MHC Class II and Non-MHC Class II Genes Differentially Influence Humoral Immunity to Bacillus anthracis Lethal Factor and Protective Antigen

Abstract
:1. Introduction
2. Results and Discussion
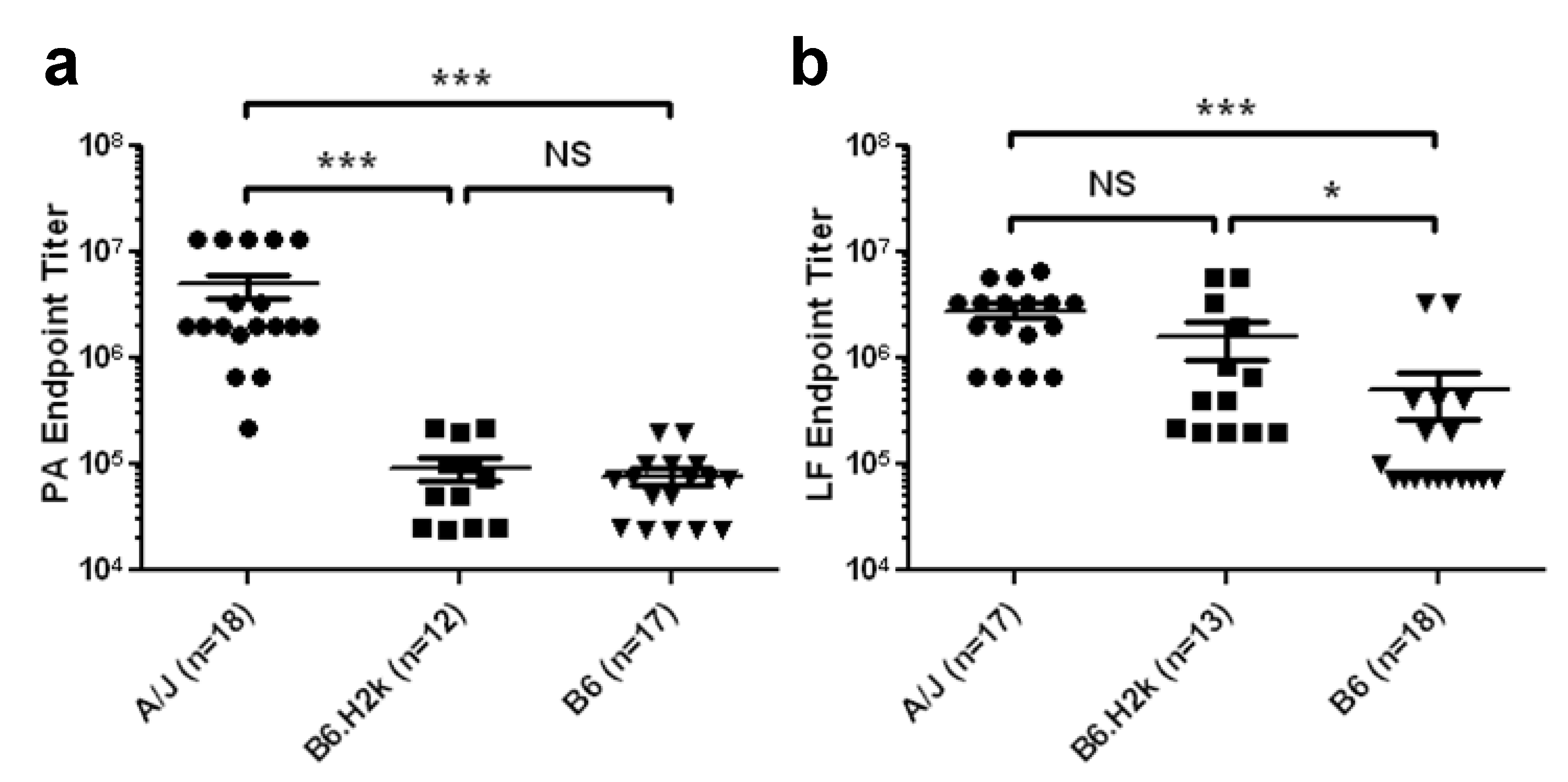
2.1. Magnitude of Serum LF IgG Response to Vaccination Is More Dependent on MHC Class II than Magnitude of Serum PA IgG Response
2.2. Kinetics and Breadth of Response to PA Are Background Strain-Dependent

{kind=link}
{kind=link}
{kind=link}
| Epitope Number | Decapeptide number | Sequence | Amino acid number | Domain | Reactive strain |
|---|---|---|---|---|---|
| 1 | 36–38 | DLSIPSSELENIPSENQY | 71–84 | I | A/J, B6, B6.H2k (Common) |
| 2 | 47–48 | IWSGFIKVKKSD | 93–104 | I | A/J (Background) |
| 3 | 75–77 | YQRENPTEKGLDFK | 149–162 | I | B6 (MHC II) |
| 4 | 97–98 | RKKRSTSAGPTV | 193–204 | I' | A/J, B6 |
| 5 | 110–111 | EGYTVDVKNKRT | 219–230 | I' | B6 (MHC II) |
| 6 | 120–122 | IHEKKGLTKYKSSP | 239–252 | I' | A/J, B6 |
| 7 | 130–132 | SDPYSDFEKVTGRI | 259–272 | I' | A/J, B6 |
| 8 | 170–171 | HASFFDIGGSVS | 339–350 | II | A/J (Background) |
| 9 | 287–289 | GKDITEFDFNFDQQ | 573–586 | III | A/J (Background) |
| 10 | 320–322 | SVVKEAHREVINSS | 639–652 | IV | A/J, B6, B6.H2k (Common) |
| 11 | 329–330 | LLLNIDKDIRKIL | 657–668 | IV | B6.H2k |
| 12 | 357–358 | KLPLYISNPNYK | 713–724 | IV | A/J (Background) |
2.3. Kinetics and Breadth of Response to LF Are Background Strain-Dependent

| Epitope number | Decapeptide number | Sequence | Amino acid number | Domain | Reactive strain |
|---|---|---|---|---|---|
| 1 | 19–20 | HGDVGMHVKEKE | 37–48 | I | A/J, B6 |
| 2 | 24–25 | KEKNKDENKRKD | 47–58 | I | B6, B6.H2k (Background) |
| 3 | 31–34 | RNKTQEEHLKEIMKHI | 61–76 | I | A/J (Background) |
| 4 | 49–51 | EKVPSDVLEMYK | 97–110 | I | A/J, B6.H2k (MHCII) |
| 5 | 66–70 | SEDKKKIKDIYGKDALLH | 131–148 | I | A/J (Background) |
| 6 | 74–75 | LHEHYVYAKEGY | 147–158 | I | A/J (Background) |
| 7 | 133–135 | VLQLYAPEAFNYMD | 265–278 | I | A/J, B6, B6.H2k (Common) |
| 8 | 149–151 | MLARYEKWEKIKQH | 297–310 | II | A/J (Background) |
| 9 | 172–174 | SLSQEEKELLKRIQ | 343–356 | III | A/J, B6, B6.H2k (Common) |
| 10 | 182–184 | LSTEEKEFLKKLQI | 363–376 | III | A/J, B6, B6.H2k (Common) |
| 11 | 201–203 | LSEKEKEFLKKLKL | 401–414 | III | A/J, B6.H2k (MHCII) |
