Spatio-Temporal Monitoring of Benthic Anatoxin-a-Producing Tychonema sp. in the River Lech, Germany



Abstract
:1. Introduction
2. Results
2.1. Occurrence and Distribution of Tychonema in the River Lech
2.2. Molecular Analyses
2.2.1. Illumina MiSeq Sequencing
2.2.2. Detection of Toxin Genes
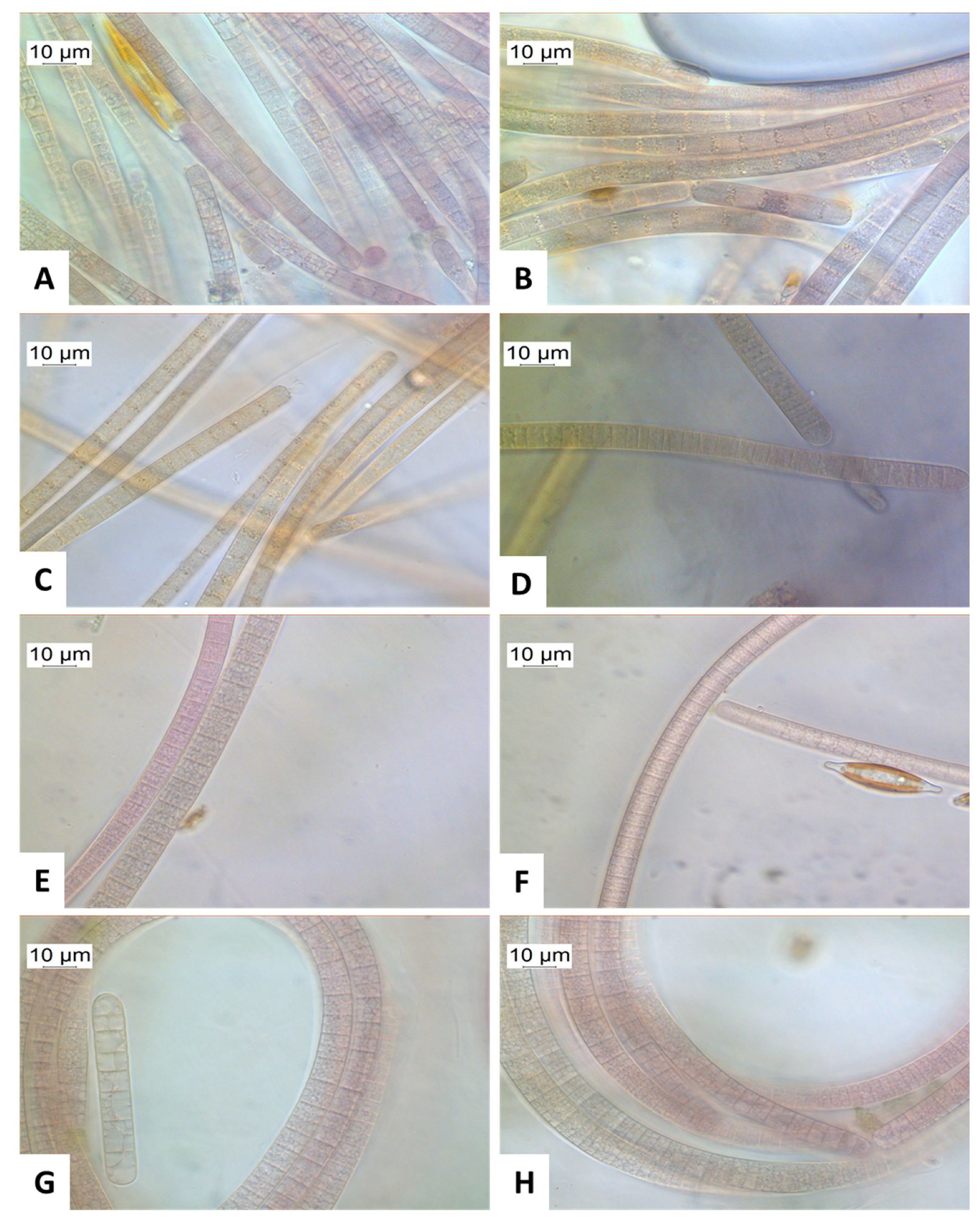
2.3. Macroscopic and Microscopic Appearance of Tychonema
2.4. Toxin Analyses
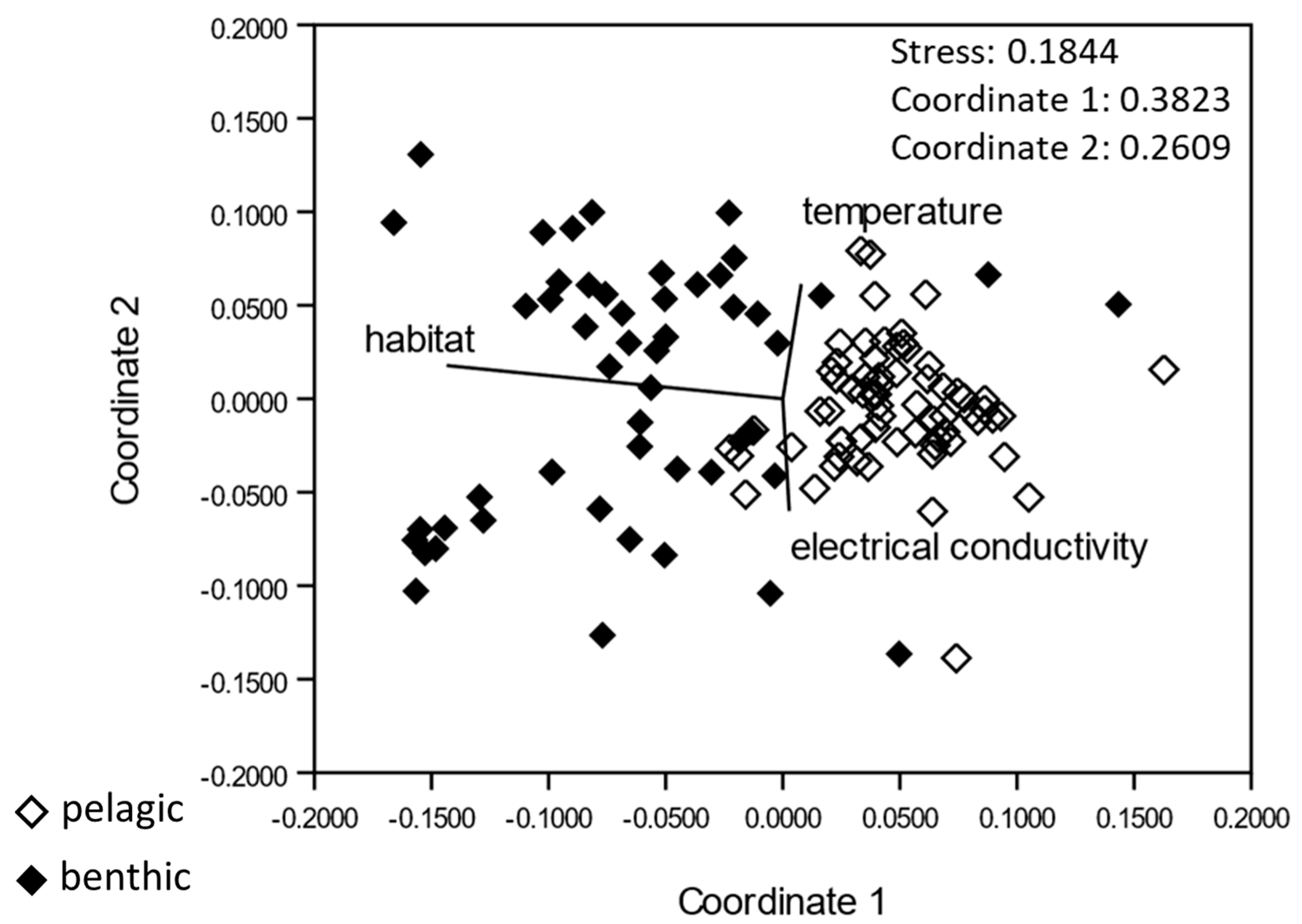
2.5. Habitat Characterization
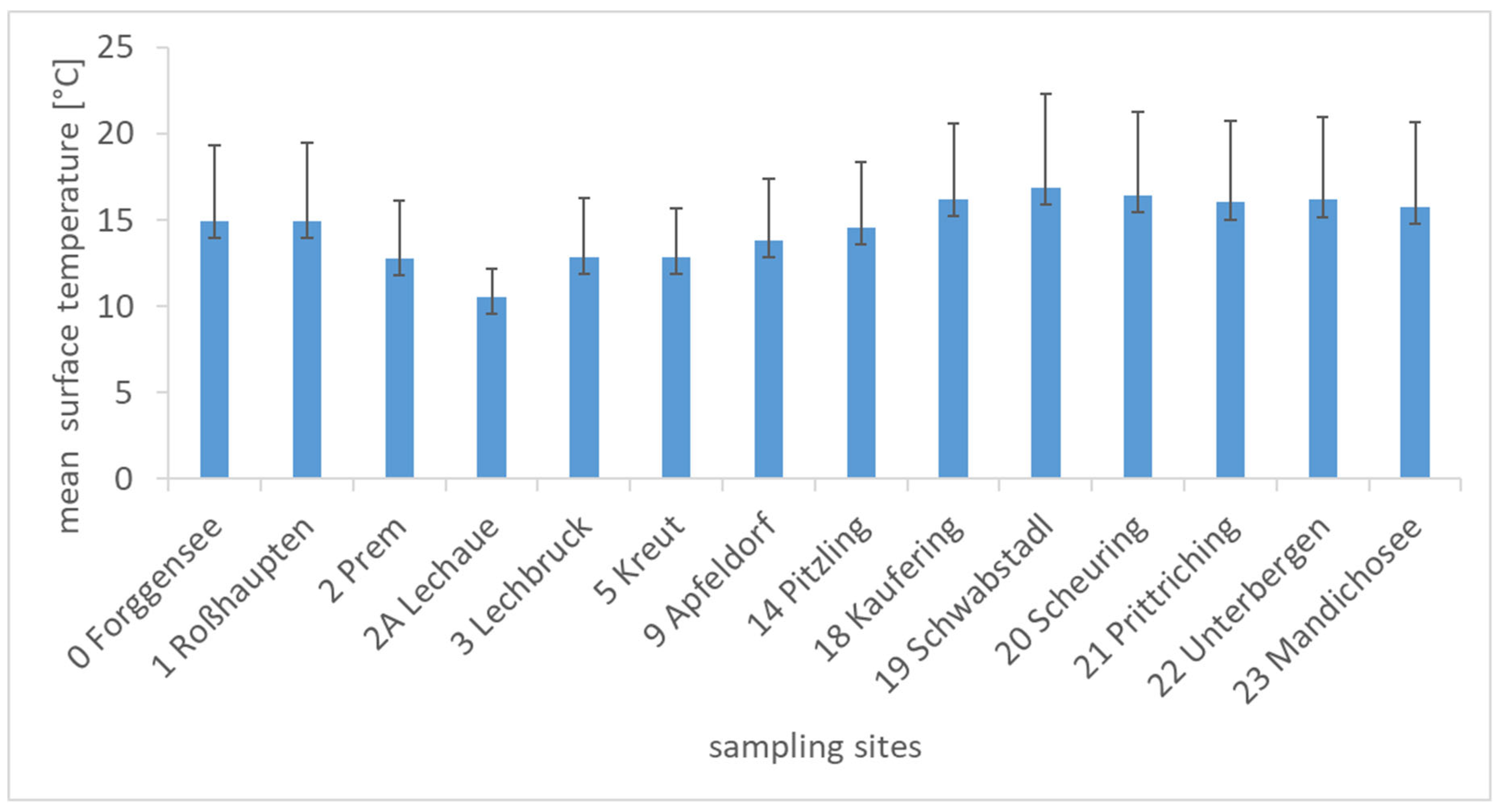
2.5.1. Hydro-Physical Parameters
2.5.2. Substrate
3. Discussion
4. Conclusions
5. Materials and Methods
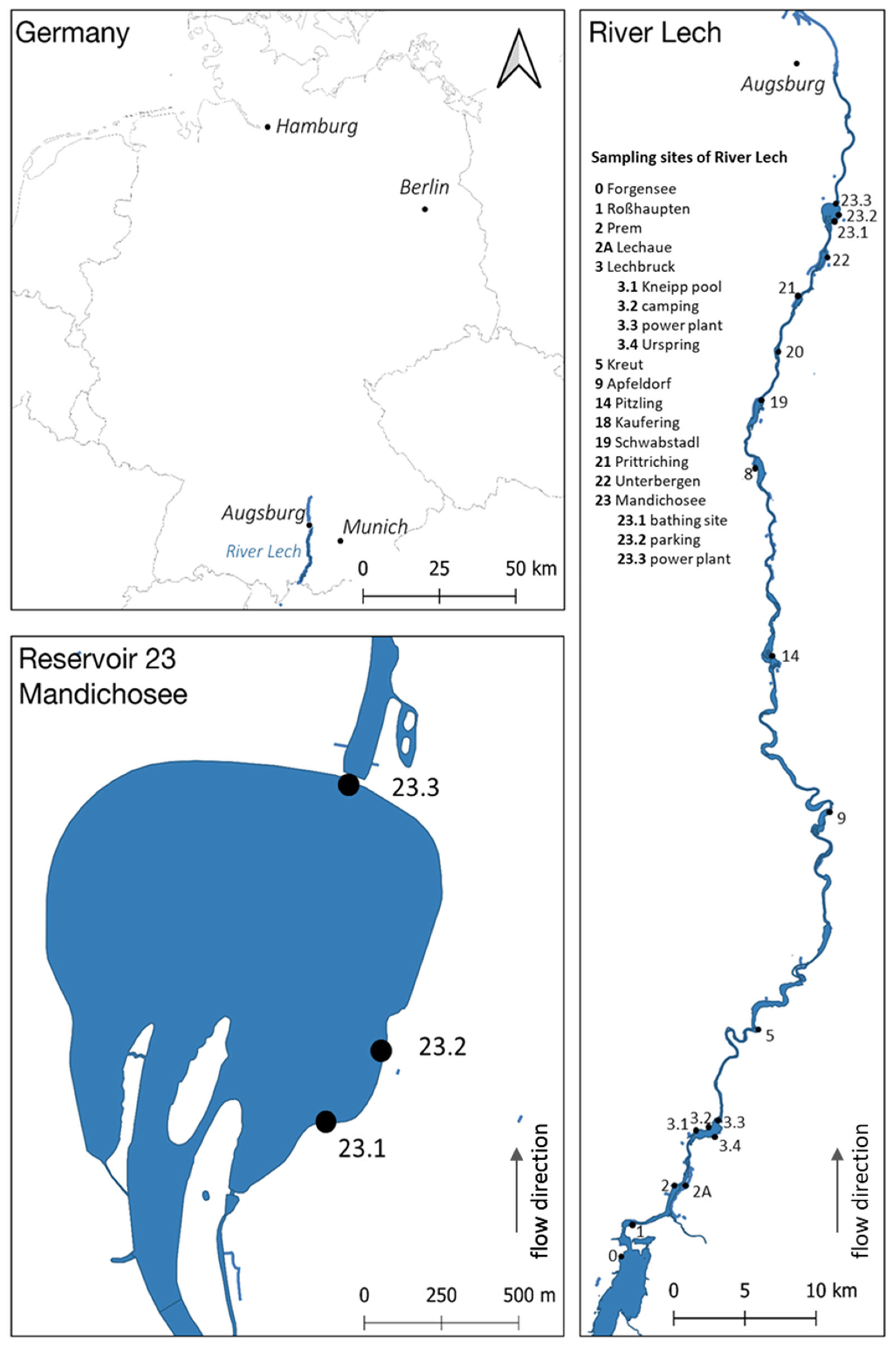
5.1. Study Site
5.2. Sampling and Post-Processing of the Samples
5.3. Molecular Detection
5.3.1. DNA Extraction and Purification
5.3.2. Polymerase Chain Reaction
5.3.3. Sequencing
5.4. Morphological Examinations
5.4.1. Macroscopy
5.4.2. Microscopy
5.5. Toxin Analyses
5.6. Habitat Specification
5.6.1. In Situ Measurements of Hydro-Physical Variables
5.6.2. Substrate Characterization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carmichael, W.W. (Ed.) Freshwater blue-green algae (cyanobacteria) toxins—A review. In The Water Environment; Springer: Boston, MA, USA, 1981; pp. 1–13. [Google Scholar]
- Vasconcelos, V. Cyanobacteria toxins: Diversity and ecological effects. Limnetica 2001, 20, 45–58. [Google Scholar] [CrossRef]
- Sivonen, K. Cyanobacterial toxins. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Elsevier: Oxford, UK, 2009; pp. 290–307. [Google Scholar]
- Leão, P.N.; Niclas, E.; Antunes, A.; Gerwick, W.H.; Vasconcelos, V. The chemical ecology of cyanobacteria. Nat. Prod. Rep. 2012, 29, 372–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitois, S.; Jackson, M.H.; Wood, B.J.B. Problems associated with the presence of cyanobacteria in recreational and drinking waters. Int. J. Environ. Health Res. 2000, 10, 203–218. [Google Scholar] [CrossRef]
