemolytic phenotypes and genotypes of Shiga toxin-producing Escherichia coli O111 strains from patients with diarrhea and hemolytic-uremic syndrome. J. Clin. Microbiol. 1996, 34, 2364–2367. [Google Scholar] [CrossRef] [Green Version]
- Ateba, C.N.; Mbewe, M. Genotypic characterization of Escherichia coli O157: H7 isolates from different sources in the north-west province, South Africa, using enterobacterial repetitive intergenic consensus PCR analysis. Int. J. Mol. Sci. 2014, 15, 9735–9747. [Google Scholar] [CrossRef]
- Karama, M.; Mainga, A.O.; Cenci-Goga, B.T.; Malahlela, M.; El-Ashram, S.; Kalake, A. Molecular profiling and antimicrobial resistance of Shiga toxin-producing Escherichia coli O26, O45, O103, O121, O145 and O157 isolates from cattle on cow-calf operations in South Africa. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mainga, A.O.; Cenci-Goga, B.T.; Malahlela, M.N.; Tshuma, T.; Kalake, A.; Karama, M. Occurrence and characterization of seven major Shiga toxin-producing Escherichia coli serotypes from healthy cattle on cow–calf operations in South Africa. Zoonoses Public Health. 2018, 65, 777–789. [Google Scholar] [CrossRef]
- Cortés, C.; De la Fuente, R.; Blanco, J.; Blanco, M.; Blanco, J.E.; Dhabi, G.; Mora, A.; Justel, P.; Contreras, A.; Sánchez, A.; et al. Serotypes, virulence genes and intimin types of verotoxin-producing Escherichia coli and enteropathogenic E. coli isolated from healthy dairy goats in Spain. Vet. Microbiol. 2005, 110, 67–76. [Google Scholar] [CrossRef]
- Oliveira, M.G.; Brito, J.R.; Gomes, T.A.; Guth, B.E.; Vieira, M.A.; Naves, Z.V.; Vaz, T.M.; Irino, K. Diversity of virulence profiles of Shiga toxin-producing Escherichia coli serotypes in food-producing animals in Brazil. Int. J. Food Microbiol. 2008, 127, 139–146. [Google Scholar] [CrossRef]
- Vu-Khac, H.; Cornick, N.A. Prevalence and genetic profiles of Shiga toxin-producing Escherichia coli strains isolated from buffaloes, cattle, and goats in central Vietnam. Vet. Microbiol. 2008, 126, 356–363. [Google Scholar] [CrossRef]
- Zschöck, M.; Hamann, H.P.; Kloppert, B.; Wolter, W. Shiga-toxin-producing Escherichia coli in faeces of healthy dairy cows, sheep and goats: Prevalence and virulence properties. Lett. Appl. Microbiol. 2000, 31, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Janda, J.; Bláhová, K.; Minaříková, H.; Jíková, E.; Karmali, M.A.; Laubová, J.; Šikulová, J.; Preston, M.A.; Khakhria, R.; et al. Human Escherichia coli O157:H7 infection associated with the consumption of unpasteurized goat’s milk. Epidemiol. Infect. 1997, 119, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Beutin, L.; Geier, D.; Steinruck, H.; Zimmermann, S.; Scheutz, F. Prevalence and some properties of verotoxin (Shiga-like toxin)-producing Escherichia coli in seven different species of healthy domestic animals. J. Clin. Microbiol. 1993, 31, 2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orden, J.; Ruiz-Santa-Quiteria, J.; Blanco, M.; Blanco, J.; Mora, A.; Cid, D.; González, E.; Blanco, J.; De la Fuente, R. Prevalence and characterization of Vero cytotoxin-producing Escherichia coli isolated from diarrhoeic and healthy sheep and goats. Epidemiol. Infect. 2003, 130, 313–321. [Google Scholar] [CrossRef]