| 12 | 209–210 | QPYDINQRLQDT | 417–428 | III/II | A/J (Background) |
| 13 | 216–218 | LIDSPSINLDVRKQ | 431–444 | II | A/J (Background) |
| 14 | 315–316 | VESAYLILNEWK | 629–640 | IV | A/J (Background) |
| 15 | 324–325 | LIKKVTNYLVDG | 647–658 | IV | B6.H2k |
| 16 | 328–331 | LVDGNGRFVFTDITLP | 655–670 | IV | A/J (Background) |
| 17 | 336–339 | NIAEQYTHQDEIYEQV | 671–686 | IV | A/J (Background) |
| 18 | 347–348 | VPESRSILLHGP | 693–704 | IV | B6.H2k |
| 19 | 358–359 | EGFIHEFGHAVD | 715–726 | IV | A/J (Background) |
| 20 | 372–373 | NSKKFIDIFKEE | 743–754 | IV | A/J (Background) |
| 21 | 391–394 | DHAERLKVQKNAPKTF | 781–796 | IV | A/J (Background) |
| 22 | 397–398 | PKTFQFINDQIK | 793–804 | IV | A/J (Background) |
2.4. Fine Specificity of the IgG Response to LF Is Strongly Influenced by Genetic Background
| Epitope Number | Decapeptide number | Sequence | Amino acid number | Reactive strain |
|---|---|---|---|---|
| 3 | 75–77 | YQRENPTEKGLD | 120–131 | B6.H2k, B6 (Background) |
| 9 | 287–290 | GKDITEFDFNFDQQTS | 544–559 | A/J (Background) |
| 12 | 357–358 | KLPLYISNPNYK | 684–695 | A/J (Background) |
| 3/15 (20%) | ||||
| 1 | 35–38 | TGDLSIPSSELENIPS | 40–51 | A/J, B6, B6.H2k (Common) |
| 2 | 47–48 | IWSGFIKVKKSD | 64–75 | A/J, B6, B6.H2k (Common) |
| 4 | 93–97 | LKQKSSNSRKKRSTSAGP | 156–173 | A/J, B6, B6.H2k (Common) |
| 6 | 120–122 | IHEKKGLTKYKSSP | 210–223 | A/J, B6, B6.H2k (Common) |
| 7 | 130–131 | SDPYSDFEKVTG | 234–241 | A/J, B6, B6.H2k (Common) |
| C | 304–305 | LDKIKLNAKMNI | 578–589 | A/J, B6, B6.H2k (Common) |
| 10 | 320–322 | SVVKEAHREVINSS | 610–623 | A/J, B6, B6.H2k (Common) |
| 7/15 (47%) | ||||
| 5 | 110–111 | EGYTVDVKNKRT | 190–201 | B6 (MHCII) |
| 8 | 170–171 | HASFFDIGGSVS | 310–321 | A/J, B6.H2k (MHCII) |
| A | 208–210 | GKNQTLATIKAKEN | 388–399 | A/J, B6.H2k (MHCII) |
| B | 273–274 | TTKPDMTLKEAL | 516–527 | B6 (MHCII) |
| 11 | 328–329 | GLLLNIDKDIRK | 626–637 | A/J, B6.H2k (MHCII) |
| 5/15 (33%) | ||||
| D | 351–354 | GKTFIDFKKYNDKLPL | 672–687 | B6.H2k |
| Epitope number | Decapeptide number | Sequence | Amino acid number | Reactive strain |
|---|---|---|---|---|
| 1 | 19–20 | HGDVGMHVKEKE | 37–48 | B6.H2k, B6 (Background) |
| 2 | 23–25 | KEKEKNKDENKRKD | 45–58 | B6.H2k, B6 (Background) |
| 6 | 74–75 | LHEHYVYAKEGY | 147–158 | A/J (Background) |
| C | 145–146 | LEELKDQRMLAR | 289–300 | A/J (Background) |
