In Vitro Effects of Lemon Balm Extracts in Reducing the Growth and Mycotoxins Biosynthesis of Fusarium culmorum and F. proliferatum

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. The Inhibitory Effect of Lemon Balm Extracts on Fusarium Growth
2.2. The Effect of Lemon Balm Extract on ERG Content
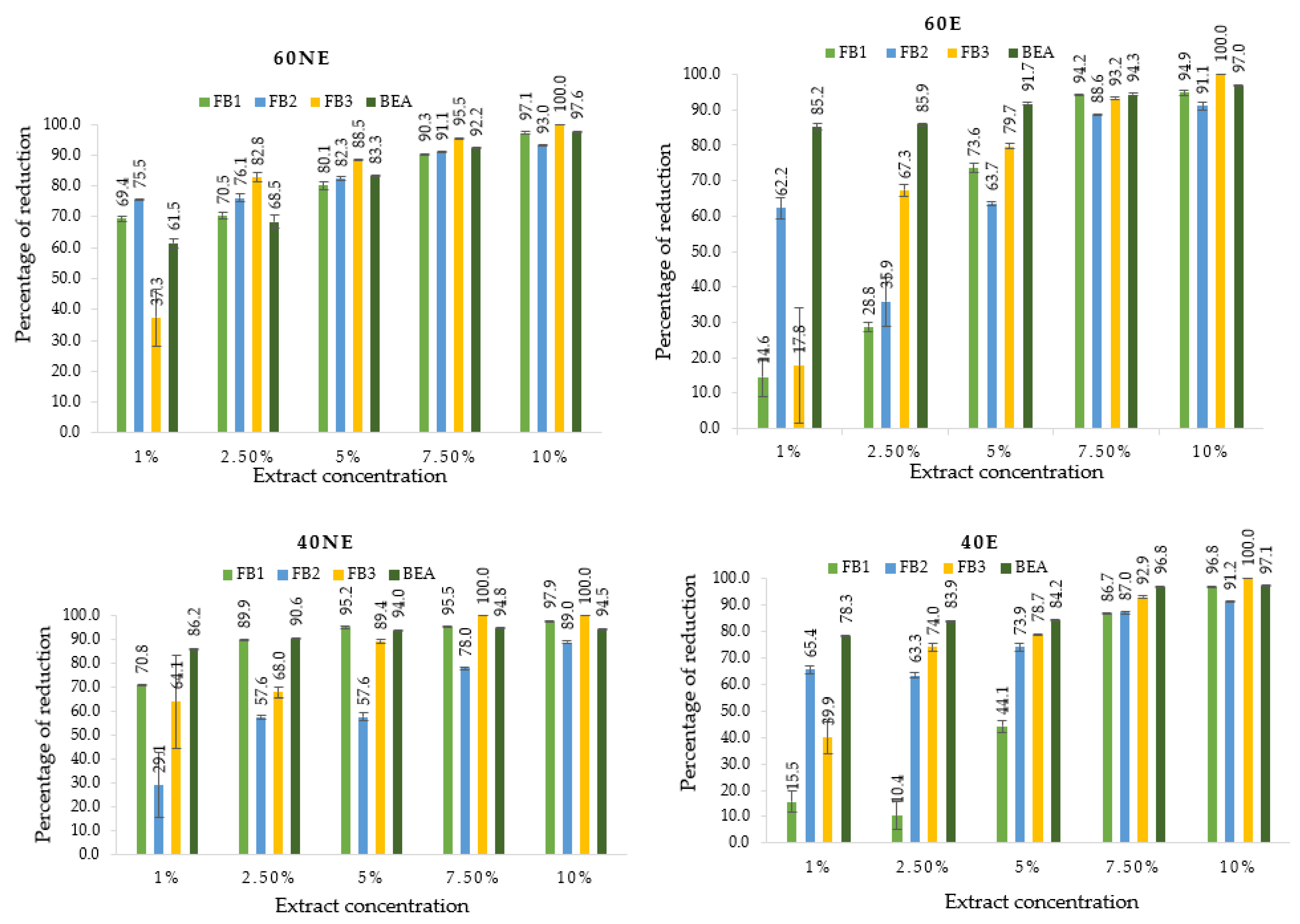
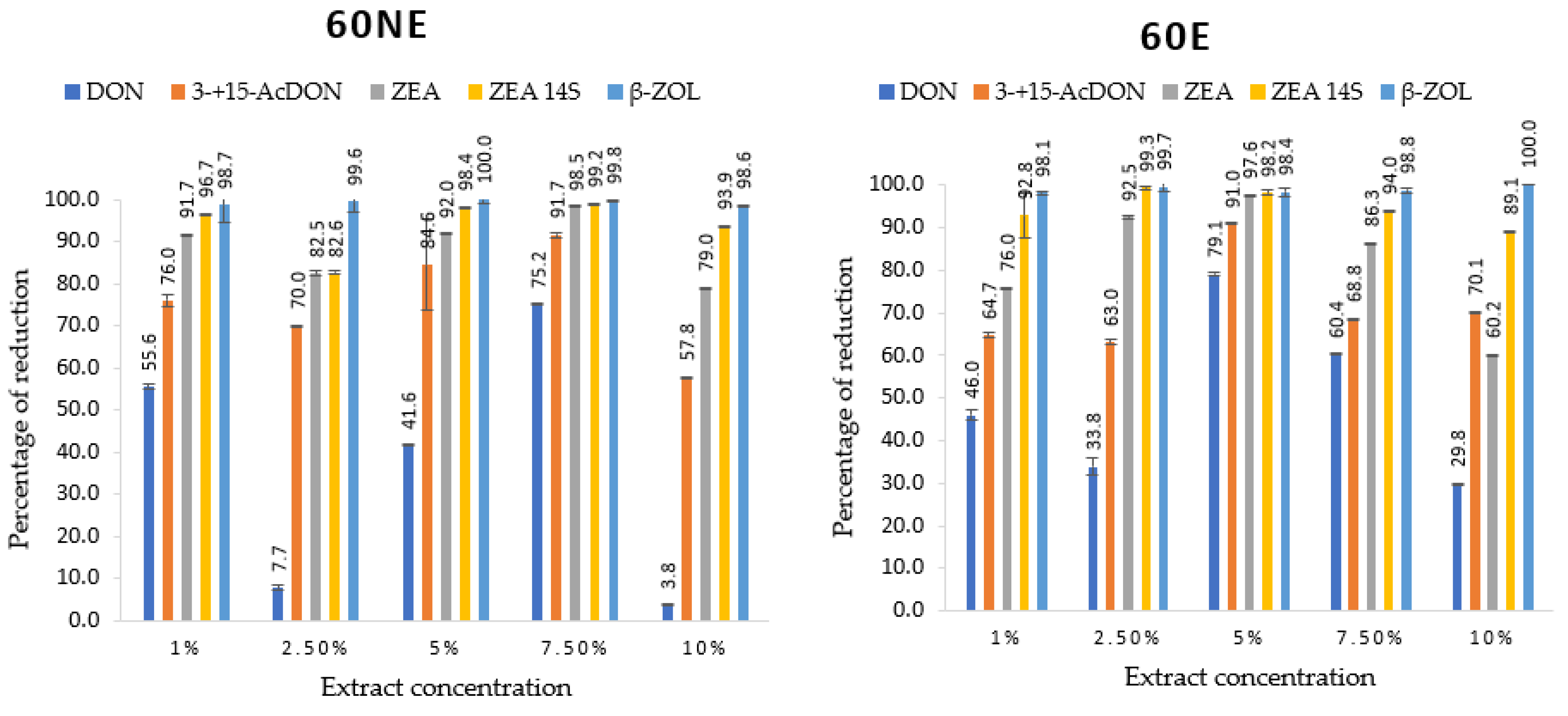
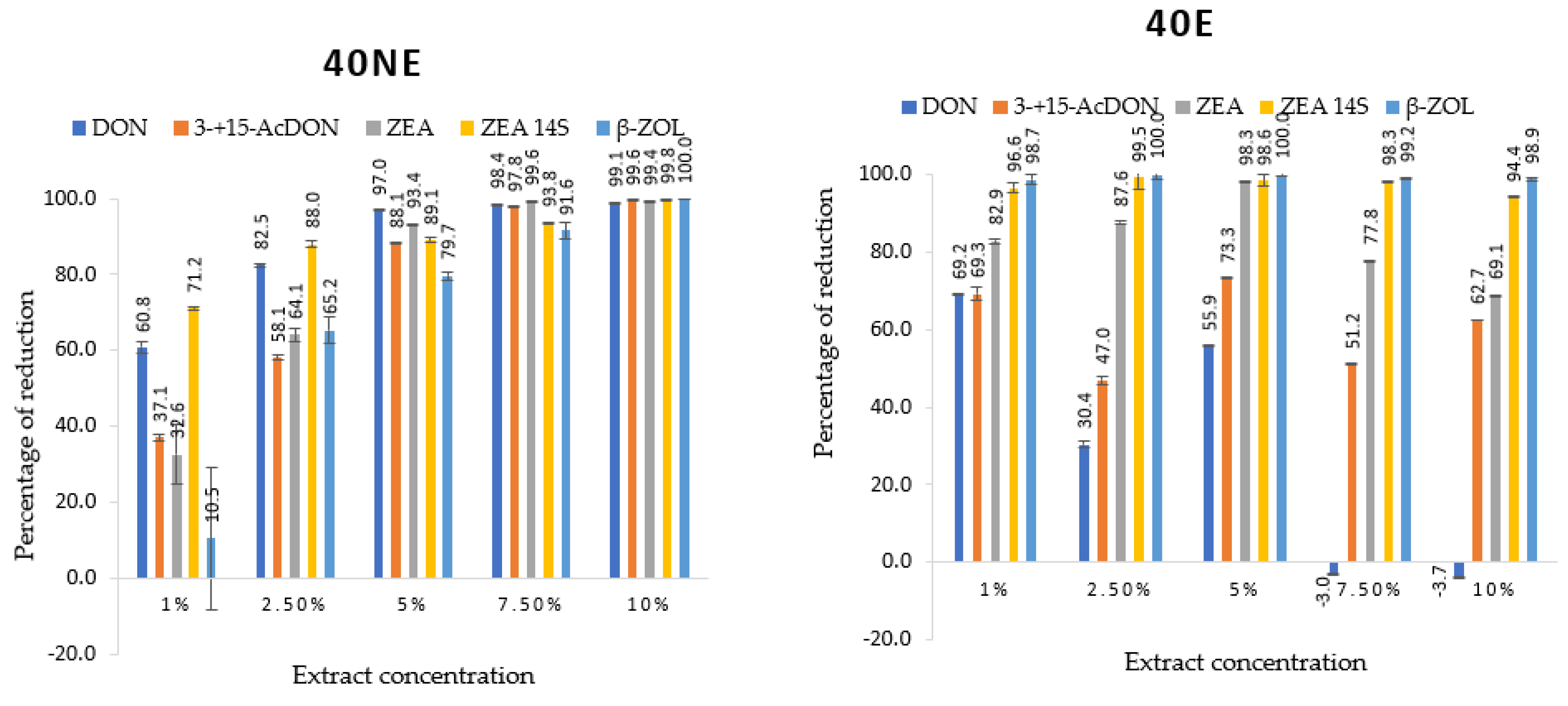
2.3. Mycotoxins Identified and Quantified by UHPLC-HESI-MS/MS and Their Reduction by Lemon Balm Extracts
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Fungal Material
5.3. Chemicals
5.4. Supercritical Carbon Dioxide (SC-CO2) Extraction
5.5. The Effect of Lemon Balm Extracts Prepared at Different Concentration Levels on Mycelia Growth
5.6. Determination of ERG in Mycelia Grown on PDA with Lemon Balm Extract
5.7. Analysis of Mycotoxins Using UHPLC-HESI-MS/MS
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Mallmann, C.A.; Brugnari, T.; de Abreu Filho, B.A.; Mikcha, J.M.G.; Machinski, M. Curcuma Longa L. Essential Oil Composition, Antioxidant Effect, and Effect on Fusarium Verticillioides and Fumonisin Production. Food Control 2017, 73, 806–813. [Google Scholar] [CrossRef]
- Shi, W.; Tan, Y.; Wang, S.; Gardiner, D.M.; De Saeger, S.; Liao, Y.; Wang, C.; Fan, Y.; Wang, Z.; Wu, A. Mycotoxigenic Potentials of Fusarium Species in Various Culture Matrices Revealed by Mycotoxin Profiling. Toxins 2017, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asam, S.; Habler, K.; Rychlik, M. Fusarium Mycotoxins in Food. In Chemical Contaminants and Residues in Food; Woodhead Publishing: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Seefelder, W.; Gossmann, M.; Humpf, H.U. Analysis of Fumonisin B1 in Fusarium proliferatum-Infected Asparagus Spears and Garlic Bulbs from Germany by Liquid Chromatography-Electrospray Ionization Mass Spectrometry. J. Agric. Food Chem. 2002, 50, 2778–2781. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Mulè, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium Species and Mycotoxins Associated with Maize Ear Rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Pinotti, L.; Ottoboni, M.; Giromini, C.; Dell’Orto, V.; Cheli, F. Mycotoxin Contamination in the EU Feed Supply Chain: A Focus on Cereal Byproducts. Toxins 2016, 8, 45. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium Species and Mycotoxins Associated with Head Blight in Small-Grain Cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of Mycotoxins on Humans and Animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Yazar, S.; Omurtag, G.Z. Fumonisins, Trichothecenes, and Zearalenone in Cereals. Int. J. Mol. Sci 2008, 9, 2062–2090. [Google Scholar] [CrossRef]
- Phoku, J.Z.; Dutton, M.F.; Barnard, T.G.; Potgieter, N. Use of a Bio-Wipe Kit to Detect Fumonisin B1 in Faecal Materials. Food Addit. Contam. 2014, 31, 1760–1768. [Google Scholar] [CrossRef]
