Regulatory Networks Controlling Neurotoxin Synthesis in Clostridium botulinum and Clostridium tetani


1
Bacterial Toxins, Institut Pasteur, 75724 Paris, France
2
Department of Biomedicine, Aarhus University, 8000 Aarhus, Denmark
*
Author to whom correspondence should be addressed.
Toxins 2022, 14(6), 364; https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14060364
Submission received: 14 April 2022
/
Revised: 16 May 2022
/
Accepted: 21 May 2022
/
Published: 24 May 2022
(This article belongs to the Special Issue Toxins: Mr Hyde or Dr Jekyll?)
Abstract
:Clostridium botulinum and Clostridium tetani are Gram-positive, spore-forming, and anaerobic bacteria that produce the most potent neurotoxins, botulinum toxin (BoNT) and tetanus toxin (TeNT), responsible for flaccid and spastic paralysis, respectively. The main habitat of these toxigenic bacteria is the environment (soil, sediments, cadavers, decayed plants, intestinal content of healthy carrier animals). C. botulinum can grow and produce BoNT in food, leading to food-borne botulism, and in some circumstances, C. botulinum can colonize the intestinal tract and induce infant botulism or adult intestinal toxemia botulism. More rarely, C. botulinum colonizes wounds, whereas tetanus is always a result of wound contamination by C. tetani. The synthesis of neurotoxins is strictly regulated by complex regulatory networks. The highest levels of neurotoxins are produced at the end of the exponential growth and in the early stationary growth phase. Both microorganisms, except C. botulinum E, share an alternative sigma factor, BotR and TetR, respectively, the genes of which are located upstream of the neurotoxin genes. These factors are essential for neurotoxin gene expression. C. botulinum and C. tetani share also a two-component system (TCS) that negatively regulates neurotoxin synthesis, but each microorganism uses additional distinct sets of TCSs. Neurotoxin synthesis is interlocked with the general metabolism, and CodY, a master regulator of metabolism in Gram-positive bacteria, is involved in both clostridial species. The environmental and nutritional factors controlling neurotoxin synthesis are still poorly understood. The transition from amino acid to peptide metabolism seems to be an important factor. Moreover, a small non-coding RNA in C. tetani, and quorum-sensing systems in C. botulinum and possibly in C. tetani, also control toxin synthesis. However, both species use also distinct regulatory pathways; this reflects the adaptation of C. botulinum and C. tetani to different ecological niches.
Keywords:
Clostridium tetani; Clostridium botulinum; botulinum neurotoxin; tetanus neurotoxin; toxin gene regulation; two-component system; small RNAKey Contribution: Clostridium botulinum and Clostridium tetani produce potent neurotoxins under the control of complex regulatory networks. This review summarizes and compares the regulation of toxin synthesis in both microorganisms.
1. Introduction
Clostridium botulinum and Clostridium tetani are Gram-positive, spore-forming, and anaerobic rod-shaped bacteria that produce the most potent toxins among bacterial, animal, and plant toxins. C. tetani synthesizes the tetanus neurotoxin (TeNT), which is responsible for an often fatal spastic paralysis in humans and animals, whereas C. botulinum produces botulinum neurotoxins (BoNTs), which induce severe flaccid paralysis in vertebrates. Although TeNT and BoNTs lead to opposite clinical symptoms, they use a similar molecular mechanism of action. Indeed, TeNT and BoNTs deliver into target neurons their intracellularly active domain (light chain, Lc), which exerts a specific protease activity towards one of the three SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor) proteins: synaptobrevin or VAMP (vesicle-associated membrane protein), SNAP25 (synaptosomal-associated protein 25), and syntaxin. This leads to the inhibition of exocytosis of synaptic vesicles containing neurotransmitters. However, through their heavy chains (Hc), which recognize distinct cell surface receptors, TeNT and BoNTs undergo distinct trafficking in the host and target different neuronal cell types. Thereby, BoNTs target the motoneuron endings of the peripheral nervous system and block the release of acetylcholine, leading to a flaccid paralysis (botulism). In contrast, TeNT enters the central nervous system via retrograde transport through motoneurons and specifically inhibits the release of neurotransmitters (glycine, gamma aminobutyric acid (GABA)) in inhibitory interneurons, thus disrupting the negative control exerted by inhibitory interneurons over motoneurons, resulting in spastic paralysis (tetanus) [1,2].
2. Diversity of Clostridial Neurotoxins and Neurotoxigenic Bacterial Strains
BoNTs constitute a family of neurotoxins that share a similar structure and mode of action. They are classically divided into seven toxinotypes (A, B, C, D, E, F, and G) based on their neutralization by corresponding specific polyclonal antibodies. At the genetic level, two additional types, H or F/A, and X, have been described [3,4]. Each BoNT type is subdivided into subtypes based on amino acid sequence variations (0.9–36.2%, in most cases >2.6%). More than 40 subtypes have been identified [5]. Most BoNTs are produced by C. botulinum strains that are classified into physiologically and genetically distinct groups: group I including proteolytic strain types A, B, F; group II including non-proteolytic strain types B, E, F; group III corresponding to C. botulinum types C and D; and group IV, referred to as Clostridium argentinense type G. BoNTs are also synthesized by some other clostridial species, such as BoNT/E by atypical Clostridium butyricum strains and BoNT/F by atypical Clostridium baratii strains [5]. Moreover, a Paraclostridium bifermentans strain produces a BoNT-like neurotoxin (PMP1), which is insect-specific, whereas BoNTs are only active in vertebrates [6]. Sequences related to bont genes have been found in the genomes of a few other non-clostridial bacteria, such as bont/Wo or bont/I from Weisenella oryzae, a bacterium isolated from fermented rice, bont/J (ebont/F or bont/En) in an Enterococcus faecalis strain isolated from a cow, and Cp1 in Chryseobacterium piperi isolated from sediment [7,8,9,10,11,12]. However, the production of BoNT by non-clostridial strains has not been reported and their activity remains to be characterized.
In contrast to the heterogeneity of BoNTs- and BoNT-producing clostridia, TeNT is highly conserved [13,14]. Only a few amino acid variations were observed, notably a four-amino-acid insertion in four strains, in a study that investigated 37 C. tetani strains [13]. TeNT-producing C. tetani strains show a high level of genomic conservation. The population of C. tetani is divided into two major closely related clades and most strains belong to clade 1 [13].
3. Genetic Organization of Clostridial Neurotoxin Genes
BoNT-producing clostridia synthesize non-toxic proteins (associated-non-toxic proteins, ANTPs), which assemble with BoNTs through non-covalent bounds to form large-sized botulinum complexes (also referred to as progenitor toxins). A main characteristic of botulinum complexes is that they are stable at acidic pH and dissociate at alkaline pH (≥7) into free BoNT and ANTPs [15]. The genes encoding BoNTs and ANTPs are clustered in a DNA fragment called the botulinum locus. The genetic organization of the bont locus was initially determined in C. botulinum type C, where it is localized on a phage [16]. It consists of two operons transcribed in opposite directions. One operon contains bont and, located immediately upstream, ntnh, which encodes for the non-toxic non-hemagglutinin (NTNH) protein. NTNH shares a similar size and structure with BoNT, but NTNH lacks the catalytic site of BoNT. The interlocked association of NTNH with BoNT confers high resistance to acidic and protease degradation, while each protein separately is sensitive to proteolysis [17]. The ntnh-bont operon is highly conserved in all BoNT-producing clostridia, suggesting that ntnh and bont derive from a common ancestor by gene duplication [18,19,20]. The second operon is more divergent; it contains either hemagglutinin (HA) genes, including ha70, ha17, and ha33, or orfX genes (orfX1, orfX2, orfX3). The botulinum loci with orfX-bont genes encompass an additional gene in the first operon, p47, located upstream of ntnh [19,21]. A gene, botR, encoding an alternative sigma factor is localized in the botulinum loci upstream of the two operons in C. botulinum C and D and between the two operons in the other C. botulinum types, but it is lacking in C. botulinum E (Figure 1) [22,23]. HA complexes have been found to facilitate the passage of BoNT through the intestinal barrier by disrupting E-cadherin intercellular junctions between intestinal epithelial cells [24,25,26]. Up to now, no function has been attributed to OrfX and P47 proteins.
In C. tetani, the tent gene is localized on large plasmids. In comparison to the chromosome, these plasmids exhibit a higher degree of diversity. A homologous gene to botR, called tetR, lies immediately upstream of tent. No genes related to C. botulinum ANTP genes were identified in C. tetani genomes (Figure 1) [13,27,28].
4. Alternative Sigma Factors
BotR and TetR are the first factors that have been identified to be involved in the regulation of neurotoxin synthesis in C. botulinum and C. tetani. Overexpression of bot/R and tetR in C. botulinum and C. tetani, respectively, enhances neurotoxin synthesis (approximately 10-fold based on overexpression with a multicopy vector; toxin levels were quantified using toxin titration by mouse lethal activity), and inversely, their partial repression reduces toxin production (approximately 10-fold, based on experiments using an antisense mRNA strategy) [22,27]. Indeed, BotR and TetR are positive regulators of neurotoxin synthesis, and BotR also regulates the HA production in C. botulinum A [22]. Interestingly, overexpression of botR from C. botulinum type C in C. tetani stimulates TeNT synthesis, indicating a common mechanism of action of BotR and TetR [27]. BotR/A from C. botulinum A binds to conserved motifs in the two promoters of both operons, ntnh-bont/A and ha, in an RNA polymerase core enzyme-dependent manner. These motifs are also conserved in the promoter of tent and facilitate the binding of the TetR-RNA polymerase core enzyme complex. Thereby, BotR/A and TetR drive the transcription of the corresponding neurotoxin genes, and in addition, BotR/A controls that of the ha genes in C. botulinum [29]. Homologs of BotR and TetR have been characterized in Clostridioides difficile and Clostridium perfringens, where they positively control toxin synthesis. TcdR from C. difficile regulates the production of Toxin A (TcdA) and Toxin B (TcdB), and UviA controls the synthesis of a C. perfringens bacteriocin. BotR, TetR, TcdR, and UviA, which are interchangeable, as tested by in vitro transcription, are assigned to a distinct subgroup (group 5) of alternative sigma factors based on their targeted DNA motifs [30,31].