- Falconer, I.R. Toxic cyanobacterial bloom problems in Australian waters: Risks and impacts on human health. Phycologia 2001, 40, 228–233. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Scott, J.T.; Marcarelli, A.M. Cyanobacteria in freshwater benthic environments. In Ecology of cyanobacteria II; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; Volume 1, pp. 271–289. [Google Scholar]
- Sukenik, A.; Quesada, A.; Salmaso, N. Global expansion of toxic and non-toxic cyanobacteria: Effect on ecosystem functioning. Biodivers. Conserv. 2015, 24, 889–908. [Google Scholar] [CrossRef]
- Mur, L.R.; Skulberg, O.M.; Utkilen, H. Cyanobacteria in the environment. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 1st ed.; Chorus, I., Bartram, J., Eds.; Taylor and Francis: Abingdon, UK, 1999. [Google Scholar]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.F. A review of current knowledge on toxic benthic freshwater cyanobacteria–ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Elliott, J.A. Is the future blue-green? A review of the current model predictions of how climate change could affect pelagic freshwater cyanobacteria. Water Res. 2012, 46, 1364–1371. [Google Scholar] [CrossRef] [Green Version]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Gobler, C.J. Climate change and harmful algal blooms: Insights and perspective. Harmful algae 2020, 91, 101731. [Google Scholar] [CrossRef]
- Edwards, C.; Beattie, K.A.; Scrimgeour, C.M.; Codd, G.A. Identification of anatoxin-a in benthic cyanobacteria (blue-green algae) and in associated dog poisonings at Loch Insh, Scotland. Toxicon 1992, 30, 1165–1175. [Google Scholar] [CrossRef]
- Gugger, M.; Lenoir, S.; Berger, C.; Ledreux, A.; Druart, J.C.; Humbert, J.F.; Guette, C.; Bernard, C. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 2005, 45, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Rasmussen, J.P.; Holland, P.T.; Campbell, R.; Crowe, A.L. First report of the cyanotoxin anatoxin-a from Aphanizomenon issatschenkoi (cyanobacteria). J. Phycol. 2007, 43, 356–365. [Google Scholar] [CrossRef]