- Chilaka, C.A.; De Boevre, M.; Atanda, O.O.; De Saeger, S. The Status of Fusarium Mycotoxins in Sub-Saharan Africa: A Review of Emerging Trends and Post-Harvest Mitigation Strategies towards Food Control. Toxins 2017, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal Agent of Foot and Root Rot and Head Blight on Wheat. Mol. Plant Pathol. 2012, 14, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Velluti, A.; Sanchis, V.; Ramos, A.J.; Egido, J.; Marín, S. Inhibitory Effect of Cinnamon, Clove, Lemongrass, Oregano, and Palmarose Essential Oils on Growth and Fumonisin B1 Production by Fusarium proliferatum in Maize Grain. Int. J. Food Microbiol. 2003, 89, 145–154. [Google Scholar] [CrossRef]
- Gálvez, L.; Clarkson, J.P.; Palmero, D. IGS Region Polymorphisms Are Responsible for Failure of Commonly Used Species-Specific Primers in Fusarium proliferatum Isolates from Diseased Garlic. Plant Pathol. 2020, 69, 713–722. [Google Scholar] [CrossRef]
- Mondani, L.; Chiusa, G.; Battilani, P. Fungi Associated with Garlic During the Cropping Season, with Focus on Fusarium proliferatum and F. oxysporum. Plant Health Prog. 2021, 22, 37–46. [Google Scholar] [CrossRef]
- Guo, Z.; Pfohl, K.; Karlovsky, P.; Dehne, H.W.; Altincicek, B. Dissemination of Fusarium proliferatum by Mealworm Beetle Tenebrio Molitor. PLoS ONE 2018, 13, e0204602. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. International Agency for Research on Cancer Iarc Monographs on the Evaluation of Carcinogenic Risks To Humans; International Agency for Research on Cancer: Lyon, France, 2002; Volume 82, pp. 307–366, 390. [Google Scholar]
- Sydenham, E.W.; Thiel, P.G.; Marasas, W.F.O.; Shephard, G.S.; Van Schalkwyk, D.J.; Koch, K.R. Natural Occurrence of Some Fusarium Mycotoxins in Corn from Low and High Esophageal Cancer Prevalence Areas of the Transkei, Southern Africa. J. Agric. Food Chem. 1990, 38, 1900–1903. [Google Scholar] [CrossRef]
- Seo, J.A.; Yu, J.H. Toxigenic Fungi, and Mycotoxins. In Handbook of Industrial Mycology; CRC Press: Boca Raton, FL, USA, 2004; Volume 56, pp. 689–722. [Google Scholar] [CrossRef]
- Wild, C.P.; Gong, Y.Y. Mycotoxins and Human Disease: A Largely Ignored Global Health Issue. Carcinogenesis 2010, 31, 71–82. [Google Scholar] [CrossRef]
- Domijan, A.M. Fumonisin B1: A Neurotoxic Mycotoxin. Arch. Ind. Hyg. Toxicol. 2012, 63, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elsalam, K.A.; Hashim, A.F.; Alghuthaymi, M.A.; Said-Galiev, E. Nanobiotechnological Strategies for Toxigenic Fungi and Mycotoxin Control. In Food Preservation; Elsevier: Amsterdam, The Netherlands, 2017; pp. 337–346. [Google Scholar] [CrossRef]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Bru, C.A.; Kna, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmu, A.; Smalling, K.; et al. Fungicides: An Overlooked Pesticide Class? Environ. Sci. Technol. 2019, 53, 54. [Google Scholar] [CrossRef]