BotR and TetR are concomitantly expressed with their corresponding neurotoxin genes, showing a maximum level of expression at the end of the exponential growth phase and beginning of the stationary phase. The transcription levels of botR or tetR are approximately 100-fold less than those of ntnh-bont, has, or tent, respectively, as it is usually observed between regulatory and target genes [32,33]. Levels of neurotoxins secreted into the culture supernatant follow a similar pattern, including a progressive accumulation during the exponential growth, reaching a maximum level at the beginning of the stationary phase and maintaining a stable toxin level. Since botR and tetR are located in close proximity to neurotoxin and ANTP genes, they primarily have an impact on these target genes. However, botR and tetR might also regulate other distantly located genes, as is observed with most of the regulatory genes that have widely pleiotropic effects. Indeed, in C. botulinum A, botR seems to control the expression of at least 21 genes, 15 and 6 being up- and downregulated, respectively [34]. In contrast to primary sigma factors that are required for controlling house-keeping genes, alternative sigma factors are involved in controlling growth phase transitions, such as from the exponential growth to stationary phase, in response to environmental factors, including nutritional factors and stress conditions such as oxidative stress or heat shock. Their regulatory activity also comprises morphological differentiation, flagellar biosynthesis, and sporulation. In most cases, environmental bacteria that are exposed to a wide range of external factors employ more sigma factors than obligate pathogens or commensals, which are adapted to a specific host compartment and have a more restricted environment. Many obligate pathogens have lost sigma factors after host adaptation, such as Mycobacterium leprae, a strictly obligate pathogen that contains four sigma factors, compared to 13 sigma factors in Mycobacterium tuberculosis, which can live in the environment [35,36]. Indeed, C. botulinum A and C. tetani possess 18 and 25 sigma factors, respectively [37], compared to seven in Escherichia coli K12, six in Shigella flexneri, and three in Helicobacter pylori, the two latter species being strict human pathogens [38]. Group 4 alternative sigma factors, also called sigma factors of the extracytoplasmic function (ECF) family, are mainly involved in sensing and responding to extracellular signals and regulate cell envelope functions (transport, secretion, bacterial cell wall stress response) [31,36,39]. Group 5 alternative sigma factors, including BotR and TetR, are distantly related to the other sigma factors based on amino sequence similarity [30,31,36]. They control toxin synthesis and possibly other bacterial functions in response to yet unknown environmental stimuli or conditions.
In group III C. botulinum strains, the bont locus is located on a phage, and, in contrast to group I and II C. botulinum strains, where botR lies between the two has/orfX and ntnh-bont operons, botR/C or D is upstream of both has and ntnh-bont operons. The binding motifs recognized by BotR/A in the promoters of the has and ntnh-bont operons in C. botulinum A are conserved in the corresponding promoter regions of C. botulinum C and D [29]. BotR/C and D play likely a similar regulatory role of has and ntnh-bont operons as in C. botulinum A, but possibly do not respond to the same environmental stimuli, since group III C. botulinum strains have different physiological properties—for example, a higher optimal growth temperature; they are more often associated with animals than group I and II C. botulinum [40,41,42].
Despite the fact that C. botulinum E strains lack botR in close proximity to bont/E, the kinetics of growth, bont/E expression, and BoNT/E production are similar to those in C. botulinum A and proteolytic and non-proteolytic strains of C. botulinum B [33,43,44], supporting the notion that additional regulatory genes are involved in the control of neurotoxin genes. The non-proteolytic C. botulinum E, F6, non-proteolytic C. baratii F7, and non-proteolytic C. butyricum E contain an orfX-p47-ntnh-bont locus without botR. However, botR is present just upstream of the ha operon in non-proteolytic C. botulinum B4 and C. argentinense, which both have a ha-ntnh-bont botulinum locus, similar to C. botulinum C and D. In C. baratii F7, a gene encoding an UviA-like protein belonging to the same subgroup of sigma factors as BotR lies upstream of the orfX operon. This gene is also found in non-proteolytic C. botulinum E and F6, but distantly located of the botulinum locus [45]. The UviA-like protein in non-proteolytic strains might have a similar regulatory function of botulinum locus genes to BotR.
5. Two-Component Systems
Two-component systems (TCSs) consist of two proteins which coordinately control gene transcription in response to extracellular signals. TCSs regulate various bacterial physiological processes required for adaptation to environmental changes, such as development, cell division, metabolism, pathogenicity, and antibiotic/bacteriocin resistance. One component is a transmembrane protein with an extracellular sensor domain and, more rarely, a cytoplasmic domain, called sensor histidine kinase (SHK), which senses environmental stimuli including small molecules, ions, toxics, dissolved gases, pH, temperature, osmotic pressure, redox potential, or other yet unknown factors. SHK communicates with a corresponding response regulator (RR) by a phosphorelay. Most of the SHK and RR genes are closely located and organized in operons. Signal sensing by the N-terminal region of SHK induces the phosphorylation of a conserved histidine (His) in the C-terminal part. SHKs are in dimeric form and retain the phosphorylated His in a dimeric helical domain. The His phosphoryl group is then transferred to a conserved aspartate residue in the receiver domain of the corresponding RR. This results in a conformational change of the RR and enhanced affinity for specific promoter(s) [34,46,47,48].
C. botulinum and C. tetani contain numerous TCS genes in their genomes. In C. botulinum strain Hall, 39 genes have been found to encode RR proteins based on the presence of conserved motifs such as the signal receiver domain and DNA binding domain. Thirty RR genes are located in close proximity to a SHK gene, thus corresponding to 30 TCS operons. An additional nine RR genes and nine SHK genes are considered as orphan regulators [34,37]. C. tetani strain E88 possesses 30 TCS genes, 19 of which are homologous to related genes of C. botulinum A strain Hall, with ≥45% identity at the amino acid level (Figure 2) [32]. Most of the C. botulinum and C. tetani TCS genes have homologs in other clostridia, indicating that these clostridia might share similar regulatory pathways [32,34].
In C. botulinum A strain Hall, 34 regulatory genes (29 TCSs and five RR orphan regulatory genes) and an additional TCS gene in strain ATCC3502 (a derivative of strain Hall), as well as nine TCS genes in C. tetani E88, have been investigated for their possible regulatory contribution in toxinogenesis [32,34,49]. In C. botulinum A strain Hall, three TCSs have been reported as positive regulators of BoNT synthesis in a bontR-independent manner (as tested by an antisense mRNA strategy, yielding recombinant strains with 10- to 100-fold decreased BoNT/A production, which was determined by ELISA) [50]. Among them, one TCS belonging to the OmpR family shows 65% protein identity with a C. tetani TCS and shares similarity with VirI/VirJ of Clostridium perfringens, which has been reported as a regulator of toxin synthesis (GenBank BAA78773, BAA78774). Although this TCS is a positive regulator of BoNT synthesis, the C. tetani homolog is not involved in TeNT synthesis. The two other TCSs that are positive regulators of BoNT synthesis have no homolog in C. tetani. Two additional TCSs in C. botulinum show an indirect effect on BoNT production as they have pleiotropic effects, notably by impairing cell wall synthesis or assembly. The two C. tetani TCSs that positively regulate the TeNT synthesis (as tested by an antisense mRNA strategy, yielding recombinant strains with two- to five-fold decreased TeNT production, which was determined by ELISA) have ineffective homologs in BoNT synthesis in C. botulinum A [32] (Table 1). Only one TCS that is conserved (100% protein identity) in C. botulinum A strain ATCC3502 and in C. tetani E88 is a negative regulator of neurotoxin synthesis in both microorganisms (as evaluated by the ClosTron (http://www.clostron.com (accessed on 13 April 2022), Nottingham, UK) strategy in strain ATCC3502, where BoNT/A levels were monitored by ELISA; in strain E88, the antisense mRNA strategy was applied and TeNT levels were determined by ELISA) [32,49]. In C. botulinum, this TCS binds to both promoters of the ntnh-bont and ha operons and prevents their transcription by impairing the binding of BotR [49]. In C. tetani, the corresponding TCS binds also to the tent promoter and likely retains the same mechanism of action as in C. botulinum ATCC3502. However, the apparent TCS counterpart in C. botulinum strain Hall, which shows only 58% protein identity, is apparently not involved in the regulation of BoNT synthesis, as judged from the lack of BoNT level alterations assayed by ELISA in the recombinant Hall strain using the anti-sense RNA strategy targeting this TCS (Table 1) [32,50]. Thereby, various TCSs control the neurotoxin synthesis in C. botulinum and C. tetani. Although C. botulinum and C. tetani share homologous TCSs, most of them have distinct functional roles in the control of neurotoxin synthesis in the two microorganisms.
6. Metabolism and Toxin Gene Regulation
Toxin synthesis, as with protein synthesis in general, is dependent on the metabolic activity of the bacteria. How is neurotoxin synthesis linked to the general metabolism?
6.1. CodY
In Gram-positive bacteria, CodY (control of dciA (decoyinine induced operon) Y) is a master regulator of metabolism, sporulation, and virulence. In Bacillus subtilis, CodY controls more than 100 genes involved in the adaptation to nutrient restriction and transition from the exponential growth phase to the stationary growth phase. Typically, CodY acts by binding to the promoter of target genes in a GTP(guanosine triphosphate)- and branched-chain amino acid-dependent manner; these are indicators of the general metabolism status of the bacterium [51]. Thereby, in B. subtilis, CodY senses intracellular levels of GTP and branched amino acids such as isoleucine, whose levels are high during the exponential growth and decrease in mostly repressed gene transcription. In contrast, at low GTP or isoleucine levels, CodY induces the de-repression of genes which are involved in adaptive responses to nutrient limitation, such as those coding for extracellular degradative enzymes, transport systems, and catabolic pathways [51,52]. CodY is conserved in clostridia, including the toxigenic species C. botulinum, C. tetani, C. perfringens, and C. difficile. In C. botulinum A strain ATCC3502, CodY binds to the promoter of the ntnh-bont operon at high GTP levels, whereas isoleucine is ineffective, and stimulates toxin gene transcription and BoNT/A synthesis (as tested with the ClosTron system, and determining BoNT/A levels by ELISA and bont/A transcription by qPCR) [53]. The precise role of CodY in C. botulinum is still elusive: does CodY directly regulate ntnh-bont transcription or interfere with botR or a TCS gene such as by repressing the negative TCS regulator? CodY is also a positive regulator of TeNT synthesis in C. tetani (as tested by the antisense mRNA strategy, and determining TeNT levelsby ELISA, and tent transcription by qPCR) [32]. CodY binds to the tent promoter but not to that of tetR [32]. BoNT and TeNT synthesis is dependent on the availability of a carbon source such as glucose [54,55,56]. CodY controls carbon metabolism in B. subtilis. Notably, under glucose-rich conditions in culture medium, CodY and CcpA (catabolite control protein A), a regulator of carbon catabolism, facilitate the conversion of excess pyruvate resulting from glycolysis into excretable overflow compounds such as acetate, lactate, and acetoin [57]. A similar mechanism of CodY in glucose/pyruvate metabolism has been suggested in C. botulinum A [53]. CcpA is conserved in C. botulinum and C. tetani. However, the role of CcpA in these microorganisms remains to be elucidated. In contrast, in C. difficile, CodY and CcpA are negative regulators of toxin A (TcdA) and toxin B (TcdB). Glucose and rapidly metabolizable carbohydrates inhibit toxin synthesis in C. difficile. CodY and CcpA, which are activated by glucose and rapidly metabolizable carbohydrates, bind to the promoter of TcdR and repress its transcription and subsequently that of tcdA and tcdB [58,59,60]. The opposite regulatory pathways of toxin synthesis linked to carbohydrate metabolism controlled by CodY and CcpA between C. botulinum/C. tetani and C. difficile are intriguing. This indicates that toxin synthesis in C. botulinum and C. tertani requires energy from carbohydrate metabolism, mainly glucose, the main carbohydrate fermented by these bacteria, while C. difficile mainly uses amino acid metabolism as an energy source, notably through the Stickland reaction, for toxin production [60,61,62,63]. These divergent regulatory pathways might have evolved during bacterial adaptation to different environments: soil for C. botulinum/C. tetani and the intestine for C. difficile.