- Faassen, E.; Harkema, L.; Begeman, L.; Lurling, M. First report of (homo)anatoxin-a and dog neurotoxicosis after ingestion of benthic cyanobacteria in The Netherlands. Toxicon 2012, 60, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Bauer, F.; Fastner, J.; Bartha-Dima, B.; Breuer, W.; Falkenau, A.; Mayer, C.; Raeder, U. Mass occurrence of anatoxin-a-and dihydroanatoxin-a-producing Tychonema sp. in mesotrophic reservoir Mandichosee (River Lech, Germany) as a cause of neurotoxicosis in dogs. Toxins 2020, 12, 726. [Google Scholar] [CrossRef]
- Urrutia-Cordero, P.; Zhang, H.; Chaguaceda, F.; Geng, H.; Hansson, L.A. Climate warming and heat waves alter harmful cyanobacterial blooms along the benthic–pelagic interface. Ecology 2020, 101, e03025. [Google Scholar] [CrossRef] [Green Version]
- Méjean, A.; Mann, S.; Maldiney, T.; Vassiliadis, G.; Lequin, O.; Ploux, O. Evidence that biosynthesis of the neurotoxic alkaloids anatoxin-a and homoanatoxin-a in the cyanobacterium Oscillatoria PCC 6506 occurs on a modular polyketide synthase initiated by l-proline. J. Am. Chem. Soc. 2009, 131, 7512–7513. [Google Scholar] [CrossRef]
- Wood, S.A.; Biessy, L.; Puddick, J. Anatoxins are consistently released into the water of streams with Microcoleus autumnalis-dominated (cyanobacteria) proliferations. Harmful Algae 2018, 80, 88–95. [Google Scholar] [CrossRef]
- Lugomela, C.; Pratap, H.B.; Mgaya, Y.D. Cyanobacteria blooms—a possible cause of mass mortality of Lesser Flamingos in Lake Manyara and Lake Big Momela, Tanzania. Harmful Algae 2006, 5, 534–541. [Google Scholar] [CrossRef]
- Mez, K.; Beattie, K.A.; Codd, G.A.; Hanselmann, K.; Hauser, B.; Naegeli, H.; Preisig, H.R. Identification of a microcystin in benthic cyanobacteria linked to cattle deaths on alpine pastures in Switzerland. Eur. J. Phycol. 1997, 32, 111–117. [Google Scholar] [CrossRef]
- Fastner, J.; Beulker, C.; Geiser, B.; Hoffmann, A.; Kröger, R.; Teske, K.; Hoppe, J.; Mundhenk, L.; Neurath, H.; Sagebiel, D.; et al. Fatal neurotoxicosis in dogs associated with tychoplanktic, anatoxin-a producing Tychonema sp. in mesotrophic Lake Tegel, Berlin. Toxins 2018, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komarék, J.; Anagnostidis, K. Cyanoprokaryota: Oscillatoriales. In Freshwater Flora of Central Europe, 1st ed.; Büdel, B., Gärtner, G., Krienitz, L., Schlagerl, M., Eds.; Spektrum: Heidelberg, Germany, 2008; Volume 2. [Google Scholar]