- Mielniczuk, E.; Skwaryło-Bednarz, B. Fusarium Head Blight, Mycotoxins and Strategies for Their Reduction. Agronomy 2020, 10, 509. [Google Scholar] [CrossRef] [Green Version]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium Toxins in Cereals: Occurrence, Legislation, Factors Promoting the Appearance and Their Management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Cruz Cabral, L.; Fernández Pinto, V.; Patriarca, A. Application of Plant Derived Compounds to Control Fungal Spoilage and Mycotoxin Production in Foods. Int. J. Food Microbiol. 2013, 166, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Perczak, A.; Gwiazdowska, D.; Gwiazdowski, R.; Juś, K.; Marchwińska, K.; Waśkiewicz, A. The Inhibitory Potential of Selected Essential Oils on Fusarium Spp. Growth and Mycotoxins Biosynthesis in Maize Seeds. Pathogens 2020, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gakuubi, M.M.; Maina, A.W.; Wagacha, J.M. Antifungal Activity of Essential Oil of Eucalyptus camaldulensis Dehnh. against Selected Fusarium Spp. Int. J. Microbiol. 2017, 8761610. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.R.N. An Overview of Plant-Derived Products on Control of Mycotoxigenic Fungi and Mycotoxins Fungal Diseases on Red-Fleshed Dragon Fruit (Hylocereus Polyrhizus) in Malaysia View Project Ochratoxins in Grapevine: A Risk Caused by Aspergillus Spp. View Project. Artic. Asian J. Plant Sci. 2010, 9, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, M.; Nasrin, L.; Podder, S. Investigation of Potential Biological Control of Fusarium oxysporum f.Sp.Lycopersici by Plant Extracts, Antagonistic Sp. and Chemical Elicitors In Vitro. Fungal Genom. Biol. 2018, 8, 8–11. [Google Scholar] [CrossRef]
- Střelková, T.; Nemes, B.; Kovács, A.; Novotný, D.; Božik, M.; Klouček, P. Inhibition of Fungal Strains Isolated from Cereal Grains via Vapor Phase of Essential Oils. Molecules 2021, 26, 1313. [Google Scholar] [CrossRef] [PubMed]
- Aziz, Z.A.A.; Ahmad, A.; Setapar, S.H.M.; Karakucuk, A.; Azim, M.M.; Lokhat, D.; Rafatullah, M.; Ganash, M.; Kamal, M.A.; Ashraf, G.M. Essential Oils: Extraction Techniques, Pharmaceutical And Therapeutic Potential-A Review. Curr. Drug Metab. 2018, 19, 1100–1110. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, A.; Sahebkar, A.; Javadi, B. Melissa Officinalis L: A Review of Its Traditional Uses, Phytochemistry and Pharmacology. J. Ethnopharmacol. 2016, 188, 204–228. [Google Scholar] [CrossRef]
- Sawicka, B.; Pszczółkowski, P.; Barbas, P.; Bienia, B. Botanical, Geographical, Herbicidal Aspects of Lemon Balm (Melissa officinalis L.). Acta Sci. Agric. 2020, 4, 34–39. [Google Scholar] [CrossRef]
- Moradkhani, H.; Sargsyan, E.; Bibak, H.; Naseri, B.; Sadat-Hosseini, M.; Fayazi-Barjin, A.; Meftahizade, H. Melissa officinalis L., a Valuable Medicine Plant: A Review. J. Med. Plants Res. 2010, 4, 2753–2759. [Google Scholar]
- Skotti, E.P.; Sotiropoulou, Ν.D.; Lappa, I.K.; Kaiafa, M.; Dimitrios, I. Screening of Lemon Balm Extracts for Anti-Aflatoxigenic, Antioxidant and Other Biological Activities. Life Sci. 2019, 1–16. [Google Scholar] [CrossRef]
- Herodež, Š.S.; Hadolin, M.; Škerget, M.; Knez, Ž. Solvent Extraction Study of Antioxidants from Balm (Melissa officinalis L.) Leaves. Food Chem. 2003, 80, 275–282. [Google Scholar] [CrossRef]
- Bogdanovic, A.; Tadic, V.; Arsic, I.; Milovanovic, S.; Petrovic, S.; Skala, D. Supercritical and High Pressure Subcritical Fluid Extraction from Lemon Balm (Melissa officinalis L., Lamiaceae). J. Supercrit. Fluids 2016, 107, 234–242. [Google Scholar] [CrossRef]