6.2. Spo0A
Spo0A is a master regulator of the initial steps of sporulation in Bacillus and clostridia. However, the mode of activation of Spo0A differs in the two classes of bacteria. Nutrient limitation, notably carbohydrate, nitrogen, and phosphorus limitation, is the major signal leading to Spo0A activation through a phosphorelay including five sensor kinases and subsequent positive transcriptional regulation of critical sporulation-essential genes [64]. The kinases that activate Spo0A in Bacillus are not conserved in clostridia. Orphan Spo0A-activating histidine kinases have been identified in clostridia, such as Clostridium acetobutylicum, C. perfringens, C. difficile, C. botulinum, and C. tetani. Clostridia sense different environmental stimuli to initiate sporulation, including external pH resulting from fermentation, with the subsequent release of acidic end-products (acetate, butyrate) and unknown factors [65,66,67,68,69]. In clostridia, Spo0A displays additional functions apart from sporulation initiation. In C. acetobutylicum, Spo0A is activated at the end of the exponential growth phase and controls the shift between acidogenesis that occurs during the exponential growth, and solventogenesis that is coupled to the onset of sporulation [70,71]. In C. perfringens, Spo0A controls the production of toxins (C. perfringens enterotoxin and TpeL), which are synthesized during the sporulation process [72,73]. The role of Spo0A in the regulation of TcdA and TcdB synthesis is variable according to the genetic background of C. difficile strains [62,74]. Spo0A coordinates the expression of a large number of C. difficile genes involved in multiple additional functions, such as nutrient transport, metabolic pathways including the production of butyrate, surface protein assembly, and flagellar biosynthesis [75].
Spo0A is highly conserved in all C. botulinum genomes: an orphan sensor histidine kinase that is able to phosphorylate Spo0A has been identified [69]. In C. botulinum ATCC3502, Spo0A is expressed during the exponential growth and its expression decreases during the entry into the stationary phase, while the subsequent transcription of sigma factors essential for sporulation increases [76,77]. It is not known whether Spo0A affects the expression of bont in group I C. botulinum. Adaptation to cultivation at high temperatures (45 °C) represses both bont/A and sporulation genes in strain ATCC3502, but no co-regulation of these genes has been evidenced [78]. No correlation between sporulation and the production of BoNT/A has been observed in two other C. botulinum A strains [33]. Moreover, the strain Hall A-hyper produces high levels of BoNT/A and is unable to sporulate [79]. Similarly, the highly TeNT-producing C. tetani strain used for vaccine production is a non-sporulating strain [80], and Spo0A has not been found to control TeNT synthesis (as tested by the antisense mRNA strategy, TeNT monitoring by ELISA, and tent transcription by qPCR) [32]. In contrast, in group II C. botulinum E, Spo0A is a positive regulator of BoNT/E synthesis and sporulation (as tested with the ClosTron system, toxin monitoring by ELISA, and gene transcription by qPCR). Spo0A binds to a conserved motif in the promoters of the ntnh-bont/A operon together with CodY, AbrB (putative repressor of bont/E), sigma K (belonging to the sigma factor cascade of sporulation), and an UviA-like regulator [43]. Thus, Spo0A might directly and indirectly regulate the transcription of bont/E. Thereby, group II C. botulinum E strains that have an UviA-like regulator instead of BotR likely use specific and common regulatory pathways of bont expression compared to C. botulinum A strains which belong to the distinct physiological and genetic group 1.
In group III C. botulinum C and D, the production of the C2 toxin, which is an ADP-ribosyltransferase targeting monomeric actin, is linked to sporulation [81]. However, the regulatory pathway of C2 toxin genes, and the possible involvement of spo0A and/or other sporulation genes, has not yet been elucidated.
6.3. Amino Acid/Peptide Metabolism
C. botulinum and C. tetani produce high levels of toxins in complex media rich in peptones and other nutrients, whereas chemically defined media even containing almost all amino acids and vitamins as well as a carbon source usually yield 10- to 100-fold lower toxin titers [55,82,83,84]. Licona-Cassani et al. showed that, although C. tetani grew in a chemically defined medium, toxin production was obtained only when casein-derived peptides were added to the medium [82]. In addition to variations in toxin production according to different media, variations in BoNT or TeNT yields are often observed from batch to batch of the same culture medium, even using the same bacterial strain. The transcription of neurotoxin genes and toxin synthesis occur mainly within a short time interval between the late exponential growth and early stationary growth phase [33,85,86]. Thus, nutritional and environmental factors influence the regulation of toxin synthesis in C. botulinum and C. tetani, which takes place in a restricted phase of bacterial growth. Peptides and amino acids appear to be important regulatory factors at the transcriptional and posttranscriptional levels. Indeed, large amounts (0.8–0.9 g/L) of amino acids (aspartate, glutamate, serine, histidine, threonine) downregulate tetR and tent by a yet non-identified regulatory pathway [86]. Arginine is an essential amino acid for C. botulinum growth, but an excess of arginine represses BoNT production in proteolytic group I C. botulinum [87]. Arginine deiminase leads to arginine catabolites that increase the pH and induce subsequent BoNT degradation by not yet characterized proteases. BoNT and botulinum complexes are stable at acidic pH in media without excess of arginine [88]. Supplementation with a high amount of glucose (50 g/L) that induces acidification counteracts the effect of arginine. Interestingly, BoNT synthesis is coupled to protease production [89]. Likely, proteases that are active at alkaline pH induce BoNT degradation. Secreted proteases are required for protein substrate degradation, resulting in peptides and amino acids that are taken up into the bacteria through transport systems and used for protein synthesis, including neurotoxin synthesis. Indeed, the C. botulinum A and C. tetani genomes contain numerous protease/peptidase and transport system genes [37,80].
Peptides in culture media were found to be critical for TeNT synthesis by C. tetani. Since culture media containing casein pancreatic digests support high levels of TeNT production, peptides derived from casein tryptic digestion were investigated. Histidine-containing peptides as well as hydrophobic peptides containing the motif proline–aromatic acid–proline were the most effective in promoting TeNT production [84,90,91]. It is noteworthy that genome analysis of C. tetani shows the presence of numerous peptidases and amino acid degradation pathways [92]. The kinetics of C. tetani growth in a complex medium show rapid exponential growth (stage I, around 10–12 h), then a slower linear growth (stage II, around 30 h), followed by a stationary phase and subsequent autolysis. During stage I, the amino acids are consumed and the genes involved in amino acid degradation pathways are overexpressed, corroborating amino acid catabolism that provides energy used for the rapid biomass formation during this growth phase. The pH decreases due to organic acid production, tetR and tent are not expressed, and TeNT is not synthesized. Once free amino acids are depleted in the culture medium, C. tetani uses peptides, whose metabolism requires transporters that are more energy-costly, and enters the linear growth phase II. The transition from free amino acid to peptide consumption is associated with increased pH due to the reduction of organic acids to alcohols and solvents, and the production of ammonia from peptide metabolism. During phase II, tetR and tent are highly expressed, as well as codY, the TCS that positively regulates tent, and two additional sigma factors located on the large plasmid containing tent, resulting in TeNT synthesis [54,82,86,93]. In complex media, glucose is consumed during the first phase of growth, leading to rapid bacterial multiplication and pH decrease. Then, the nitrogen source is used and the pH increases. TeNT is synthesized only during this second phase, when glucose is no longer or weakly available and when peptides are used for energy production. Thus, as shown by Fratelli et al., the balance between the nitrogen and carbon sources, as well as the subsequent pH of culture media, are critical factors [54,93]. C. botulinum and C. tetani likely share common metabolic pathways and subsequent toxin gene regulatory networks. In both microorganisms, the transition from amino acid utilization to peptides that are more common substrates in the environment seems to elicit the production of proteases. BoNT and TeNT are metalloproteases, but which recognize specific substrates in host neuronal cells. BoNT and TeNT possibly evolve from ancestor metalloproteases with a broader substrate range that were used by the bacteria for nutrient acquisition and that were regulated as the other proteases. Thus, the regulation of toxin synthesis in C. botulinum and C. tetani might represent a reminiscent common regulatory circuit controlling protease synthesis.
6.4. Other Nutritional and Environmental Factors
In addition to nutrients required for growth and protein synthesis, some nutritional and environmental factors might influence, directly or indirectly, toxin synthesis.
CO2—A high concentration of CO2 in the gas phase increases bont expression and BoNT synthesis in non-proteolytic group II C. botulinum B and E, although the growth rate is decreased. Indeed, a 70% CO2 atmosphere versus 10% stimulates 2- to 5-fold greater toxin gene expression and BoNT formation. In high and low CO2 concentrations, toxin gene expression occurs in the same growth phase, mainly in the late exponential growth and early stationary phase [94,95]. The signaling pathways in the regulation by CO2 are not known. CO2 can dissolve in the liquid medium and generate bicarbonate, which influences protein synthesis through carboxylation reactions. CO2 (35%) in the gas phase of C. tetani culture versus nitrogen atmosphere (unpublished) or the addition of sodium carbonate (100 mM) in the culture medium increases the production of TeNT approximately two-fold [32], despite reduced growth in the CO2-rich atmosphere (approximately three-fold). In contrast, elevated CO2 in the gas phase of proteolytic group I C. botulinum B and E has no effect on toxin gene expression [96], suggesting that CO2 triggers a signaling pathway controlling toxin synthesis in non-proteolytic strains.
Inorganic phosphate—Inorganic phosphate has been found to control TeNT synthesis in C. tetani. Supplementation of culture medium with inorganic phosphate (optimum concentration 40 mM) stimulates tent expression and TeNT production approximately three-fold without impairing the growth rate [32]. Inorganic phosphate is involved in multiple biochemical reactions; its effect on toxin gene transcription might be mediated by TCSs. C. tetani genome contains two TCSs putatively involved in phosphate uptake, one of which has been found to negatively regulate TeNT synthesis [32]. Inorganic phosphate is apparently not involved in BoNT production in C. botulinum A, as tested by supplementation of the TGY (trypticase-glucose-yeast extract) culture medium with 20 to 150 mM Na2HPO4 and monitoring BoNT/A production (strain Hall) in the culture supernatant by titration of the mouse lethal activity (unpublished). Control of the virulence mechanism by inorganic phosphate and TCS from the PhoP/PhoR family has been found in several pathogens [97,98]. TCSs control the homeostasis of phosphate according to the availability in the environment. However, the precise subsequent phosphate-dependent signaling pathways controlling virulence remain largely unknown.
pH—C. botulinum and C. tetani grow and produce the neurotoxins in a wide range of pH (pH 4.5–5 to 9). The initial pH of the growth medium was found to influence the autolysis of C. tetani. An initial pH of 6.1 seems optimal for TeNT production [84]. In a complex medium, high pH (7.8) downregulates tent [99]. The mechanism of toxin gene regulation by pH is not yet identified.