- Conklin, K.Y.; Stancheva, R.; Otten, T.G.; Fadness, R.; Boyer, G.L.; Read, R.; Zhang, X.; Sheath, R.G. Molecular and morphological characterization of a novel dihydroanatoxin-a producing Microcoleus species (cyanobacteria) from the Russian River, California, USA. Harmful Algae 2020, 93, 101767. [Google Scholar] [CrossRef] [PubMed]
- Puddick, J.; van Ginkel, R.; Page, C.D.; Murray, J.S.; Greenhough, H.E.; Bowater, J.; Selwood, A.I.; Wood, S.A.; Prinsep, M.R.; Truman, P. Acute toxicity of dihydroanatoxin-a from Microcoleus autumnalis in comparison to anatoxin-a. Chemosphere 2020, 263, 127937. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, Y.; Mohammadipanah, F.; Te, S.H.; You, L.; Gin, K.Y.H. Biodiversity, phylogeny and toxin production profile of cyanobacterial strains isolated from Lake Latyan in Iran. Harmful Algae 2021, 106, 102054. [Google Scholar] [CrossRef]
- Anagnostidis, K. Geitlerinema, a new genus of oscillatorialean cyanophytes. Plant Syst. Evol. 1989, 164, 33–46. [Google Scholar] [CrossRef]
- Rangel, M.; Brunetti, R.L.; Garcia, A.N.; Cambui, C.C.N.; Conserva, G.A.A.; Neves, A.C.; Sant’Anna, E.L.; Carvalho, L.R. Acute effects of three Geitlerinema spp. (cyanobacteria) extracts administrated in mice: Symptoms and histopathological aspects. Phytochem. Rev. 2013, 12, 543–553. [Google Scholar] [CrossRef]
- Strunecký, O.; Elster, J.; Komárek, J. Taxonomic revision of the freshwater cyanobacterium “Phormidium” murrayi = Wilmottia murrayi. Fottea 2011, 11, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Machado-De-Lima, N.M.; Martins, M.D.; Branco, L.H. Description of a tropical new species of Wilmottia (Oscillatoriales, Cyanobacteria) and considerations about the monophyly of W. murrayi. Phytotaxa 2017, 307, 43–54. [Google Scholar] [CrossRef]
- Shams, S.; Capelli, C.; Cerasino, L.; Ballot, A.; Dietrich, D.R.; Sivonen, K.; Salmaso, N. Anatoxin-a producing Tychonema (Cyanobacteria) in European waterbodies. Water Res. 2015, 69, 68–79. [Google Scholar] [CrossRef] [Green Version]