- Miraj, S.; Rafieian-Kopaei; Kiani,, S. Melissa officinalis L: A Review Study With an Antioxidant Prospective. J. Evid. -Based Complementary Altern. Med. 2017, 22, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Cunha, F.; Tintino, S.R.; Figueredo, F.; Barros, L.; Duarte, A.E.; Vega Gomez, M.C.; Coronel, C.; Rolón, M.; Leite, N.; Sobral-Souza, C.E.; et al. Pharmaceutical Biology HPLC-DAD Phenolic Profile, Cytotoxic and Anti-Kinetoplastidae Activity of Melissa officinalis HPLC-DAD Phenolic Profile, Cytotoxic and Anti-Kinetoplastidae Activity of Melissa officinalis. Pharm. Biol. 2016, 54, 1664–1670. [Google Scholar] [CrossRef] [Green Version]
- Mabrouki, H.; Duarte, C.M.M.; Akretche, D.E. Estimation of Total Phenolic Contents and In Vitro Antioxidant and Antimicrobial Activities of Various Solvent Extracts of Melissa officinalis L. Arab. J. Sci. Eng. 2018, 43, 3349–3357. [Google Scholar] [CrossRef]
- Mimica-Dukic, N.; Bozin, B.; Sokovic, M.; Simin, N. Antimicrobial and Antioxidant Activities of Melissa officinalis L. (Lamiaceae) Essential Oil. J. Agric. Food Chem. 2004, 52, 2485–2489. [Google Scholar] [CrossRef]
- Moacs, E.-A.; Farcag, C.; Ghiuu, A.; Coricovac, D.; Popovici, R.; Csrsba-Meius, N.-L.; Ardelean, F.; Antal, D.S.; Dehelean, C.; Avram, F. A Comparative Study of Melissa officinalis Leaves and Stems Ethanolic Extracts in Terms of Antioxidant, Cytotoxic, and Antiproliferative Potential. Evid.-Based Complementary Altern. Med. 2018, 7860456. [Google Scholar] [CrossRef] [Green Version]
- Sibel, O.; Dervisoglu, M.; Azime, Y. Antifungal Activity of Lemon Balm and Sage Essential Oils on the Growth of Ochratoxigenic Penicillium Verrucosum. Afr. J. Microbiol. Res. 2012, 6, 3079–3084. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, H.; Matsubara, Y.-I. Effect of Lemon Balm Water Extract on Fusarium Wilt Control in Strawberry and Antifungal Properties of Secondary Metabolites. Hortic. J. 2020, 89, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Stanojević, D.; Čomić, L.; Stefanović, O.; Solujić-Sukdolak, S. In Vitro Synergistic Antibacterial Activity of Salvia officinalis L. and Some Preservatives. Arch. Biol. Sci. 2010, 62, 167–174. [Google Scholar] [CrossRef]
- Ince, A.E.; Şahin, S.; Şümnü, S.G. Extraction of Phenolic Compounds from Melissa Using Microwave and Ultrasound. Turk. J. Agric. For. 2013, 37, 69–75. [Google Scholar] [CrossRef]
- Ordaz, J.J.; Hernández, J.M.; Ramírez-Godínez, J.; Castañeda-Ovando, A.; González-Olivares, L.G.; Contreras-López, E. Bioactive Compounds in Aqueous Extracts of Lemon Balm (Melissa officinalis) Cultivated in Mexico. Arch. Latinoam. Nutr. 2018, 68, 268–279. [Google Scholar]
- Ehsani, A.; Alizadeh, O.; Hashemi, M.; Afshari, A.; Aminzare, M.; Dvm, M.H. Phytochemical, Antioxidant and Antibacterial Properties of Melissa officinalis and Dracocephalum Moldavica Essential Oils. Vet. Res. Forum 2017, 8, 223–229. [Google Scholar] [PubMed]