| E | 191–193 | SLSEEEKELLNRIQ | 381–394 | B6.H2k, B6 (Background) |
| 11 | 201–203 | LSEKEKEFLKKLKL | 401–414 | B6.H2k, B6 (Background) |
| 12 | 209–210 | QPYDINQRLQDT | 417–428 | A/J (Background) |
| 20 | 372–373 | NSKKFIDIFKEE | 743–754 | A/J (Background) |
| 21 | 393–394 | RLKVQKNAPKTF | 785–796 | A/J (Background) |
| 9/17 (53%) | ||||
| 4 | 49–51 | EKVPSDVLEMYKAI | 97–110 | A/J, B6, B6.H2k (Common) |
| 7 | 134–135 | QLYAPEAFNYMD | 267–278 | A/J, B6, B6.H2k (Common) |
| 9 | 172–174 | SLSQEEKELLKRIQ | 343–356 | A/J, B6, B6.H2k (Common) |
| 10 | 182–183 | LSTEEKEFLKKL | 363–374 | A/J, B6, B6.H2k (Common) |
| 4/17 (23%) | ||||
| A | 109–110 | DGQDLLFTNQLK | 217–228 | B6 (MHCII) |
| B | 141–142 | NEQEINLSLEEL | 281–292 | B6 (MHCII) |
| 16 | 330–331 | NGRFVFTDITLP | 659–670 | A/J, B6.H2k (MHCII) |
| 17 | 336–339 | NIAEQYTHQDEIYEQV | 671–686 | A/J, B6.H2k (MHCII) |
| 4/17 (23%) | ||||
| 3 | 32–33 | KTQEEHLKEIMK | 63–74 | B6.H2k |
| 5 | 66–69 | SEDKKKIKDIYGKD | 131–146 | A/J, B6 |
| D | 167–168 | IEPKKDDIIHSL | 333–344 | B6.H2k |
| 13 | 216–217 | LIDSPSINLDVR | 431–442 | A/J, B6 |
| 18 | 347–348 | VPESRSILLHGP | 693–704 | B6.H2k |
2.5. Discussion
3. Experimental Section
3.1. Animal Use
3.2. Production of Recombinant(r)PA and rLF Proteins
3.3. Immunization and Blood Sampling
3.4. Recombinant Protein ELISAs
3.5. Solid Phase Epitope Mapping
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Keim, P.; Smith, K.L.; Keys, C.; Takahashi, H.; Kurata, T.; Kaufmann, A. Molecular investigation of the aum shinrikyo anthrax release in Kameido, Japan. J. Clin. Microbiol. 2001, 39, 4566–4567. [Google Scholar] [CrossRef]
- Blendon, R.J.; Benson, J.M.; DesRoches, C.M.; Pollard, W.E.; Parvanta, C.; Herrmann, M.J. The impact of anthrax attacks on the American public. MedGenMed 2002, 4, 1. [Google Scholar]
- Dixon, T.C.; Meselson, M.; Guillemin, J.; Hanna, P.C. Anthrax. N. Engl. J. Med. 1999, 341, 815–826. [Google Scholar] [CrossRef]
- Brossier, F.; Weber-Levy, M.; Mock, M.; Sirard, J.C. Role of toxin functional domains in anthrax pathogenesis. Infect. Immun. 2000, 68, 1781–1786. [Google Scholar] [CrossRef]
- Reuveny, S.; White, M.D.; Adar, Y.Y.; Kafri, Y.; Altboum, Z.; Gozes, Y.; Kobiler, D.; Shafferman, A.; Velan, B. Search for correlates of protective immunity conferred by anthrax vaccine. Infect. Immun. 2001, 69, 2888–2893. [Google Scholar] [CrossRef]