- Islam, M.A.; Mondol, A.S.; De Boer, E.; Beumer, R.R.; Zwietering, M.H.; Talukder, K.A.; Heuvelink, A.E. Prevalence and genetic characterization of shiga toxin-producing Escherichia coli isolates from slaughtered animals in Bangladesh. Appl. Environ. Microbiol. 2008, 74, 5414–5421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čobeljić, M.; Dimić, B.; Opačić, D.; Lepšanović, Z.; Stojanović, V.; Lazić, S. The prevalence of Shiga toxin-producing Escherichia coli in domestic animals and food in Serbia. Epidemiol. Infect. 2005, 133, 359–366. [Google Scholar] [CrossRef]
- Schilling, A.-K.; Hotzel, H.; Methner, U.; Sprague, L.D.; Schmoock, G.; El-Adawy, H.; Ehricht, R.; Wöhr, A.-C.; Erhard, M.; Geue, L. Zoonotic agents in small ruminants kept on city farms in Southern Germany. Appl. Environ. Microbiol. 2012, 78, 3785–3793. [Google Scholar] [CrossRef] [Green Version]
- Isler, M.; Wissmann, R.; Morach, M.; Zurfluh, K.; Stephan, R.; Nüesch-Inderbinen, M. Animal petting zoos as sources of Shiga toxin-producing Escherichia coli, Salmonella and extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae. Zoonoses Public Health 2021, 68, 79–87. [Google Scholar] [CrossRef]
- Novotna, R.; Alexa, P.; Hamrik, J.; Madanat, A.; Smola, J.; Cizek, A. Isolation and characterization Shiga toxin-producing Escherichia coli from sheep and goats in Jordan with evidence of multiresistant serotype O157: H7. Vet. Med. Czech 2005, 50, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Fan, R.; Shao, K.; Yang, X.; Bai, X.; Fu, S.; Sun, H.; Xu, Y.; Wang, H.; Li, Q.; Hu, B.; et al. High prevalence of non-O157 Shiga toxin-producing Escherichia coli in beef cattle detected by combining four selective agars. BMC Microbiol. 2019, 19, 213. [Google Scholar] [CrossRef] [Green Version]
- Stromberg, Z.R.; Lewis, G.L.; Marx, D.B.; Moxley, R.A. Comparison of enrichment broths for supporting growth of Shiga toxin-producing Escherichia coli. Curr. Microbiol. 2015, 71, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Gouali, M.; Ruckly, C.; Carle, I.; Lejay-Collin, M.; Weill, F.X. Evaluation of CHROMagar STEC and STEC O104 chromogenic agar media for detection of Shiga Toxin-producing Escherichia coli in stool specimens. J. Clin. Microbiol. 2013, 51, 894–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradel, N.; Livrelli, V.; De Champs, C.; Palcoux, J.B.; Reynaud, A.; Scheutz, F.; Sirot, J.; Joly, B.; Forestier, C. Prevalence and characterization of Shiga toxin-producing Escherichia coli isolated from cattle, food, and children during a one-year prospective study in France. J. Clin. Microbiol. 2000, 38, 1023–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhaegen, B.; Van Damme, I.; Heyndrickx, M.; Botteldoorn, N.; Elhadidy, M.; Verstraete, K.; Dierick, K.; Denayer, S.; De Zutter, L.; De Reu, K. Evaluation of detection methods for non-O157 Shiga toxin-producing Escherichia coli from food. Int. J. Food Microbiol. 2016, 219, 64–70. [Google Scholar] [CrossRef]