- Pinto, F.; Tett, A.; Armanini, F.; Asnicar, F.; Boscaini, A.; Pasolli, E.; Zolfo, M.; Donati, C.; Salmaso, N.; Segata, N. Draft genome sequence of the planktic cyanobacterium Tychonema bourrellyi, isolated from alpine lentic freshwater. Genome Announc. 2017, 5, e01294-17. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J.; Komárkova, J.; Kling, H. Filamentous Cyanobacteria. In Freshwater Algae of North America; Wehr, J.D., Sheat, R.G., Eds.; Elsevier: Waltham, MA, USA, 2003; pp. 117–196. [Google Scholar]
- Linne von Berg, K.-H.; Hoef-Emden, K.; Birger, M.; Melkonian, M. Der Kosmos-Algenfuehrer: Die Wichtigsten Süsswasseralgen im Mikroskop, 2nd ed.; Franckh-Kosmos-Verlag: Stuttgart, Germany, 2012; Volume 1, pp. 1–366. [Google Scholar]
- Rogers, E.H.; Hunter III, E.S.; Moser, V.C.; Phillips, P.M.; Herkovits, J.; Munoz, L.; Hall, L.L.; Chernoff, N. Potential developmental toxicity of anatoxin-a, a cyanobacterial toxin. J. Appl. Toxicol. 2005, 25, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; Taylor and Francis: Abingdon, UK, 2021; pp. 1–858. [Google Scholar]
- Burford, M.A.; Carey, C.C.; Hamilton, D.P.; Huisman, J.; Paerl, H.W.; Wood, S.A.; Wulff, A. Perspective: Advancing the research agenda for improving understanding of cyanobacteria in a future of global change. Harmful Algae 2020, 91, 101601. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, A.; Bober, B.; Lechowski, Z.; Bialczyk, J. Determination of anatoxin-a stability under certain abiotic factors. Harmful Algae 2013, 28, 83–87. [Google Scholar] [CrossRef]
- Osswald, J.; Rellan, S.; Gago, A.; Vasconcelos, V. Toxicology and detection methods of the alkaloid neurotoxin produced by cyanobacteria, anatoxin-a. Environ. Int. 2007, 33, 1070–1089. [Google Scholar] [CrossRef]
- Bates, H.A.; Rapoport, H. Synthesis of anatoxin-a via intramolecular cyclization of iminium salts. J. Am. Chem. Soc. 1979, 101, 1259–1265. [Google Scholar] [CrossRef] [Green Version]
- Wonnacott, S.; Jackman, S.; Swanson, K.L.; Rapoport, H.; Albuquerque, E.X. Nicotinic pharmacology of anatoxin analogs. Side chain structure-activity relationships at neuronal nicotinic ligand binding sites. J. Pharmacol. Exp. Ther. 1991, 259, 387–391. [Google Scholar]