- Marcus, H.; Danieli, R.; Epstein, E.; Velan, B.; Shafferman, A.; Reuveny, S. Contribution of immunological memory to protective immunity conferred by a Bacillus anthracis protective antigen-based vaccine. Infect. Immun. 2004, 72, 3471–3477. [Google Scholar] [CrossRef]
- Turnbull, P.C. Anthrax vaccines: Past, present and future. Vaccine 1991, 9, 533–539. [Google Scholar] [CrossRef]
- Weiss, S.; Kobiler, D.; Levy, H.; Marcus, H.; Pass, A.; Rothschild, N.; Altboum, Z. Immunological correlates for protection against intranasal challenge of Bacillus anthracis spores conferred by a protective antigen-based vaccine in rabbits. Infect. Immun. 2006, 74, 394–398. [Google Scholar] [CrossRef]
- Chitlaru, T.; Altboum, Z.; Reuveny, S.; Shafferman, A. Progress and novel strategies in vaccine development and treatment of anthrax. Immunol. Rev. 2011, 239, 221–236. [Google Scholar] [CrossRef]
- Albrecht, M.T.; Li, H.; Williamson, E.D.; LeButt, C.S.; Flick-Smith, H.C.; Quinn, C.P.; Westra, H.; Galloway, D.; Mateczun, A.; Goldman, S.; et al. Human monoclonal antibodies against anthrax lethal factor and protective antigen act independently to protect against Bacillus anthracis infection and enhance endogenous immunity to anthrax. Infect. Immun. 2007, 75, 5425–5433. [Google Scholar]
- Brossier, F.; Levy, M.; Landier, A.; Lafaye, P.; Mock, M. Functional analysis of Bacillus anthracis protective antigen by using neutralizing monoclonal antibodies. Infect. Immun. 2004, 72, 6313–6317. [Google Scholar] [CrossRef]
- Chen, Z.; Moayeri, M.; Crown, D.; Emerson, S.; Gorshkova, I.; Schuck, P.; Leppla, S.H.; Purcell, R.H. Novel chimpanzee/human monoclonal antibodies that neutralize anthrax lethal factor, and evidence for possible synergy with anti-protective antigen antibody. Infect. Immun. 2009, 77, 3902–3908. [Google Scholar]
- Nguyen, M.L.; Crowe, S.R.; Kurella, S.; Teryzan, S.; Cao, B.; Ballard, J.D.; James, J.A.; Farris, A.D. Sequential B-cell epitopes of Bacillus anthracis lethal factor bind lethal toxin-neutralizing antibodies. Infect. Immun. 2009, 77, 162–169. [Google Scholar] [CrossRef]