- Espinosa-Pardo, F.A.; Nakajima, V.M.; Macedo, G.A.; Macedo, J.A.; Martínez, J. Extraction of Phenolic Compounds from Dry and Fermented Orange Pomace Using Supercritical CO2 and Cosolvents. Food Bioprod. Process. 2017, 101, 1–10. [Google Scholar] [CrossRef]
- Fornari, T.; Vicente, G.; Vázquez, E.; García-Risco, M.R.; Reglero, G. Isolation of Essential Oil from Different Plants and Herbs by Supercritical Fluid Extraction. J. Chromatogr. A 2012, 1250, 34–48. [Google Scholar] [CrossRef] [Green Version]
- Capuzzo, A.; Maffei, M.E.; Occhipinti, A. Supercritical Fluid Extraction of Plant Flavors and Fragrances. Molecules 2013, 18, 7194–7238. [Google Scholar] [CrossRef] [Green Version]
- Miron, T.L.; Herrero, M.; Ibáñez, E. Enrichment of Antioxidant Compounds from Lemon Balm (Melissa officinalis) by Pressurized Liquid Extraction and Enzyme-Assisted Extraction. J. Chromatogr. A 2013, 1288, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Sánchez, A.; Barrajón-Catalán, E.; Herranz-López, M.; Castillo, J.; Micol, V. Lemon Balm Extract (Melissa officinalis, L.) Promotes Melanogenesis and Prevents UVB-Induced Oxidative Stress and DNA Damage in a Skin Cell Model. J. Dermatol. Sci. 2016, 84, 169–177. [Google Scholar] [CrossRef]
- Aziz, A.; Al-Askar, A. In Vitro Antifungal Activity Of Three Saudi Plant Extracts Against Some Phytopathogenic Fungi. J. PLANT Prot. Res. 2012, 52, 458–462. [Google Scholar] [CrossRef]
- Manssouri, M.; Znini, M.; El Harrak, A.; Majidi, L. Antifungal Activity of Essential Oil from the Fruits of Ammodaucus leucotrichus Coss. & Dur., in Liquid and Vapour Phase against Postharvest Phytopathogenic Fungi in Apples. J. Appl. Pharm. Sci. 2016, 6, 131–136. [Google Scholar] [CrossRef] [Green Version]
- De Lira Mota, K.S.; De Oliveira Pereira, F.; Araújo De Oliveira, W.; Lima, I.O.; De, E.; Lima, O. Molecules Antifungal Activity of Thymus vulgaris L. Essential Oil and Its Constituent Phytochemicals against Rhizopus Oryzae: Interaction with Ergosterol. Molecules 2012, 17, 14418–14433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Zhang, J.; Kong, W.; Zhao, G.; Yang, M. Mechanisms of Antifungal and Anti-Aflatoxigenic Properties of Essential Oil Derived from Turmeric (Curcuma longa L.) on Aspergillus flavus. Food Chem. 2017, 220, 1–8. [Google Scholar] [CrossRef]
- Charcosset, J.-Y.; Chauvet, E. Effect of Culture Conditions on Ergosterol as an Indicator of Biomass in the Aquatic Hyphomycetes. Appl. Environ. Microbiol. 2001, 67, 2051–2055. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto-Ribeiro, M.M.G.; Grespan, R.; Kohiyama, C.Y.; Ferreira, F.D.; Mossini, S.A.G.; Silva, E.L.; De Abreu Filho, B.A.; Mikcha, J.M.G.; Machinski, M. Effect of Zingiber Officinale Essential Oil on Fusarium verticillioides and Fumonisin Production. Food Chem. 2013, 141, 3147–3152. [Google Scholar] [CrossRef] [Green Version]
- Caloni, F.; Fossati, P.; Anadón, A.; Bertero, A. Beauvericin: The Beauty and the Beast. Environ. Toxicol. Pharmacol. 2020, 75, 103349. [Google Scholar] [CrossRef]
- Barral, B.; Chillet, M.; Doizy, A.; Grassi, M.; Ragot, L.; Léchaudel, M.; Durand, N.; Rose, L.J.; Viljoen, A.; Schorr-Galindo, S. Diversity and Toxigenicity of Fungi That Cause Pineapple Fruitlet Core Rot. Toxins 2020, 12, 339. [Google Scholar] [CrossRef]