- Wylie, J.L.; Van Caeseele, P.; Gilmour, M.W.; Sitter, D.; Guttek, C.; Giercke, S. Evaluation of a new chromogenic agar medium for detection of Shiga toxin-producing Escherichia coli (STEC) and relative prevalences of O157 and non-O157 STEC in Manitoba, Canada. J. Clin. Microbiol. 2013, 51, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Banjo, M.; Iguchi, A.; Seto, K.; Kikuchi, T.; Harada, T.; Scheutz, F.; Iyoda, S. Escherichia coli H-genotyping PCR: A complete and practical platform for molecular H typing. J. Clin. Microbiol. 2018, 56, e00190-18. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, A.; Iyoda, S.; Seto, K.; Morita-Ishihara, T.; Scheutz, F.; Ohnishi, M. Escherichia coli O-genotyping PCR: A comprehensive and practical platform for molecular O serogrouping. J. Clin. Microbiol. 2015, 53, 2427–2432. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, A.; Nishii, H.; Seto, K.; Mitobe, J.; Lee, K.; Konishi, N.; Obata, H.; Kikuchi, T.; Iyoda, S. Additional Og-typing PCR techniques targeting Escherichia coli-novel and Shigella-unique O-antigen biosynthesis gene clusters. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef]
- Keen, J.E.; Wittum, T.E.; Dunn, J.R.; Bono, J.L.; Durso, L.M. Shiga-toxigenic Escherichia coli O157 in agricultural fair livestock, United States. Emerg. Infect. Dis. 2006, 12, 780–786. [Google Scholar] [CrossRef]
- Mersha, G.; Asrat, D.; Zewde, B.M.; Kyule, M. Occurrence of Escherichia coli O157:H7 in faeces, skin and carcasses from sheep and goats in Ethiopia. Lett. Appl. Microbiol. 2010, 50, 71–76. [Google Scholar] [CrossRef]
- Espié, E.; Vaillant, V.; Mariani-Kurkdjian, P.; Grimont, F.; Martin-Schaller, R.; De Valk, H.; Vernozy-Rozand, C. Escherichia coli O157 outbreak associated with fresh unpasteurized goats’ cheese. Epidemiol. Infect. 2006, 134, 143–146. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, L.; Fung, J.; Paccagnella, A.; Isaac-Renton, J.; Rockwell, F.; Emerson, B.; Preston, T. Escherichia coli O157 outbreak associated with the ingestion of unpasteurized goat’s milk in British Columbia, 2001. Can. Commun. Dis. Rep. 2002, 28, 6–8. [Google Scholar] [PubMed]
- Bai, X.; Hu, B.; Xu, Y.; Sun, H.; Zhao, A.; Ba, P.; Fu, S.; Fan, R.; Jin, Y.; Wang, H.; et al. Molecular and phylogenetic characterization of non-O157 Shiga toxin-producing Escherichia coli strains in China. Front. Cell. Infect. Microbiol. 2016, 6, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, D.; Sanz, M.; Parma, A.; Padola, N. Characterization of Shiga toxin-producing Escherichia coli isolated from newborn, milk-fed, and growing calves in Argentina. J. Dairy Sci. 2012, 95, 5340–5343. [Google Scholar] [CrossRef] [PubMed]
- Baba, H.; Kanamori, H.; Kudo, H.; Kuroki, Y.; Higashi, S.; Oka, K.; Takahashi, M.; Yoshida, M.; Oshima, K.; Aoyagi, T.; et al. Genomic analysis of Shiga toxin-producing Escherichia coli from patients and asymptomatic food handlers in Japan. PLoS ONE 2019, 14, e0225340. [Google Scholar] [CrossRef]