The culture pH of proteolytic C. botulinum A, B grown in complex media typically drops (pH 6–6.3 with initial glucose concentration up to 1%, and until pH 5.5 with glucose 1.5%) during the exponential growth phase, and then stabilizes and slightly increases during the stationary phase [56,85,88,100]. Maintaining an acidic pH (pH 5.7–6) during the culture does not modify the BoNT yield in the culture supernatant, whereas an alkaline pH (pH 7.2 and above, manually adjusted or by supplementation of the culture medium with 2% arginine) decreases the BoNT level [56,88]. The pH does not influence BoNT synthesis at the transcriptional level, but affects BoNT stability by activating a BoNT degrading metalloprotease in alkaline conditions [88].
Temperature—In contrast to C. difficile, in which a high temperature (42 °C) prevents tcdR and toxin gene expression, temperatures of 37–44 °C have no influence on botR and bont transcription in group I C. botulinum A. However, a high temperature induces the production of protease(s), which inactivate BoNT/A [33]. TeNT production is usually obtained by C. tetani culture at 33–35 °C [82,84,99].
Group II C. botulinum has an optimum temperature of 25 °C for growth and toxin production, but the strains of this group can grow and form toxins at temperatures as low as 3.0–3.3 °C in 5 to 7 weeks [101]. Investigation with C. botulinum E showed that growth and toxin production are lower at 10 °C than at 30 °C. However, bontE transcription relative to growth was similar at 10 °C and 30 °C [102]. A TCS is involved in the cold adaptation of C. botulinum E [103,104]. Similarly, cold tolerance of growth at 15 °C in C. botulinum A strain ATCC3502 requires the contribution of a TCS [105]. This TCS is conserved in C. botulinum A strain Hall, but it has not been identified as a regulator of BoNT/A synthesis [50]. Thus, temperature is important for growth and toxin production, but temperature seems to have no direct role in the regulation of toxin synthesis in C. botulinum and C. tetani.
7. Small RNA
In addition to regulatory proteins, bacteria use regulatory RNAs to modulate gene transcription or translation initiation for adaptive responses to environmental changes. Environmental bacteria have to adapt their physiology and metabolism to various hostile conditions and pathogenic bacteria have to cope with adverse host interactions; thus, they have to adapt rapidly their gene expression, notably that of virulence genes [106,107]. Regulatory RNAs are small molecules, typically between 50 and 500 nucleotides (small RNAs or sRNAs), and are non-protein coding. The main advantage of sRNAs versus regulatory proteins is their speed in controlling gene expression based on the faster availability of sRNAs due to the lower energy needed for their production by transcription and not by translation, as in the case of regulatory proteins, the faster turnover of sRNAs since RNAs are less stable than proteins, the rapid control of mRNA function by pairing with specific motifs in the untranslated region (UTR), or, in some cases, by acting at a posttranscriptional level [108,109]. Although most sRNAs are inhibitors of gene expression, some are activators. sRNAs lie in or overlap with the 5′ or 3′ UTRs of target genes, in intergenic regions, or in the opposite DNA strand and are transcribed as antisense sRNAs. Based on their genomic localization and mechanism of action, sRNAs are divided into several classes: cis-encoded sRNAs are transcribed from the DNA strand opposite to the target sequence and interact by perfect base pairing with mRNAs; trans-encoded sRNAs are distantly transcribed from target mRNA genes and recognize their target mRNAs by multiple and discontinuous short contacts. Clustered, regularly interspaced short palindromic repeat (CRISPR) RNAs interact with foreign DNA or RNA. Another class of sRNAs bind to regulatory proteins and antagonize their function. RNA riboswitches sense metabolites or environmental cues; they are usually located in the 5′ UTR of their target genes. Certain sRNAs interact with regulatory proteins, notably by promoting protein sequestration, but most sRNAs interfere with mRNA by inhibition of their translation, such as by blocking the ribosome binding site and/or by impairment of mRNA stability, resulting in subsequent degradation [108,109,110,111]. Numerous sRNAs act at a posttranscriptional level, whereas regulatory proteins preferentially act at transcriptional steps. In fact, numerous bacterial gene regulations involve mixed regulatory networks, including both transcriptionally acting regulatory proteins and posttranscriptionally acting sRNAs [112].
sRNAs are widespread in Gram-positive bacteria and notably in clostridia. In environmental clostridia such as C. acetobutylicum, sRNAs (159 predicted) have a crucial role in solventogenesis, growth, and the response to toxic metabolites [113,114,115]. In pathogenic clostridia, sRNAs have been described in C. perfringens to be involved in toxin production, as well as in Clostridioides difficile in adaptation to host and anti-phage defenses [116,117,118]. More than 200 sRNAs are predicted in the genomes of groups I and II C. botulinum, and 137 in the genome of C. tetani E88 [119]. A sRNA has been identified in the 3′ UTR of tent in the C. tetani E88 strain that is conserved in all toxigenic C. tetani strains [120]. However, no such sRNA has been reported in C. botulinum. This sRNA is expressed concomitantly with tent and negatively regulates tent expression and TeNT synthesis. The sRNA (approximately 140 nucleotides) contains a predicted junction-loop-exposed 14-nucleotide-long sequence that perfectly matches to a complementary sequence in the 5′ region of tent mRNA. Thus, the sRNA-mediated inhibitory regulatory activity is likely based on the sequestration of tent mRNA. In addition, this sRNA impairs C. tetani growth, notably by reducing the exponential growth phase [120]. Pleiotropic effects of sRNAs have also been found for the regulatory RNA VR-RNA of C. perfringens that controls 147 genes, including genes of toxins and virulence factors (alpha-toxin, kappa-toxin, hyaluronidase, sialidases) as well as genes involved in capsule synthesis [116,121].
8. Quorum Sensing
Quorum sensing is a cell-to-cell communication that bacteria use to adapt their physiology and behavior in response to cell densities. Bacteria produce and secrete extracellular signaling molecules called autoinducers. When a certain threshold of bacterial density is reached, accumulated autoinducers are detected by the bacteria, leading to coordinated changes in gene expression and behavior. Thus, quorum sensing allows a synchronized adaptation to environmental conditions in nonclonal bacterial populations [122,123]. A well-characterized quorum-sensing system is the Agr system of Staphylococcus aureus [124]. The autoinducer is a small peptide (autoinducing peptide, AIP) produced by agrD as a precursor, which is processed by the membrane endoprotease AgrB. When AIP reaches a threshold concentration in the environment, it is sensed by the TCS AgrA/AgrC, which in turn upregulates the expression of a small RNA and subsequently stimulates toxin synthesis. AgrC is the histidine kinase cell surface receptor that recognizes AIP and activates the response regulator AgrA [122]. Quorum sensing has been evidenced in C. botulinum [125], and an Agr-like system has been identified in group I C. botulinum strains called agr-1/agr-2; agr-1 and agr-2 are homologs of agrB, and agrD, respectively [126]. However, the Agr system in C. botulinum plays a different role than in S. aureus. Agr-1 seems to be involved in sporulation, while Agr-2 possibly controls toxin production [126]. Homologous genes of agrA and agrC have also been identified in C. botulinum A Hall strain, but silencing agrA did not impact BoNT/A production [34]. Thus, the regulatory quorum-sensing pathways in C. botulinum remain to be defined. A computational model of group I C. botulinum A growth and toxin production based on nutrient availability, cell density, and quorum-sensing signaling has been proposed in agreement with experimental data [127].
It is noteworthy that a cultivation technique was developed in the 1950s and 1960s to obtain high levels of toxin [128]. This method consists of dialyzed cultures. The bacteria are inoculated inside a dialysis bag (usually a cellophane bag) containing saline, which is immersed in a culture medium. This technique was used for the production of BoNT and TeNT, with a 5–50-fold increase in toxin yields compared to cultures in a flask with the same culture medium [128,129,130,131]. A very high bacterial density is obtained by culturing clostridia in dialysis bags and likely a quorum-sensing-mediated regulation is involved in the production of high toxin levels. Moreover, the high bacterial density leads to increased autolysis, which contributes to toxin release into the extracellular medium.
9. Concluding Remarks
The toxigenic environmental bacteria C. botulinum and C. tetani contain multiple and complex regulatory networks to control neurotoxin production. These only partially deciphered networks include alternative sigma factors, TCSs, sRNAs, quorum-sensing systems, and regulators of bacterial metabolism that interlock the bacterial growth with toxin production (Figure 3). Both microbial species share some common regulatory mechanisms, notably an alternative sigma factor, the gene of which is located upstream of the neurotoxin gene, and an inhibitory TCS. They retain a similar kinetic pattern of toxin production, mainly occurring at the transition between the exponential growth and early stationary growth phases. This is possibly linked to the alternative sigma factors BotR and TetR, which are expressed concomitantly with the neurotoxin genes. However, C. botulinum and C. tetani use distinct signaling pathways, notably distinct TCSs, probably reflecting the recognition of different nutritional and/or environmental signals. Although toxinogenesis is dependent on the general metabolism in C. botulinum and C. tetani, the nutritional requirements for toxinogenesis seem different between these microorganisms. However, only a few nutritional and environmental factors controlling the toxinogenesis have so far been identified, such as CO2 in group II C. botulinum and inorganic phosphate in C. tetani. Differences in nutritional factors, particularly in the nature and composition of peptides and amino acids required for toxinogenesis, seem a major hallmark between C. botulinum and C. tetani. This likely reflects the different ecological niches used by these bacteria. Group I C. botulinum prefers neutral to slightly alkaline soils with low organic content, while group II C. botulinum are mostly found in more acidic soils with high levels of organic matter or as commensals in the intestines of certain animals. C. tetani is found primarily in neutral or alkaline soils at sufficient temperatures (>20 °C) and levels of moisture (15%) [41,132]. Moreover, group I but not group II C. botulinum strains can colonize the intestines of humans and produce BoNT in situ, leading to infant botulism or adult intestinal toxemia botulism [133]. C. tetani has not been reported to colonize the digestive tract and to induce intestinal tetanus [134]. Thus, C. botulinum groups and C. tetani have adapted to particular environments, notably through complex and specific regulatory systems that sense extracellular signals, leading to adapted gene expression. In addition to regulatory proteins and sRNAs, clostridia sense environmental factors by specific arrays of surface-associated proteins [135]. Do BoNTs and TeNT represent adaptive factors? These neurotoxins that attack specifically the nervous systems of vertebrates seem not to be involved in environmental adaptation. Indeed, non-toxigenic strains of C. botulinum and C. tetani can multiply, sporulate, and survive in the environment in the same manner as their toxigenic counterparts. BoNTs and TeNT, which likely evolved from a common protease ancestor, possibly retain common regulatory mechanisms with other proteases/peptidases required for the utilization of specific nutrient sources [20,136]. This is further supported by the observation that toxin synthesis is initiated at the transition from amino acid/carbohydrate to peptide metabolism in C. Tetani [54,93], and possibly in C. botulinum.