- Sumalan, R.-M.; Alexa, E.; Poiana, M.-A. Assessment of Inhibitory Potential of Essential Oils on Natural Mycoflora and Fusarium Mycotoxins Production in Wheat. Chem. Cent. J. 2013, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Bryła, M.; Ksieniewicz-Wózniak, E.; Waśkiewicz, A.; Yoshinari, T.; Szymczyk, K.; Podolska, G.; Gwiazdowski, R.; Kubiak, K. Transformations of Selected Fusarium Toxins and Their Modified Forms during Malt Loaf Production. Toxins 2020, 12, 385. [Google Scholar] [CrossRef]
- Freire, L.; Sant’Ana, A.S. Modified Mycotoxins: An Updated Review on Their Formation, Detection, Occurrence, and Toxic Effects. Food Chem. Toxicol. 2018, 111, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Perczak, A.; Gwiazdowska, D.; Marchwińska, K.; Juś, K.; Gwiazdowski, R.; Waśkiewicz, A. Antifungal Activity of Selected Essential Oils against Fusarium culmorum and F. graminearum and Their Secondary Metabolites in Wheat Seeds. Arch. Microbiol. 2019, 201, 1085–1097. [Google Scholar] [CrossRef] [Green Version]
- Góral, T.; Wiśniewska, H.; Ochodzki, P.; Nielsen, L.K.; Walentyn-Góral, D.; Stępień, Ł. Relationship between Fusarium Head Blight, Kernel Damage, Concentration of Fusarium Biomass, and Fusarium Toxins in Grain of Winter Wheat Inoculated with Fusarium culmorum. Toxins 2019, 11, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perincherry, L.; Witaszak, N.; Urbaniak, M.; Waśkiewicz, A.; Stępień, Ł. Effects of Secondary Metabolites from Pea on Fusarium Growth and Mycotoxin Biosynthesis. J. Fungi 2021, 7, 1004. [Google Scholar] [CrossRef] [PubMed]
- Salhi, N.; Mohammed Saghir, S.A.; Terzi, V.; Brahmi, I.; Ghedairi, N.; Bissati, S. Antifungal Activity of Aqueous Extracts of Some Dominant Algerian Medicinal Plants. Biomed Res. Int. 2017, 7526291. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variants | Extract Concentrations (%) | Mycelium Growth Inhibition (%) | |
|---|---|---|---|
| F. proliferatum | F. culmorum | ||
| 60NE | 1 | 13.0 ± 0.1 ab | 2.5 ± 0.0 ab |
| 2.5 | 35.0 ± 1.8 abc | 30.7 ± 1.9 abcd | |
| 5 | 86.7 ± 0.1 de | 48.2 ± 2.1 bcde | |
| 7.5 | 96.1 ± 0.2 e | 100.0 ± 0.0 f | |
| 10 | 100.0 ± 0.0 e | 100.0 ± 0.0 f | |
| 40NE | 1 | 7.4 ± 0.0 ab | 4.1 ± 0.9 ab |
| 2.5 | 48.9 ± 0.2 bcd | 22.3 ± 1.5 abc | |
| 5 | 71.9 ± 0.5 cde | 59.7 ± 1.1 cdef | |
| 7.5 | 100.0 ± 0.0 e | 96.1 ± 0.5 ef | |
| 10 | 100.0 ± 0.0 e | 100.0 ± 0.0 f | |
| 40E | 1 | 9.4 ± 0.0 ab | 1.7 ± 1.3 a |
| 2.5 | 24.0 ± 0.2 ab | 17.2 ± 1.0 abc | |
| 5 | 72.7 ± 1.2 cde | 90.0 ± 0.4 ef | |
| 7.5 | 100.0 ± 0.0 e | 96.8 ± 0.3 ef | |
| 10 | 100.0 ± 0.0 e | 100.0 ± 0.0 f | |
| 60E | 1 | 4.6 ± 0.0 ab | 0.7 ± 1.8 a |
| 2.5 | 33.4 ± 0.6 abc | 54.5 ± 0.4 cdef | |
| 5 | 81.9 ± 0.5 cde | 80.5 ± 0.1 def | |
| 7.5 | 100.0 ± 0.0 e | 94.3 ± 0.2 ef | |
| 10 | 100.0 ± 0.0 e | 100.0 ± 0.0 f | |
| Variants | Extract Concentrations [%] | ERG Concentration [µg/g] and Percentage of Reduction [%] | |||
|---|---|---|---|---|---|
| F. proliferatum | F. culmorum | ||||
| [µg/g] | [%] | [µg/g] | [%] | ||