- Herdman, M.; Rippka, R. Cellular differentiation: Hormogonia and baeocytes. In Methods in Enzymology; Packer, L., Glazer, A.N., Eds.; Academic Press: Boston, MA, USA, 1988; Volume 167, pp. 232–242. [Google Scholar]
- Fattom, A.; Shilo, M. Hydrophobicity as an adhesion mechanism of benthic cyanobacteria. Appl. Environ. Microbiol. 1984, 47, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Harding, S.E.; Liu, Z. Cyanobacterial exopolysaccharides: Their nature and potential biotechnological applications. Biotechnol. Genet. Eng. Rev. 2001, 18, 375–404. [Google Scholar] [CrossRef]
- Sekar, R.; Venugopalan, V.P.; Satpathy, K.K.; Nair, K.V.K.; Rao, V.N.R. Laboratory studies on adhesion of microalgae to hard substrates. In Asian Pacific Phycology in the 21st Century: Prospects and Challenges, 1st ed.; Martens, K., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 109–116. [Google Scholar]
- Zwirglmaier, K.; Keiz, K.; Engel, M.; Geist, J.; Raeder, U. Seasonal and spatial patterns of microbial diversity along a trophic gradient in the interconnected lakes of the Osterseen Lake District, Bavaria. Front. Microbiol. 2015, 6, 1168. [Google Scholar] [CrossRef]
- Rantala-Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a synthetase gene cluster of the cyanobacterium Anabaena sp. strain 37 and molecular methods to detect potential producers. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef] [Green Version]
- Rantala, A.; Rajaniemi-Wacklin, P.; Lyra, C.; Lepistö, L.; Rintala, J.; Mankiewicz-Boczek, J.; Sivonen, K. Detection of microcystin-producing cyanobacteria in Finnish lakes with genus-specific microcystin synthetase gene E (mcyE) PCR and associations with environmental factors. Appl. Environ. Microbiol. 2006, 72, 6101–6110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Tebrineh, J.; Mihali, T.K.; Pomati, F.; Neilan, B.A. Detection of saxitoxin-producing cyanobacteria and Anabaena circinalis in environmental water blooms by quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 7836–7842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capelli, C.; Cerasino, L.; Boscaini, A.; Salmaso, N. Molecular tools for the quantitative evaluation of potentially toxigenic Tychonema bourrellyi (Cyanobacteria, Oscillatoriales) in large lakes. Hydrobiologia 2018, 824, 109–119. [Google Scholar] [CrossRef]