Author Contributions
Conceptualization, writing and editing, M.R.P.; review, H.B. All authors have read and agreed to the published version of the manuscript.
Funding
This review received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dong, M.; Masuyer, G.; Stenmark, P. Botulinum and Tetanus Neurotoxins. Annu. Rev. Biochem. 2019, 88, 811–837. [Google Scholar] [CrossRef] [PubMed]
- Pirazzini, M.; Montecucco, C.; Rossetto, O. Toxicology and pharmacology of botulinum and tetanus neurotoxins: An update. Arch. Toxicol. 2022, 96, 1521–1539. [Google Scholar] [CrossRef] [PubMed]
- Dover, N.; Barash, J.R.; Hill, K.K.; Xie, G.; Arnon, S.S. Molecular characterization of a novel botulinum neurotoxin type H gene. J. Infect. Dis. 2014, 209, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Masuyer, G.; Zhang, J.; Shen, Y.; Lundin, D.; Henriksson, L.; Miyashita, S.I.; Martinez-Carranza, M.; Dong, M.; Stenmark, P. Identification and characterization of a novel botulinum neurotoxin. Nat. Commun. 2017, 8, 14130. [Google Scholar] [CrossRef]
- Peck, M.W.; Smith, T.J.; Anniballi, F.; Austin, J.W.; Bano, L.; Bradshaw, M.; Cuervo, P.; Cheng, L.W.; Derman, Y.; Dorner, B.G.; et al. Historical Perspectives and Guidelines for Botulinum Neurotoxin Subtype Nomenclature. Toxins 2017, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Contreras, E.; Masuyer, G.; Qureshi, N.; Chawla, S.; Dhillon, H.S.; Lee, H.L.; Chen, J.; Stenmark, P.; Gill, S.S. A neurotoxin that specifically targets Anopheles mosquitoes. Nat. Commun. 2019, 10, 2869. [Google Scholar] [CrossRef]
- Zornetta, I.; Azarnia Tehran, D.; Arrigoni, G.; Anniballi, F.; Bano, L.; Leka, O.; Zanotti, G.; Binz, T.; Montecucco, C. The first non Clostridial botulinum-like toxin cleaves VAMP within the juxtamembrane domain. Sci. Rep. 2016, 6, 30257. [Google Scholar] [CrossRef]
- Brunt, J.; Carter, A.T.; Stringer, S.C.; Peck, M.W. Identification of a novel botulinum neurotoxin gene cluster in Enterococcus. FEBS Lett. 2018, 592, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Lebreton, F.; Mansfield, M.J.; Miyashita, S.I.; Zhang, J.; Schwartzman, J.A.; Tao, L.; Masuyer, G.; Martinez-Carranza, M.; Stenmark, P.; et al. Identification of a Botulinum Neurotoxin-like Toxin in a Commensal Strain of Enterococcus faecium. Cell Host Microbe 2018, 23, 169–176.e6. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, M.J.; Wentz, T.G.; Zhang, S.; Lee, E.J.; Dong, M.; Sharma, S.K.; Doxey, A.C. Bioinformatic discovery of a toxin family in Chryseobacterium piperi with sequence similarity to botulinum neurotoxins. Sci. Rep. 2019, 9, 1634. [Google Scholar] [CrossRef]
- Wentz, T.G.; Muruvanda, T.; Lomonaco, S.; Thirunavukkarasu, N.; Hoffmann, M.; Allard, M.W.; Hodge, D.R.; Pillai, S.P.; Hammack, T.S.; Brown, E.W.; et al. Closed Genome Sequence of Chryseobacterium piperi Strain CTM(T)/ATCC BAA-1782, a Gram-Negative Bacterium with Clostridial Neurotoxin-Like Coding Sequences. Genome Announc. 2017, 5, e01296-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, M.; Stenmark, P. The Structure and Classification of Botulinum Toxins. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–23. [Google Scholar]
- Chapeton-Montes, D.; Plourde, L.; Bouchier, C.; Ma, L.; Diancourt, L.; Criscuolo, A.; Popoff, M.R.; Bruggemann, H. The population structure of Clostridium tetani deduced from its pan-genome. Sci. Rep. 2019, 9, 11220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, J.E.; Wang, R.; Shen, R.F.; Wu, W.W.; Keller, J.E. Comparative pathogenomics of Clostridium tetani. PLoS ONE 2017, 12, e0182909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.R.; Wang, T.; Kukreja, R.; Cai, S. The botulinum neurotoxin complex and the role of ancillary proteins. In Molecular Aspects of Botulinum Neurotoxin; Foster, K.A., Ed.; Springer: New York, NY, USA, 2014; Volume 4, pp. 68–101. [Google Scholar]
- Hauser, D.; Eklund, M.W.; Boquet, P.; Popoff, M.R. Organization of the botulinum neurotoxin C1 gene and its associated non-toxic protein genes in Clostridium botulinum C468. Mol. Gen. Genet. 1994, 243, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Rumpel, S.; Zhou, J.; Strotmeier, J.; Bigalke, H.; Perry, K.; Shoemaker, C.B.; Rummel, A.; Jin, R. Botulinum neurotoxin is shielded by NTNHA in an interlocked complex. Science 2012, 335, 977–981. [Google Scholar] [CrossRef] [Green Version]
- Doxey, A.C.; Lynch, M.D.; Muller, K.M.; Meiering, E.M.; McConkey, B.J. Insights into the evolutionary origins of clostridial neurotoxins from analysis of the Clostridium botulinum strain A neurotoxin gene cluster. BMC Evol. Biol. 2008, 8, 316. [Google Scholar] [CrossRef] [Green Version]
- Popoff, M.R.; Marvaud, J.C. Structural and genomic features of clostridial neurotoxins. In The Comprehensive Sourcebook of Bacterial Protein Toxins, 2nd ed.; Alouf, J.E., Freer, J.H., Eds.; Academic Press: London, UK, 1999; Volume 2, pp. 174–201. [Google Scholar]
- Popoff, M.R.; Bouvet, P. Genetic characteristics of toxigenic Clostridia and toxin gene evolution. Toxicon 2013, 75, 63–89. [Google Scholar] [CrossRef]
- Hill, K.K.; Smith, T.J. Genetic diversity within Clostridium botulinum serotypes, botulinum neurotoxin gene clusters and toxin subtypes. Curr. Top. Microbiol. Immunol. 2013, 364, 1–20. [Google Scholar]
- Marvaud, J.C.; Gibert, M.; Inoue, K.; Fujinaga, V.; Oguma, K.; Popoff, M.R. botR is a positive regulator of botulinum neurotoxin and associated non toxic protein genes in Clostridium botulinum A. Mol. Microbiol. 1998, 29, 1009–1018. [Google Scholar] [CrossRef]
- Poulain, B.; Molgo, J.; Popoff, M.R. Clostridial neurotoxins: From the cellular and molecular mode of action to their therapeutic use. In The Comprehensive Sourcebook of Bacterial Protein Toxins, 4th ed.; Alouf, J., Ladant, D., Popoff, M.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 287–336. [Google Scholar]
- Fujinaga, Y.; Popoff, M.R. Translocation and dissemination of botulinum neurotoxin from the intestinal tract. Toxicon 2018, 147, 13–18. [Google Scholar] [CrossRef]
- Lee, K.; Zhong, X.; Gu, S.; Kruel, A.M.; Dorner, M.B.; Perry, K.; Rummel, A.; Dong, M.; Jin, R. Molecular basis for disruption of E-cadherin adhesion by botulinum neurotoxin A complex. Science 2014, 344, 1405–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, T.; Sugawara, Y.; Yutani, M.; Amatsu, S.; Yagita, H.; Kohda, T.; Fukuoka, S.; Nakamura, Y.; Fukuda, S.; Hase, K.; et al. Botulinum toxin A complex exploits intestinal M cells to enter the host and exert neurotoxicity. Nat. Commun. 2015, 6, 6255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marvaud, J.C.; Eisel, U.; Binz, T.; Niemann, H.; Popoff, M.R. tetR is a positive regulator of the Tetanus toxin gene in Clostridium tetani and is homologous to botR. Infect. Immun. 1998, 66, 5698–5702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruggemann, H. Genomics of clostridial pathogens: Implication of extrachromosomal elements in pathogenicity. Curr. Opin. Microbiol. 2005, 8, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Raffestin, S.; Dupuy, B.; Marvaud, J.C.; Popoff, M.R. BotR/A and TetR are alternative RNA polymerase sigma factors controlling the expression of the neurotoxin and associated protein genes in Clostridium botulinum type A and Clostridium tetani. Mol. Microbiol. 2005, 55, 235–249. [Google Scholar] [CrossRef]
- Dupuy, B.; Raffestin, S.; Matamouros, S.; Mani, N.; Popoff, M.R.; Sonenshein, A.L. Regulation of toxin and bacteriocin gene expression in Clostridium by interchangeable RNA polymerase sigma factors. Mol. Microbiol. 2006, 60, 1044–1057. [Google Scholar] [CrossRef]
- Helmann, J.D. The extracytoplasmic function (ECF) sigma factors. Adv. Microbial. Physiol. 2002, 46, 47–110. [Google Scholar]
- Chapeton-Montes, D.; Plourde, L.; Deneve, C.; Garnier, D.; Barbirato, F.; Colombié, V.; Demay, S.; Haustant, G.; Gorgette, O.; Schmitt, C.; et al. Tetanus Toxin Synthesis is Under the Control of A Complex Network of Regulatory Genes in Clostridium tetani. Toxins 2020, 12, 328. [Google Scholar] [CrossRef]
- Couesnon, A.; Raffestin, S.; Popoff, M.R. Expression of botulinum neurotoxins A and E, and associated non-toxin genes, during the transition phase and stability at high temperature: Analysis by quantitative reverse transcription-PCR. Microbiology 2006, 152, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Connan, C.; Popoff, M.R. Two-component systems and toxinogenesis regulation in Clostridium botulinum. Res. Microbiol. 2015, 166, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, T.; Bose, D.; Zhang, X. Mechanisms for activating bacterial RNA polymerase. FEMS Microbiol. Rev. 2010, 34, 611–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, M.; Rowley, G.; Kormanec, J.; Zalm, M.E.J. The Role of Alternative Sigma Factors in Pathogen Virulence. In Foodborne Pathogens; Gurtler, J., Doyle, M., Kornacki, J., Eds.; Springer: Cham, Switzerland, 2017; pp. 229–303. [Google Scholar] [CrossRef]