| Control (without extract) | 178.0 ± 23.4 g | - | 55.7 ± 5.3 i | - | |
| 60NE | 1 | 74.3 ± 6.4 e | 58.3 | 28.5 ± 3.9 fg | 48.8 |
| 2.5 | 31.7 ± 3.8 bcd | 82.2 | 14.2 ± 3.0 bcd | 74.4 | |
| 5 | 23.7 ± 3.9 abcd | 86.7 | 8.6 ± 0.4 abcd | 84.6 | |
| 7.5 | 21.09 ± 2.0 abc | 88.2 | 7.5 ± 0.8 abc | 86.6 | |
| 10 | 12.2 ± 3.3 abc | 93.2 | 7.6 ± 0.7 abcd | 86.3 | |
| 40NE | 1 | 141.4 ± 9.5 f | 20.6 | 45.6 ± 4.4 h | 18.0 |
| 2.5 | 47.6 ± 8.8 d | 73.3 | 28.7 ± 2.8 fg | 48.5 | |
| 5 | 35.3 ± 4.8 cd | 80.2 | 26.5 ± 1.4 ef | 52.4 | |
| 7.5 | 15.3 ± 4.3 abc | 91.4 | 16.1 ± 2.5 cd | 71.1 | |
| 10 | 6.7 ± 0.7 a | 96.2 | 0.7 ± 0.1 a | 98.8 | |
| 40E | 1 | 144.0 ± 6.8 f | 19.1 | 36.5 ± 5.0 gh | 34.4 |
| 2.5 | 26.3 ± 5.4 abcd | 85.2 | 27.9 ± 3.3 fg | 49.9 | |
| 5 | 21.1 ± 3.3 abc | 88.2 | 6.8 ± 0.7 abc | 87.8 | |
| 7.5 | 13.0 ± 3.4 abc | 92.7 | 4.8 ± 0.3 ab | 91.4 | |
| 10 | 8.9 ± 1.6 ab | 95.0 | 3.4 ± 0.4 a | 94.0 | |
| 60E | 1 | 47.7 ± 5.1 d | 73.2 | 33.5 ± 3.6 fg | 39.8 |
| 2.5 | 32.8 ± 3.4 bcd | 81.6 | 17.3 ± 4.2 de | 68.9 | |
| 5 | 11.7 ± 2.3 abc | 93.4 | 6.41 ± 1.0 abc | 88.5 | |
| 7.5 | 9.4 ± 2.0 ab | 94.7 | 3.5 ± 0.6 a | 93.7 | |
| 10 | 2.9 ± 0.5 a | 98.4 | 2.5± 0.5 a | 95.5 | |
| Variants | Extract Concentrations (%) | Mycotoxins (µg/g) | |||
|---|---|---|---|---|---|
| FB1 | FB2 | FB3 | BEA | ||
| Control (without extract) | 4.3 ± 0.1 j | 0.4 ± 0.1 h | 0.2 ± 0.0 f | 36.4 ± 0.6 l | |
| 60NE | 1 | 1.3 ± 0.0 f | 0.1 ± 0.0 bcde | 0.1 ± 0.0 e | 13.4 ± 0.6 k |
| 2.5 | 1.3 ± 0.0 f | 0.1 ± 0.0 bcde | 0.03 ± 0.0 abcd | 11.2 ± 0.8 j | |
| 5 | 0.8 ± 0.1 de | 0.1 ± 0.0 bcde | 0.02 ± 0.0 abc | 6.1 ± 0.8 h | |
| 7.5 | 0.4 ± 0.0 bc | 0.03 ± 0.0 ab | 0.01 ± 0.0 ab | 2.8 ± 0.1 def | |
| 10 | 0.1 ± 0.0 a | 0.02 ± 0.0 a | n.d.a | 0.9 ± 0.0 a | |
| 40NE | 1 | 1.2 ± 0.0 f | 0.3 ± 0.0 g | 0.1 ± 0.0 e | 4.9 ± 0.1 g |
| 2.5 | 0.4 ± 0.0 bc | 0.2 ± 0.0 f | 0.1 ± 0.0 e | 3.5 ± 0.1 f | |
| 5 | 0.2 ± 0.0 ab | 0.2 ± 0.0 f | 0.02 ± 0.0 abc | 2.2 ± 0.1 de | |
| 7.5 | 0.2 ± 0.0 ab | 0.1 ± 0.0 bcde | n.d.a | 1.9 ± 0.1 bcd | |
| 10 | 0.1 ± 0.0 a | 0.04 ± 0.0 abc | n.d.a | 2.0 ± 0.1 bcd | |
| 40E | 1 | 3.6 ± 0.2 i | 0.1 ± 0.0 bcde | 0.1 ± 0.0 e | 8.0 ± 0.1 i |
| 2.5 | 3.8 ± 0.2 i | 0.1 ± 0.0 bcde fg | 0.04 ± 0.0 bcd | 5.8 ± 0.1 gh | |
| 5 | 2.4 ± 0.1 g | 0.1 ± 0.0 bcde | 0.03 ± 0.0 abcd | 5.8 ± 0.1 gh | |
| 7.5 | 0.6 ± 0.0 cd | 0.04 ± 0.0 abc | 0.01 ± 0.0 ab | 1.1 ± 0.1 ab | |
| 10 | 0.1 ± 0.0 a | 0.03 ± 0.0 ab | n.d.a | 1.0 ± 0.1 ab | |
| 60E | 1 | 3.6 ± 0.2 i | 0.1 ± 0.0 bcde | 0.1 ± 0.0 e | 5.6 ± 0.3 gh |
| 2.5 | 3.1 ± 0.1 h | 0.2 ± 0.0 f | 0.05 ± 0.0 cd | 5.1 ± 0.1 g | |
| 5 | 1.1 ± 0.1 ef | 0.1 ± 0.0 bcde | 0.03 ± 0.0 abcd | 3.2 ± 0.2 ef | |
| 7.5 | 0.2 ± 0.0 abc | 0.04 ± 0.0 abc | 0.01 ± 0.0 ab | 2.2 ± 0.1 cde | |
| 10 | 0.2 ± 0.0 abc | 0.03 ± 0.0 ab | n.d.a | 1.2 ± 0.1 abc | |
| Variants | Extract Concentrations (%) | Mycotoxins [µg/g] | ||||
|---|---|---|---|---|---|---|
| DON | 3- + 15- AcDON | ZEN | ZEN-14S | β-ZOL | ||
| Control (without extract) | 29.8 ± 0.5 j | 40.3 ± 0.7 h | 250.6 ± 12.1 k | 672.4 ± 16.7 h | 0.5 ± 0.0 f | |
| 60NE | 1 | 13.3 ± 0.2 h | 37.2 ± 0.7 h | 146.4 ± 0.3 i | 166.5 ± 1.0 de | 0.5 ± 0.0 f |