- Mizrahi-Man, O.; Davenport, E.R.; Gilad, Y. Taxonomic classification of bacterial 16S rRNA genes using short sequencing reads: Evaluation of effective study designs. PLoS ONE 2013, 8, e53608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagkouvardos, I.; Joseph, D.; Kapfhammer, M.; Giritli, S.; Horn, M.; Haller, D.; Clavel, T. IMNGS: A comprehensive open resource of processed 16S rRNA microbial profiles for ecology and diversity studies. Sci. Rep. 2016, 6, 33721. [Google Scholar] [CrossRef]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, 590–596. [Google Scholar] [CrossRef]
- Paliy, O.; Shankar, V. Application of multivariate statistical techniques in microbial ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Jens, G. Tauchstäbe zur Messung der Strömungsgeschwindigkeit und des Abflusses. Dtsch. Gewässerkundliche Mitt. 1968, 12, 90–95. [Google Scholar]








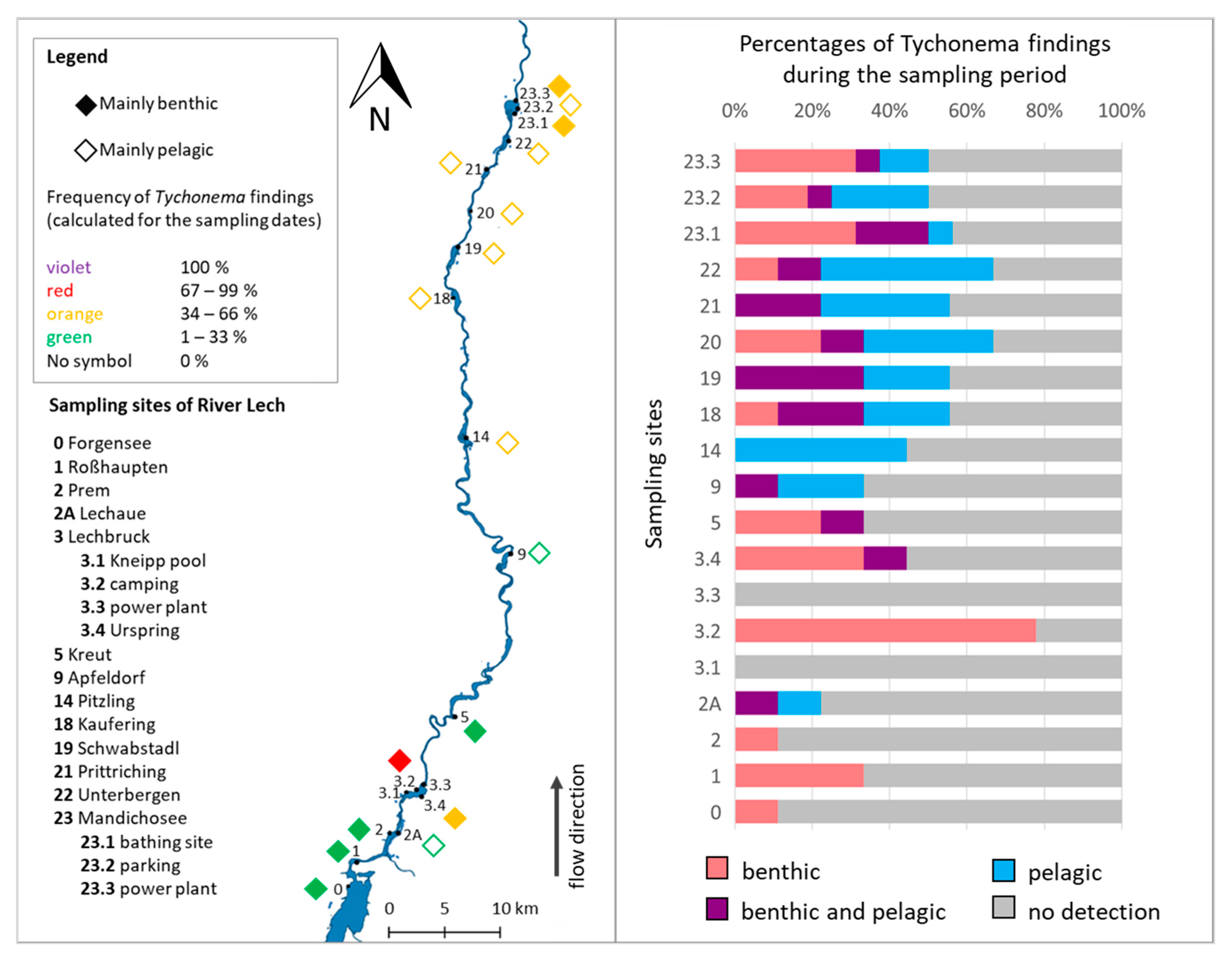{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Sampling Date | [µg/L] ATX | |
|---|---|---|---|
| 0 | Forggensee | 15 September 2020 | 0.2 |
| 0 | Forggensee | 13 October 2020 | 0.6 |
| 1 | Roßhaupten | 23 June 2020 | 9.2 |
| 1 | Roßhaupten | 21 July 2020 | 9.1 |
| 1 | Roßhaupten | 15 September 2020 | 0.2 |
| 2A | Lechaue | 22nd April 2020 | 8.7 |
| 2A | Lechaue | 5 May 2020 | 0.6 |
| 3.2 | Lechbruck-camping | 15 September 2020 | 0.6 |
| 3.2 | Lechbruck-camping | 13 October 2020 | 4.6 |
| 3.2 | Lechbruck-camping | 10 November 2020 | 0.4 |
| 3.4 | Lechbruck-Urspring | 22 April 2020 | 0.2 |
| 3.4 | Lechbruck-Urspring | 5 May 2020 | 0.1 |
| 3.4 | Lechbruck-Urspring | 15 September 2020 | 0.1 |
| 5 | Kreut | 22 April 2020 | 0.4 |
| 5 | Kreut | 5 May 2020 | 0.5 |
| 9 | Apfeldorf | 5 May 2020 | 1.8 |
| 18 | Kaufering | 5 May 2020 | 0.2 |
| 18 | Kaufering | 15 September 2020 | 0.2 |