- Sebaihia, M.; Peck, M.W.; Minton, N.P.; Thomson, N.R.; Holden, M.T.; Mitchell, W.J.; Carter, A.T.; Bentley, S.D.; Mason, D.R.; Crossman, L.; et al. Genome sequence of a proteolytic (Group I) Clostridium botulinum strain Hall A and comparative analysis of the clostridial genomes. Genome Res. 2007, 17, 1082–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, H.; Ussery, D.W. Sigma factors in a thousand E. coli genomes. Environ. Microbiol. 2013, 15, 3121–3129. [Google Scholar] [CrossRef] [PubMed]
- Gruber, T.M.; Gross, C.A. Multiple Sigma Subunits and the Partitioning of Bacterial Transcription Space. Annu. Rev. Microbiol. 2003, 57, 441–466. [Google Scholar] [CrossRef] [PubMed]
- Espelund, M.; Klaveness, D. Botulism outbreaks in natural environments—An update. Front. Microbiol. 2014, 5, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popoff, M.R. Ecology of neurotoxigenic strains of Clostridia. In Clostridial Neurotoxins: The Molecular Pathogenesis of Tetanus and Botulism; Montecucco, C., Ed.; Springer: Berlin/Heidelberg, Germany, 1995; Volume 195, pp. 1–29. [Google Scholar]
- Woudstra, C.; Skarin, H.; Anniballi, F.; Fenicia, L.; Bano, L.; Drigo, I.; Koene, M.; Bayon-Auboyer, M.H.; Buffereau, J.P.; De Medici, D.; et al. Neurotoxin gene profiling of clostridium botulinum types C and D native to different countries within Europe. Appl. Environ. Microbiol. 2012, 78, 3120–3127. [Google Scholar] [CrossRef] [Green Version]
- Mascher, G.; Mertaoja, A.; Korkeala, H.; Lindstrom, M. Neurotoxin synthesis is positively regulated by the sporulation transcription factor Spo0A in Clostridium botulinum type E. Environ. Microbiol. 2017, 19, 4287–4300. [Google Scholar] [CrossRef] [PubMed]
- Lovenklev, M.; Holst, E.; Borch, E.; Radstrom, P. Relative neurotoxin gene expression in Clostridium botulinum type B, determined using quantitative reverse transcription-PCR. Appl. Environ. Microbiol. 2004, 70, 2919–2927. [Google Scholar] [CrossRef] [Green Version]
- Dover, N.; Barash, J.R.; Burke, J.N.; Hill, K.K.; Detter, J.C.; Arnon, S.S. Arrangement of the Clostridium baratii F7 toxin gene cluster with identification of a sigma factor that recognizes the botulinum toxin gene cluster promoters. PLoS ONE 2014, 9, e97983. [Google Scholar] [CrossRef] [Green Version]
- Casino, P.; Rubio, V.; Marina, A. The mechanism of signal transduction by two-component systems. Curr. Opin. Struct. Biol. 2010, 20, 763–771. [Google Scholar] [CrossRef]
- Jacob-Dubuisson, F.; Mechaly, A.; Betton, J.M.; Antoine, R. Structural insights into the signalling mechanisms of two-component systems. Nat. Rev. Microbiol. 2018, 16, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Zschiedrich, C.P.; Keidel, V.; Szurmant, H. Molecular Mechanisms of Two-Component Signal Transduction. J. Mol. Biol. 2016, 428, 3752–3775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Korkeala, H.; Dahlsten, E.; Sahala, E.; Heap, J.T.; Minton, N.P.; Lindstrom, M. Two-component signal transduction system CBO0787/CBO0786 represses transcription from botulinum neurotoxin promoters in Clostridium botulinum ATCC 3502. PLoS Pathog. 2013, 9, e1003252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connan, C.; Brueggemann, H.; Mazuet, C.; Raffestin, S.; Cayet, N.; Popoff, M.R. Two-Component Systems Are Involved in the Regulation of Botulinum Neurotoxin Synthesis in Clostridium botulinum Type A Strain Hall. PLoS ONE 2012, 7, e41848. [Google Scholar]
- Sonenshein, A.L. CodY, a global regulator of stationary phase and virulence in Gram-positive bacteria. Curr. Opin. Microbiol. 2005, 8, 203–207. [Google Scholar] [CrossRef]
- Ratnayake-Lecamwasam, M.; Serror, P.; Wong, K.W.; Sonenshein, A.L. Bacillus subtilis CodY represses early-stationary-phase genes by sensing GTP levels. Genes Dev. 2001, 15, 1093–1103. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Dahlsten, E.; Korkeala, H.; Lindström, M. Positive Regulation of Botulinum Neurotoxin Gene Expression by CodY in Clostridium botulinum ATCC 3502. Appl. Environ. Microbiol. 2014, 80, 7651–7658. [Google Scholar] [CrossRef] [Green Version]
- Fratelli, F.; Siquini, T.J.; de Abreu, M.E.; Higashi, H.G.; Converti, A.; de Carvalho, J.C. Fed-batch production of tetanus toxin by Clostridium tetani. Biotechnol. Prog. 2010, 26, 88–92. [Google Scholar] [CrossRef]
- Shone, C.C.; Tranter, H.S. Growth of Clostridia and preparation of their neurotoxins. In Clostridial Neurotoxins; Montecucco, C., Ed.; Springer: Berlin/Heidelberg, Germany, 1995; Volume 195, pp. 143–160. [Google Scholar]
- Siegel, L.S.; Metzger, J.F. Toxin production by Clostridium botulinum type A under various fermentation conditions. Appl. Environ. Microbiol. 1979, 38, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Sonenshein, A.L. Control of key metabolic intersections in Bacillus subtilis. Nat. Rev. Microbiol. 2007, 5, 917–927. [Google Scholar] [CrossRef]
- Antunes, A.; Martin-Verstraete, I.; Dupuy, B. CcpA-mediated repression of Clostridium difficile toxin gene expression. Mol. Microbiol. 2011, 79, 882–899. [Google Scholar] [CrossRef] [PubMed]
- Dineen, S.S.; Villapakkam, A.C.; Nordman, J.T.; Sonenshein, A.L. Repression of Clostridium difficile toxin gene expression by CodY. Mol. Microbiol. 2007, 66, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.R.; Somerville, G.A.; Sonenshein, A.L. Regulating the Intersection of Metabolism and Pathogenesis in Gram-positive Bacteria. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Antunes, A.; Camiade, E.; Monot, M.; Courtois, E.; Barbut, F.; Sernova, N.V.; Rodionov, D.A.; Martin-Verstraete, I.; Dupuy, B. Global transcriptional control by glucose and carbon regulator CcpA in Clostridium difficile. Nucleic Acids Res. 2012, 40, 10701–10718. [Google Scholar] [CrossRef] [Green Version]
- Bouillaut, L.; Dubois, T.; Sonenshein, A.L.; Dupuy, B. Integration of metabolism and virulence in Clostridium difficile. Res. Microbiol. 2015, 166, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Girinathan, B.P.; DiBenedetto, N.; Worley, J.N.; Peltier, J.; Arrieta-Ortiz, M.L.; Immanuel, S.R.C.; Lavin, R.; Delaney, M.L.; Cummins, C.K.; Hoffman, M.; et al. In vivo commensal control of Clostridioides difficile virulence. Cell Host Microbe 2021, 29, 1693–1708.e1697. [Google Scholar] [CrossRef]
- Sonenshein, A.L. Control of sporulation initiation in Bacillus subtilis. Curr. Opin. Microbiol. 2000, 3, 561–566. [Google Scholar] [CrossRef]
- Dürre, P. Ancestral sporulation initiation. Mol Microbiol. 2011, 80, 584–587. [Google Scholar] [CrossRef]
- Dürre, P. Physiology and Sporulation in Clostridium. Microbiol. Spectr. 2014, 2, TBS-0010-2012. [Google Scholar] [CrossRef] [Green Version]
- Paredes, C.J.; Alsaker, K.V.; Papoutsakis, E.T. A comparative genomic view of clostridial sporulation and physiology. Nat. Rev. Microbiol. 2005, 3, 969–978. [Google Scholar] [CrossRef]
- Talukdar, P.K.; Olguín-Araneda, V.; Alnoman, M.; Paredes-Sabja, D.; Sarker, M.R. Updates on the sporulation process in Clostridium species. Res. Microbiol. 2015, 166, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Wörner, K.; Szurmant, H.; Chiang, C.; Hoch, J.A. Phosphorylation and functional analysis of the sporulation initiation factor Spo0A from Clostridium botulinum. Mol. Microbiol. 2006, 59, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Ravagnani, A.; Jennert, K.C.; Steiner, E.; Grunberg, R.; Jefferies, J.R.; Wilkinson, S.R.; Young, D.I.; Tidswell, E.C.; Brown, D.P.; Youngman, P.; et al. Spo0A directly controls the switch from acid to solvent production in solvent-forming clostridia. Mol. Microbiol. 2000, 37, 1172–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dürre, P. Metabolic networks in Clostridium acetobutylicum: Interaction of sporulation, solventogenesis and toxin formation. In Clostridia, Molecular Biology in the Postgenomic Era; Brüggemann, H., Gottschalk, G., Eds.; Caister Academic Press: Norfolk, UK, 2009; pp. 215–227. [Google Scholar]
- Harry, K.H.; Zhou, R.; Kroos, L.; Melville, S.B. Sporulation and enterotoxin (CPE) synthesis are controlled by the sporulation-specific sigma factors SigE and SigK in Clostridium perfringens. J. Bacteriol. 2009, 191, 2728–2742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes-Sabja, D.; Sarker, N.; Sarker, M.R. Clostridium perfringens tpeL is expressed during sporulation. Microb. Pathog. 2011, 51, 384–388. [Google Scholar] [CrossRef]
- Mackin, K.E.; Carter, G.P.; Howarth, P.; Rood, J.I.; Lyras, D. Spo0A differentially regulates toxin production in evolutionarily diverse strains of Clostridium difficile. PLoS ONE 2013, 8, e79666. [Google Scholar] [CrossRef]