| 2.5 | 7.2 ± 0.3 d | 12.1 ± 0.1 d | 38.6 ± 1.4 de | 56.1 ± 3.3 bc | 0.2 ± 0.0 d | |
| 5 | 2.5 ± 0.1 b | 7.0 ± 5.3 c | 20.2 ± 0.9 bcd | 9.9 ± 7.3 a | 0.1 ± 0.0 bc | |
| 7.5 | 1.0 ± 0.1 a | 7.0 ± 0.3 c | 4.1 ± 0.0 abc | 5.1 ± 0.1 a | 0.03 ± 0.0 ab | |
| 10 | 0.4 ± 0.0 a | 0.2 ± 0.0 a | 0.1 ± 0.0 a | 1.3 ± 0.1 a | 0.01 ± 0.0 a | |
| 40NE | 1 | 11.7 ± 0.5 g | 25.4 ± 0.5 f g | 168.9 ± 20.8 j | 193.9 ± 3.3 e | 0.4 ± 0.1 f |
| 2.5 | 5.2 ± 0.2 c | 16.9 ± 0.4 e | 89.9 ± 5.0 g | 80.5 ± 6.0 bc | 0.2 ± 0.0 cd | |
| 5 | 0.9 ± 0.6 a | 4.8 ± 0.1 bc | 16.6 ± 0.2 abc | 73.5 ± 5.9 bc | 0.1 ± 0.0 bc | |
| 7.5 | 0.5 ± 0.0 a | 0.9 ± 0.0 ab | 1.1 ± 0.0 ab | 41.5 ± 0.9 ab | 0.04 ± 0.0 ab | |
| 10 | 0.3 ± 0.0 a | 0.1 ± 0.0 a | 1.5 ± 0.2 ab | 1.2 ± 0.1 a | n.d. a | |
| 40E | 1 | 9.2 ± 0.1 e | 28.1 ± 0.8 g | 110.5 ± 2.1 h | 679.4 ± 11.7 h | 0.5 ± 0.0 f |
| 2.5 | 9.2 ± 0.3 e | 21.4 ± 0.6 f | 67.0 ± 1.6 f | 328.4 ± 25.1 g | 0.2 ± 0.0 cd | |
| 5 | 5.1 ± 0.1 c | 5.0 ± 0.1 bc | 4.3 ± 0.0 abc | 149.4 ± 12.5 d | 0.1 ± 0.0 cd | |
| 7.5 | 1.0 ± 0.0 a | 0.2 ± 0.0 a | 3.5 ± 0.1 abc | 11.4 ± 0.8 a | 0.02 ± 0.0 ab | |
| 10 | 0.4 ± 0.0 a | n.d. a | 0.1 ± 0.0 a | 5.6 ± 0.2 a | 0.01 ± 0.0 a | |
| 60E | 1 | 16.1 ± 0.5 i | 26.7 ± 0.3 g | 52.4 ± 0.2 ef | 266.2 ± 43.0 f | 0.3 ± 0.0 e |
| 2.5 | 10.5 ± 0.7 f | 14.9 ± 0.3 de | 22.6 ± 1.3 cd | 209.9 ± 2.7 e | 0.1 ± 0.0 cd | |
| 5 | 7.2 ± 0.2 d | 3.0 ± 0.1 abc | 6.0 ± 0.0 abc | 92.1 ± 4.1 c | 0.2 ± 0.0 d | |
| 7.5 | 2.1 ± 0.1 b | 0.3 ± 0.0 a | 4.4 ± 0.7 abc | 40.7 ± 0.5 ab | 0.05 ± 0.0 ab | |
| 10 | 0.6 ± 0.0 a | 0.1 ± 0.0 a | 4.1 ± 0.1 abc | 8.1 ± 0.4 a | n.d. a | |
| Analyte | Ion Mass [m/z] | Retention Time [min] | LOQ * [ng/g] | LOD * [ng/g] |
|---|---|---|---|---|
| FB1 | 722.837 (M + H)+ | 8.41 | 50 | 15 |
| FB2 | 706.837 (M + H)+ | 10.56 | 50 | 15 |
| FB3 | 706.837 (M + H)+ | 9.45 | 50 | 15 |
| DON | 297.327 (M + H)+ | 3.21 | 80 | 24 |
| 3- + 15-AcDON | 339.357 (M + H)+ | 5.80 | 20 | 6 |
| ZEN | 317.352 (M − H)− | 9.44 | 5 | 1.5 |
| ZEN-14S | 397.081 (M − H)− | 7.24 | 10 | 3 |
| Β-ZOL | 319.372 (M − H)− | 8.31 | 4 | 1.2 |
| BEA | 801.983 (M + NH4)+ | 17.86 | 2 | 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uwineza, P.A.; Urbaniak, M.; Bryła, M.; Stępień, Ł.; Modrzewska, M.; Waśkiewicz, A. In Vitro Effects of Lemon Balm Extracts in Reducing the Growth and Mycotoxins Biosynthesis of Fusarium culmorum and F. proliferatum. Toxins 2022, 14, 355. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050355
Uwineza PA, Urbaniak M, Bryła M, Stępień Ł, Modrzewska M, Waśkiewicz A. In Vitro Effects of Lemon Balm Extracts in Reducing the Growth and Mycotoxins Biosynthesis of Fusarium culmorum and F. proliferatum. Toxins. 2022; 14(5):355. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050355
Chicago/Turabian StyleUwineza, Pascaline Aimee, Monika Urbaniak, Marcin Bryła, Łukasz Stępień, Marta Modrzewska, and Agnieszka Waśkiewicz. 2022. "In Vitro Effects of Lemon Balm Extracts in Reducing the Growth and Mycotoxins Biosynthesis of Fusarium culmorum and F. proliferatum" Toxins 14, no. 5: 355. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050355