- Hong, S.; Oh, K.-H.; Cho, S.-H.; Kim, J.-C.; Park, M.-S.; Lim, H.-S.; Lee, B.-K. Asymptomatic healthy slaughterhouse workers in South Korea carrying Shiga toxin-producing Escherichia coli. FEMS Immunol. Med. Microbiol. 2009, 56, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Horcajo, P.; Domínguez-Bernal, G.; De La Fuente, R.; Ruiz-Santa-Quiterai, J.A.; Orden, J.A. Association of Vt1C with Verotoxin-producing Escherichia coli from goats and sheep. J. Vet. Diagn. Investig. 2010, 22, 332–334. [Google Scholar] [CrossRef] [Green Version]
- Jajarmi, M.; Askari Badouei, M.; Imani Fooladi, A.A.; Ghanbarpour, R.; Ahmadi, A. Pathogenic potential of Shiga toxin-producing Escherichia coli strains of caprine origin: Virulence genes, Shiga toxin subtypes, phylogenetic background and clonal relatedness. BMC Vet. Res. 2018, 14, 97. [Google Scholar] [CrossRef] [Green Version]
- Mahanti, A.; Samanta, I.; Bandyopadhyay, S.; Joardar, S.N. Molecular characterization and antibiotic susceptibility pattern of caprine Shiga toxin producing-Escherichia coli (STEC) isolates from India. Iran. J. Vet. Res. 2015, 16, 31–35. [Google Scholar]
- Wani, S.A.; Samanta, I.; Munshi, Z.H.; Bhat, M.A.; Nishikawa, Y. Shiga toxin-producing Escherichia coli and enteropathogenic Escherichia coli in healthy goats in India: Occurrence and virulence properties. J. Appl. Microbiol. 2006, 100, 108–113. [Google Scholar] [CrossRef]
- EFSA. Pathogenicity assessment of Shiga toxin-producing Escherichia coli (STEC) and the public health risk posed by contamination of food with STEC. EFSA J. 2020, 18, e05967. [Google Scholar]
- Karmali, M.A.; Petric, M.; Lim, C.; Fleming, P.C.; Arbus, G.S.; Lior, H. The association between idiopathic hemolytic uremic syndrome and infection by verotoxin-producing Escherichia coli. J. Infect. Dis. 1985, 151, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Paton, A.W.; Woodrow, M.C.; Doyle, R.M.; Lanser, J.A.; Paton, J.C. Molecular characterization of a Shiga toxigenic Escherichia coli O113:H21 strain lacking eae responsible for a cluster of cases of hemolytic- uremic syndrome. J. Clin. Microbiol. 1999, 37, 3357–3361. [Google Scholar] [CrossRef] [Green Version]
- Paton, A.W.; Paton, J.C. Detection and characterization of Shiga toxigenic Escherichia coli by using multiplex PCR assays for stx1, stx2, eaeA, enterohemorrhagic E. coli hlyA, rfbO111, and rfbO157. J. Clin. Microbiol. 1998, 36, 598–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herds | Herd Occurrence | O Serogroups (N = 34) | Serotypes (N = 63) | Isolates (N = 628) | Number of Goats (N = 289) |
|---|---|---|---|---|---|
| A | (75.3%) 116/154 | O3 (58) | O3:H2 | 1 | 1 |
| O3:H11 | 1 | 1 | |||
| O3:H19 | 1 | 1 | |||
| O3:H21 | 55 | 14 | |||
| O8 (29) | O8:H14 | 1 | 1 | ||
| O8:H19 | 8 | 2 | |||
| O8:H21 | 5 | 1 | |||
| O8:H49 | 15 | 3 | |||
| O22 (3) | O22:H8 | 3 | 1 | ||
| O26 (1) | O26:H2 | 1 | 1 | ||
| O43 (50) | O43:H2 | 49 | 12 | ||
| O43:H8 | 1 | 1 | |||
| O49 (1) | O49:H11 | 1 | 1 | ||
| O54 (2) | O54:H16 | 1 | 1 | ||
| O54:H19 | 1 | 1 | |||
| O64 (1) | O64:H18 | 1 | 1 | ||
| O71 (3) | O71:H1/12 | 2 | 1 | ||
| O71:H14 | 1 | 1 | |||
| O76 (39) | O76:H2 | 1 | 1 | ||
| O76:H19 | 38 | 9 | |||
| O103 (99) | O103:H8 | 98 | 22 | ||
| O103:H56 | 1 | 1 | |||