- Pettit, L.J.; Browne, H.P.; Yu, L.; Smits, W.K.; Fagan, R.P.; Barquist, L.; Martin, M.J.; Goulding, D.; Duncan, S.H.; Flint, H.J.; et al. Functional genomics reveals that Clostridium difficile Spo0A coordinates sporulation, virulence and metabolism. BMC Genom. 2014, 15, 160. [Google Scholar] [CrossRef] [Green Version]
- Kirk, D.G.; Palonen, E.; Korkeala, H.; Lindstrom, M. Evaluation of normalization reference genes for RT-qPCR analysis of spo0A and four sporulation sigma factor genes in Clostridium botulinum Group I strain ATCC 3502. Anaerobe 2014, 26, 14–19. [Google Scholar] [CrossRef]
- Kirk, D.G.; Zhang, Z.; Korkeala, H.; Lindstrom, M. Alternative sigma factors SigF, SigE, and SigG are essential for sporulation in Clostridium botulinum ATCC 3502. Appl. Environ. Microbiol. 2014, 80, 5141–5150. [Google Scholar] [CrossRef] [Green Version]
- Selby, K.; Mascher, G.; Somervuo, P.; Lindström, M.; Korkeala, H. Heat shock and prolonged heat stress attenuate neurotoxin and sporulation gene expression in group I Clostridium botulinum strain ATCC 3502. PLoS ONE 2017, 12, e0176944. [Google Scholar] [CrossRef]
- Dineen, S.S.; Bradshaw, M.; Johnson, E.A. Neurotoxin gene clusters in Clostridium botulinum type A strains: Sequence comparison and evolutionary implications. Curr. Microbiol. 2003, 46, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Brüggemann, H.; Bäumer, S.; Fricke, W.F.; Wiezr, A.; Liesagang, H.; Decker, I.; Herzberg, C.; Martinez-Arias, R.; Henne, A.; Gottschalk, G. The genome sequence of Clostridium tetani, the causative agent of tetanus disease. Proc. Natl. Acad. Sci. USA 2003, 100, 1316–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, S.; Serikawa, T.; Yamakawa, K.; Nishida, S.; Kozaki, S.; Sakaguchi, S. Sporulation and C2 toxin production by Clostridium botulinum type C strains producing no C1 toxin. Microbiol. Immunol. 1978, 22, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Licona-Cassani, C.; Steen, J.A.; Zaragoza, N.E.; Moonen, G.; Moutafis, G.; Hodson, M.P.; Power, J.; Nielsen, L.K.; Marcellin, E. Tetanus toxin production is triggered by the transition from amino acid consumption to peptides. Anaerobe 2016, 41, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitmer, M.E.; Johnson, E.A. Development of improved defined media for Clostridium botulinum serotypes A, B, and E. Appl. Environ. Microbiol. 1988, 54, 753–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrigues, L.; Do, T.D.; Bideaux, C.; Guillouet, S.E.; Meynial-Salles, I. Insights into Clostridium tetani: From genome to bioreactors. Biotechnol. Adv. 2021, 54, 107781. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, M.; Dineen, S.S.; Maks, N.D.; Johnson, E.A. Regulation of neurotoxin complex expression in Clostridium botulinum strains 62A, Hall A-hyper, and NCTC2916. Anaerobe 2004, 10, 321–333. [Google Scholar] [CrossRef]
- Orellana, C.A.; Zaragoza, N.E.; Licona-Cassani, C.; Palfreyman, R.W.; Cowie, N.; Moonen, G.; Moutafis, G.; Power, J.; Nielsen, L.K.; Marcellin, E. Time-course transcriptomics reveals that amino acids catabolism plays a key role in toxinogenesis and morphology in Clostridium tetani. J. Ind. Microbiol. Biotechnol. 2020, 47, 1059–1073. [Google Scholar] [CrossRef]
- Fredrick, C.M.; Lin, G.; Johnson, E.A. Regulation of Botulinum Neurotoxin Synthesis and Toxin Complex Formation by Arginine and Glucose in Clostridium botulinum ATCC 3502. Appl. Environ. Microbiol. 2017, 83, e00642-17. [Google Scholar] [CrossRef] [Green Version]
- Inzalaco, H.N.; Tepp, W.H.; Fredrick, C.; Bradshaw, M.; Johnson, E.A.; Pellett, S. Posttranslational Regulation of Botulinum Neurotoxin Production in Clostridium botulinum Hall A-hyper. mSphere 2021, 6, e0032821. [Google Scholar] [CrossRef]
- Patterson-Curtis, S.I.; Johnson, E.A. Regulation of neurotoxin and protease formation in Clostridium botulinum Okra B and Hall A by arginine. Appl. Environ. Microbiol. 1989, 55, 1544–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, J.H.; Miller, P.A. Essential role of histidine peptides in tetanus toxin production. J. Biol. Chem. 1956, 223, 185–194. [Google Scholar] [CrossRef]
- Porfirio, Z.; Prado, S.M.; Vancetto, M.D.C.; Fratelli, F.; Alves, E.W.; Raw, I.; Fernandes, B.L.; Camargo, A.C.M.; Lebrun, I. Specific peptides of casein pancreatic digestion enhance the production of tetanus toxin. J. Appl. Microbiol. 1997, 83, 678–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brüggemann, H.; Gottschalk, G. Insights in metabolism and toxin production from the complete genome sequence of Clostridium tetani. Anaerobe 2004, 10, 53–68. [Google Scholar] [CrossRef]
- Fratelli, F.; Siquini, T.J.; Prado, S.M.; Higashi, H.G.; Converti, A.; de Carvalho, J.C. Effect of medium composition on the production of tetanus toxin by Clostridium tetani. Biotechnol. Prog. 2005, 21, 756–761. [Google Scholar] [CrossRef]
- Artin, I.; Carter, A.T.; Holst, E.; Lovenklev, M.; Mason, D.R.; Peck, M.W.; Radstrom, P. Effects of carbon dioxide on neurotoxin gene expression in nonproteolytic Clostridium botulinum Type E. Appl. Environ. Microbiol. 2008, 74, 2391–2397. [Google Scholar] [CrossRef] [Green Version]
- Lovenklev, M.; Artin, I.; Hagberg, O.; Borch, E.; Holst, E.; Radstrom, P. Quantitative interaction effects of carbon dioxide, sodium chloride, and sodium nitrite on neurotoxin gene expression in nonproteolytic Clostridium botulinum type B. Appl. Environ. Microbiol. 2004, 70, 2928–2934. [Google Scholar] [CrossRef] [Green Version]
- Artin, I.; Mason, D.R.; Pin, C.; Schelin, J.; Peck, M.W.; Holst, E.; Radstrom, P.; Carter, A.T. Effects of carbon dioxide on growth of proteolytic Clostridium botulinum, its ability to produce neurotoxin, and its transcriptome. Appl. Environ. Microbiol. 2010, 76, 1168–1172. [Google Scholar] [CrossRef] [Green Version]
- Chekabab, S.M.; Harel, J.; Dozois, C.M. Interplay between genetic regulation of phosphate homeostasis and bacterial virulence. Virulence 2014, 5, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Lamarche, M.G.; Wanner, B.L.; Crépin, S.; Harel, J. The phosphate regulon and bacterial virulence: A regulatory network connecting phosphate homeostasis and pathogenesis. FEMS Microbiol. Rev. 2008, 32, 461–473. [Google Scholar] [CrossRef] [Green Version]
- Pennings, J.L.A.; Abachin, E.; Esson, R.; Hodemaekers, H.; Francotte, A.; Claude, J.B.; Vanhee, C.; Uhlrich, S.; Vandebriel, R.J. Regulation of Clostridium tetani Neurotoxin Expression by Culture Conditions. Toxins 2022, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Siegel, L.S.; Metzger, J.F. Effect of fermentation conditions on toxin production by Clostridium botulinum type B. Appl. Environ. Microbiol. 1980, 40, 1023–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, M.W.; Stringer, S.C.; Carter, A.T. Clostridium botulinum in the post-genomic era. Food Microbiol. 2011, 28, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Korkeala, H.; Linden, J.; Lindström, M. Quantitative real-time reverse transcription-PCR analysis reveals stable and prolonged neurotoxin cluster gene activity in a Clostridium botulinum type E strain at refrigeration temperature. Appl. Environ. Microbiol. 2008, 74, 6132–6137. [Google Scholar] [CrossRef] [Green Version]
- Mascher, G.; Derman, Y.; Kirk, D.G.; Palonen, E.; Lindstrôm, M.; Korkeala, H. The CLO3403/CLO3404 two-component system of Clostridium botulinum E1 Beluga is important for cold shock response and growth at low temperatures. Appl. Environ. Microbiol. 2014, 80, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Derman, Y.; Isokallio, M.; Lindström, M.; Korkeala, H. The two-component system CBO2306/CBO2307 is important for cold adaptation of Clostridium botulinum ATCC 3502. Int. J. Food Microbiol. 2013, 167, 87–91. [Google Scholar] [CrossRef]
- Dahlsten, E.; Zhang, Z.; Somervuo, P.; Minton, N.P.; Lindström, M.; Korkeala, H. The cold-induced two-component system CBO0366/CBO0365 regulates metabolic pathways with novel roles in group I Clostridium botulinum ATCC 3502 cold tolerance. Appl. Environ. Microbiol. 2014, 80, 306–319. [Google Scholar] [CrossRef] [Green Version]
- Papenfort, K.; Vogel, J. Regulatory RNA in bacterial pathogens. Cell Host Microbe 2010, 8, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Westermann, A.J. Regulatory RNAs in Virulence and Host-Microbe Interactions. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Gripenland, J.; Netterling, S.; Loh, E.; Tiensuu, T.; Toledo-Arana, A.; Johansson, J. RNAs: Regulators of bacterial virulence. Nat. Rev. Microbiol. 2010, 8, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Waters, L.S.; Storz, G. Regulatory RNAs in bacteria. Cell 2009, 136, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheah, H.L.; Raabe, C.A.; Lee, L.P.; Rozhdestvensky, T.S.; Citartan, M.; Ahmed, S.A.; Tang, T.H. Bacterial regulatory RNAs: Complexity, function, and putative drug targeting. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 335–355. [Google Scholar] [CrossRef] [PubMed]