| 18 | Kaufering | 10 November 2020 | 15.7 |
| 19 | Schwabstadl | 5 May 2020 | 0.2 |
| 19 | Schwabstadl | 13 October 2020 | 40.9 |
| 19 | Schwabstadl | 10 November 2020 | 212.5 |
| 21 | Prittriching | 5 May 2020 | 1.1 |
| 21 | Prittriching | 21 July 2020 | 11.2 |
| 22 | Unterbergen | 21 July 2020 | 0.6 |
| 22 | Unterbergen | 15 November 2020 | 13.0 |
| 22 | Unterbergen | 13 October 2020 | 89.6 |
| 23.1 | Mandichosee-bathing site | 4 August 2020 | 0.2 |
| 23.1 | Mandichosee-bathing site | 1 September 2020 | 0.2 |
| 23.1 | Mandichosee-bathing site | 15 September 2020 | 218.4 |
| 23.1 | Mandichosee WW | 29 September 2020 | 0.1 |
| 23.1 | Mandichosee-bathing site | 13 October 2020 | 144.8 |
| 23.1 | Mandichosee-bathing site | 27 October 2020 | 220.5 |
| 23.1 | Mandichosee-bathing site | 10 November 2020 | 2.3 |
| 23.2 | Mandichosee-parking | 4 August 2020 | 0.2 |
| 23.2 | Mandichosee-parking | 1 September 2020 | 0.2 |
| 23.3 | Mandichosee-power plant | 1 September 2020 | 0.2 |
| 23.3 | Mandichosee-power plant | 27 October 2020 | 0.8 |
| 23.3 | Mandichosee-power plant | 10 November 2929 | 143.8 |
| Sediment Group | Clay | Silt | Sand | Gravel | Stones | Blocks |
|---|---|---|---|---|---|---|
| Grain size [mm] | <0.0002–0.0002 | 0.0002–0.063 | 0.063–2 | 2–63 | 63–200 | >200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bauer, F.; Stix, M.; Bartha-Dima, B.; Geist, J.; Raeder, U. Spatio-Temporal Monitoring of Benthic Anatoxin-a-Producing Tychonema sp. in the River Lech, Germany. Toxins 2022, 14, 357. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050357
Bauer F, Stix M, Bartha-Dima B, Geist J, Raeder U. Spatio-Temporal Monitoring of Benthic Anatoxin-a-Producing Tychonema sp. in the River Lech, Germany. Toxins. 2022; 14(5):357. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050357
Chicago/Turabian StyleBauer, Franziska, Michael Stix, Bernadett Bartha-Dima, Juergen Geist, and Uta Raeder. 2022. "Spatio-Temporal Monitoring of Benthic Anatoxin-a-Producing Tychonema sp. in the River Lech, Germany" Toxins 14, no. 5: 357. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050357