| O108 (13) | O108:H19 | 1 | 1 | ||
| O108:H25 | 12 | 3 | |||
| O111 (2) | O111:H8 | 2 | 1 | ||
| O113 (7) | O113:H8 | 7 | 3 | ||
| O146 (7) | O146:H21 | 7 | 2 | ||
| O157 (30) | O157:H7 | 29 | 7 | ||
| O157:H8 | 1 | 1 | |||
| O163 (1) | O163:H2 | 1 | 1 | ||
| O175 (2) | O175:H7 | 1 | 1 | ||
| O175:H19 | 1 | 1 | |||
| O185 (4) | O185:H8 | 4 | 3 | ||
| OgN8 (4) | OgN8:H7 | 4 | 1 | ||
| OgN13 (11) | OgN13:H19 | 9 | 1 | ||
| OgN13:H10 | 1 | 1 | |||
| OgN13:H- | 1 | 1 | |||
| OgSB9 (7) | OgSB9:H2 | 1 | 1 | ||
| OgSB9:H19 | 6 | 2 | |||
| OgX18 (2) | OgX18:H2 | 2 | 1 | ||
| OgX25 (15) | OgX25:H8 | 15 | 3 | ||
| ONT (6) | ONT:H18 | 1 | 1 | ||
| ONT:H19 | 1 | 1 | |||
| ONT:H26 | 1 | 1 | |||
| ONT:HNT | 3 | 2 | |||
| B | (90.6%) 39/43 | O5 (3) | O5:H19 | 3 | 1 |
| O6 (14) | O6:H8 | 1 | 1 | ||
| O6:H21 | 1 | 1 | |||
| O6:H49 | 12 | 6 | |||
| O7 (1) | O7:H7 | 1 | 1 | ||
| O8 (3) | O8:H7 | 1 | 1 | ||
| O8:H8 | 2 | 2 | |||
| O8:H14 | 2 | 2 | |||
| O75 (37) | O75:H8 | 37 | 9 | ||
| O76 (8) | O76:H19 | 7 | 4 | ||
| O76:49 | 1 | 1 | |||
| O79 (1) | O79:H8 | 1 | 1 | ||
| O125 (1) | O125:H19 | 1 | 1 | ||
| O132 (1) | O132:H8 | 1 | 1 | ||
| O159 (1) | O159:H49 | 1 | 1 | ||
| O163 (1) | O163:H8 | 1 | 1 | ||
| O176 (5) | O176:H4 | 5 | 1 | ||
| OgSB9 (6) | OgSB9:H19 | 5 | 2 | ||
| OgSB9:H21 | 1 | 1 | |||
| OgX18 (1) | OgX18:H21 | 1 | 1 | ||
| OgX25 (2) | OgX25:H8 | 2 | 1 | ||
| C | (78.8%) 41/52 | O43 (25) | O43:H2 | 24 | 7 |
| O43:H8 | 1 | 1 | |||
| O76 (4) | O76:H19 | 4 | 3 | ||
| O113 (1) | O113:H8 | 1 | 1 | ||
| O146 (11) | O146:H21 | 11 | 3 | ||
| O157 (2) | O157:H7 | 1 | 1 | ||
| O157:H29 | 1 | 1 | |||
| O174 (12) | O174:H8 | 12 | 4 | ||
| O175 (1) | O175:H21 | 1 | 1 | ||
| OgX18 (9) | OgX18:H2 | 9 | 2 | ||
| D | (90%) 36/40 | O3 (39) | O3:H21 | 39 | 9 |
| O8 (6) | O8:H2 | 1 | 1 | ||
| O8:H19 | 5 | 1 | |||
| O76 (1) | O76:H19 | 1 | 1 | ||
| O146 (13) | O146:H21 | 13 | 2 | ||
| O159 (1) | O159:H2 | 1 | 1 | ||
| O174 (4) | O174:H8 | 4 | 1 | ||
| OgX18 (15) | OgX18:H2 | 15 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malahlela, M.N.; Cenci-Goga, B.T.; Marufu, M.C.; Fonkui, T.Y.; Grispoldi, L.; Etter, E.; Kalake, A.; Karama, M. Occurrence, Serotypes and Virulence Characteristics of Shiga-Toxin-Producing Escherichia coli Isolates from Goats on Communal Rangeland in South Africa. Toxins 2022, 14, 353. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050353
Malahlela MN, Cenci-Goga BT, Marufu MC, Fonkui TY, Grispoldi L, Etter E, Kalake A, Karama M. Occurrence, Serotypes and Virulence Characteristics of Shiga-Toxin-Producing Escherichia coli Isolates from Goats on Communal Rangeland in South Africa. Toxins. 2022; 14(5):353. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050353
Chicago/Turabian StyleMalahlela, Mogaugedi N., Beniamino T. Cenci-Goga, Munyaradzi C. Marufu, Thierry Y. Fonkui, Luca Grispoldi, Eric Etter, Alan Kalake, and Musafiri Karama. 2022. "Occurrence, Serotypes and Virulence Characteristics of Shiga-Toxin-Producing Escherichia coli Isolates from Goats on Communal Rangeland in South Africa" Toxins 14, no. 5: 353. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050353