- Romby, P.; Charpentier, E. An overview of RNAs with regulatory functions in gram-positive bacteria. Cell Mol. Life Sci. 2010, 67, 217–237. [Google Scholar] [CrossRef] [PubMed]
- Brosse, A.; Guillier, M. Bacterial Small RNAs in Mixed Regulatory Networks. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Jones, A.J.; Fast, A.G.; Clupper, M.; Papoutsakis, E.T. Small and Low but Potent: The Complex Regulatory Role of the Small RNA SolB in Solventogenesis in Clostridium acetobutylicum. Appl. Environ. Microbiol. 2018, 84, e00597-18. [Google Scholar] [CrossRef] [Green Version]
- Venkataramanan, K.P.; Jones, S.W.; McCormick, K.P.; Kunjeti, S.G.; Ralston, M.T.; Meyers, B.C.; Papoutsakis, E.T. The Clostridium small RNome that responds to stress: The paradigm and importance of toxic metabolite stress in C. acetobutylicum. BMC Genom. 2013, 14, 849. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, H.; Lang, N.; Zhang, L.; Chai, C.; He, H.; Jiang, W.; Gu, Y. The Small RNA sr8384 Is a Crucial Regulator of Cell Growth in Solventogenic Clostridia. Appl. Environ. Microbiol. 2020, 86, e00665-20. [Google Scholar] [CrossRef]
- Ohtani, K.; Shimizu, T. Regulation of Toxin Production in Clostridium perfringens. Toxins 2016, 8, 207. [Google Scholar] [CrossRef]
- Soutourina, O. RNA-based control mechanisms of Clostridium difficile. Curr. Opin. Microbiol. 2017, 36, 62–68. [Google Scholar] [CrossRef]
- Kreis, V.; Soutourina, O. Clostridioides difficile—Phage relationship the RNA way. Curr. Opin. Microbiol. 2021, 66, 1–10. [Google Scholar] [CrossRef]
- Chen, Y.; Indurthi, D.C.; Jones, S.W.; Papoutsakis, E.T. Small RNAs in the genus Clostridium. mBio 2010, 2, e00340-10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brüggemann, H.; Chapeton-Montes, D.; Plourde, L.; Popoff, M.R. Identification of a non-coding RNA and its putative involvement in the regulation of tetanus toxin synthesis in Clostridium tetani. Sci. Rep. 2021, 11, 4157. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Hirakawa, H.; Tashiro, K.; Yoshizawa, S.; Kuhara, S.; Shimizu, T. Identification of a two-component VirR/VirS regulon in Clostridium perfringens. Anaerobe 2010, 16, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Abisado, R.G.; Benomar, S.; Klaus, J.R.; Dandekar, A.A.; Chandler, J.R. Bacterial Quorum Sensing and Microbial Community Interactions. mBio 2018, 9, e02331-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Payne, G.F.; Bentley, W.E. Quorum Sensing Communication: Molecularly Connecting Cells, Their Neighbors, and Even Devices. Annu. Rev. Chem. Biomol. Eng. 2020, 11, 447–468. [Google Scholar] [CrossRef] [PubMed]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus Virulence. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Zhao, L.; Montville, T.J.; Schaffner, D.W. Evidence for quorum sensing in Clostridium botulinum 56A. Lett. Appl. Microbiol. 2006, 42, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Cooksley, C.M.; Davis, I.J.; Winzer, K.; Chan, W.C.; Peck, M.W.; Minton, N.P. Regulation of neurotoxin production and sporulation by a Putative agrBD signaling system in proteolytic Clostridium botulinum. Appl. Environ. Microbiol. 2010, 76, 4448–4460. [Google Scholar] [CrossRef] [Green Version]
- Ihekwaba, A.E.; Mura, I.; Walshaw, J.; Peck, M.W.; Barker, G.C. An Integrative Approach to Computational Modelling of the Gene Regulatory Network Controlling Clostridium botulinum Type A1 Toxin Production. PLoS Comput. Biol. 2016, 12, e1005205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterne, M.; Wentzel, L.M. A new method for the large-scale production of high-titre botulinum formol-toxoid types C and D. J. Immunol. 1950, 65, 175–183. [Google Scholar]
- Koch, W.; Kaplan, D. A simple method for obtaining highly potent tetanus toxin. J. Immunol. 1953, 70, 1–5. [Google Scholar] [PubMed]
- Vraný, B.; Hnátková, Z.; Lettl, A. Production of toxic antigens in dialyzed cultures of microorganisms. Folia Microbiol. 1988, 33, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Wentzel, L.M.; Sterne, M.; Polson, A. High toxicity of pure botulinum type D toxin. Nature 1950, 166, 739–740. [Google Scholar] [CrossRef] [PubMed]
- Rasetti-Escargueil, C.; Lemichez, E.; Popoff, M.R. Public Health Risk Associated with Botulism as Foodborne Zoonoses. Toxins 2019, 12, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabritz, H.A.; Hill, K.K.; Barash, J.R.; Ticknor, L.O.; Helma, C.H.; Dover, N.; Payne, J.R.; Arnon, S.S. Molecular Epidemiology of Infant Botulism in California and Elsewhere, 1976-2010. J. Infect. Dis. 2014, 210, 1711–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.R.; Li, B.; Read, D. Botulinum versus tetanus neurotoxins: Why is botulinum neurotoxin but not tetanus neurotoxin a food poison? Toxicon 1995, 33, 1541–1547. [Google Scholar] [CrossRef]
- Bruggemann, H.; Gottschalk, G. Comparative genomics of clostridia: Link between the ecological niche and cell surface properties. Ann. N. Y. Acad. Sci. 2008, 1125, 73–81. [Google Scholar] [CrossRef]
- Mansfield, M.J.; Doxey, A.C. Genomic insights into the evolution and ecology of botulinum neurotoxins. Pathog. Dis. 2018, 76, 4978416. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Genetic organization of neurotoxin genes in representative Clostridium botulinum and Clostridium tetani strains. red, neurotoxin genes; orange, ntnh genes; purple, regulatory genes; green, ha genes; cyan, orfX and p47 genes.
Figure 1.
Genetic organization of neurotoxin genes in representative Clostridium botulinum and Clostridium tetani strains. red, neurotoxin genes; orange, ntnh genes; purple, regulatory genes; green, ha genes; cyan, orfX and p47 genes.

Figure 2.
Two-component systems (TCSs) involved in neurotoxin gene regulation in Clostridium botulinum strain Hall and ATCC3502 and in Clostridium tetani strain E88.
Figure 2.
Two-component systems (TCSs) involved in neurotoxin gene regulation in Clostridium botulinum strain Hall and ATCC3502 and in Clostridium tetani strain E88.

Figure 3.
Schematic representation of the regulatory pathways in Clostridium botulinum and Clostridium tetani. TCS, two-component system; AIP, autoinducing peptide. External factors act through TCSs and/or other unknown receptors or transporters (blue arrows).
Figure 3.
Schematic representation of the regulatory pathways in Clostridium botulinum and Clostridium tetani. TCS, two-component system; AIP, autoinducing peptide. External factors act through TCSs and/or other unknown receptors or transporters (blue arrows).


{kind=link}
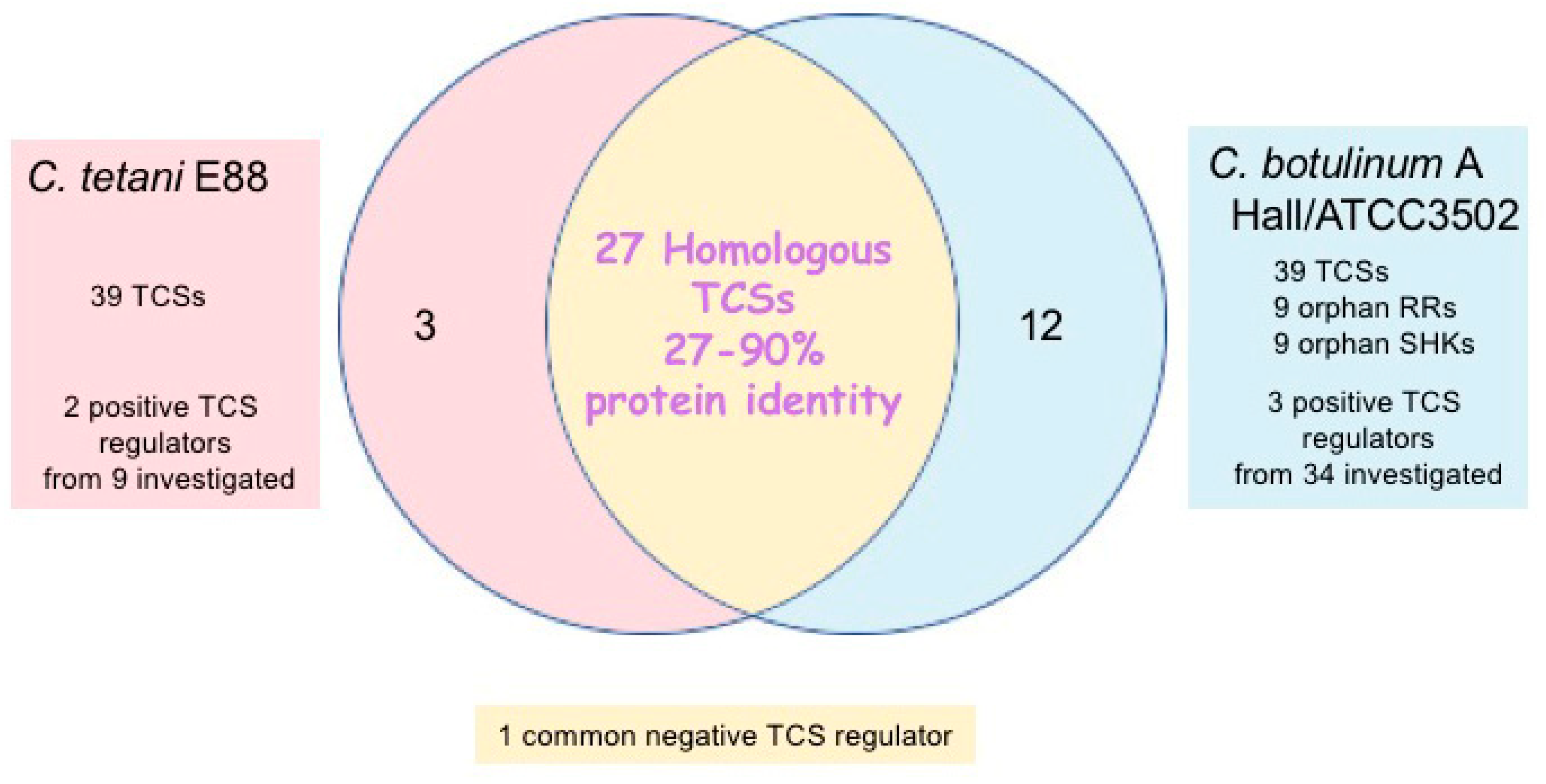{kind=link}
{kind=link}
Table 1.
Two-component systems involved in toxin gene regulation in C. botulinum Hall and C. tetani E88.
Table 1.
Two-component systems involved in toxin gene regulation in C. botulinum Hall and C. tetani E88.
| C. tetani E88 | C. botulinum Hall | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Genetic Localization | Locus Tag | Role | Family RR | Regulation of TeNT Synthesis | Ref. | Homolog | Protein Identity (RR) | Regulation of BoNT Synthesis | Refs. |
| chr | CTC_RS10150 CTC_RS10155 | SHK RR | LytR/AlgR | Positive | [32] | CLC_3250 CLC_3251 | 55% | None | [50] |
| plasmid | CTC_RS13810 CTC_RS13805 | SHK RR | OmpR | Positive | [32] | CLC_1431 CLC_1432 | 56% | None | [50] |
| No homolog | OmpR | CLC_1093 CLC_1094 | Positive | [50] | |||||
| No homolog | OmpR | CLC_1913 CLC_1914 | Positive | [50] | |||||
| chr | CTC_RS02080 CTC_RS02085 | RR SHK | OmpR | None | [32] | CLC_0661 CLC_0663 | 65% | Positive | [50] |
| chr | CTC_RS10030 CTC_RS10035 | SHK RR | OmpR | None | [32] | CLC_0410 CLC_0411 | 68% | Cell wall alteration | [50] |
| No homolog | OmpR | CLC_3293 CLC_3294 | Cell wall alteration | [50] | |||||
| chr | CTC_RS07310 CTC_RS07315 | SHK RR | OmpR | Negative | [32] | strain Hall CLC_0842 CLC_0843 | 58% | None | [50] |
| strain ATCC3502 CBO_0786 CBO_0787 | 100% | Negative | [49] | ||||||
chr, chromosome; RR, response regulator; SHK, sensor histidine kinase. positive effects are in green, and negative effects are in pink.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Popoff, M.R.; Brüggemann, H. Regulatory Networks Controlling Neurotoxin Synthesis in Clostridium botulinum and Clostridium tetani. Toxins 2022, 14, 364. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14060364
AMA Style
Popoff MR, Brüggemann H. Regulatory Networks Controlling Neurotoxin Synthesis in Clostridium botulinum and Clostridium tetani. Toxins. 2022; 14(6):364. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14060364
Chicago/Turabian StylePopoff, Michel R., and Holger Brüggemann. 2022. "Regulatory Networks Controlling Neurotoxin Synthesis in Clostridium botulinum and Clostridium tetani" Toxins 14, no. 6: 364. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14060364
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.