- Staats, H.F.; Alam, S.M.; Scearce, R.M.; Kirwan, S.M.; Zhang, J.X.; Gwinn, W.M.; Haynes, B.F. In vitro and in vivo characterization of anthrax anti-protective antigen and anti-lethal factor monoclonal antibodies after passive transfer in a mouse lethal toxin challenge model to define correlates of immunity. Infect. Immun. 2007, 75, 5443–5452. [Google Scholar] [CrossRef]
- Zhao, P.; Liang, X.; Kalbfleisch, J.; Koo, H.M.; Cao, B. Neutralizing monoclonal antibody against anthrax lethal factor inhibits intoxication in a mouse model. Hum. Antibodies 2003, 12, 129–135. [Google Scholar]
- Little, S.F.; Leppla, S.H.; Friedlander, A.M. Production and characterization of monoclonal antibodies against the lethal factor component of Bacillus anthracis lethal toxin. Infect. Immun. 1990, 58, 1606–1613. [Google Scholar]
- Crowe, S.R.; Ash, L.L.; Engler, R.J.; Ballard, J.D.; Harley, J.B.; Farris, A.D.; James, J.A. Select human anthrax protective antigen epitope-specific antibodies provide protection from lethal toxin challenge. J. Infect. Dis. 2010, 202, 251–260. [Google Scholar] [CrossRef]
- Crowe, S.R.; Garman, L.; Engler, R.J.; Farris, A.D.; Ballard, J.D.; Harley, J.B.; James, J.A. Anthrax vaccination induced anti-lethal factor IgG: Fine specificity and neutralizing capacity. Vaccine 2011, 29, 3670–3678. [Google Scholar]
- Reason, D.C.; Ullal, A.; Liberato, J.; Sun, J.; Keitel, W.; Zhou, J. Domain specificity of the human antibody response to Bacillus anthracis protective antigen. Vaccine 2008, 26, 4041–4047. [Google Scholar] [CrossRef]
- Mohamed, N.; Li, J.; Ferreira, C.S.; Little, S.F.; Friedlander, A.M.; Spitalny, G.L.; Casey, L.S. Enhancement of anthrax lethal toxin cytotoxicity: A subset of monoclonal antibodies against protective antigen increases lethal toxin-mediated killing of murine macrophages. Infect. Immun. 2004, 72, 3276–3283. [Google Scholar]
- Pajewski, N.M.; Parker, S.D.; Poland, G.A.; Ovsyannikova, I.G.; Song, W.; Zhang, K.; McKinney, B.A.; Pankratz, V.S.; Edberg, J.C.; Kimberly, R.P.; et al. The role of HLA-DR-DQ haplotypes in variable antibody responses to Anthrax Vaccine Adsorbed. Genes Immun. 2011, 12, 457–465. [Google Scholar] [CrossRef]
- Pajewski, N.M.; Shrestha, S.; Quinn, C.P.; Parker, S.D.; Wiener, H.; Aissani, B.; McKinney, B.A.; Poland, G.A.; Edberg, J.C.; Kimberly, R.P.; et al. A genome-wide association study of host genetic determinants of the antibody response to anthrax vaccine adsorbed. Vaccine 2012, 30, 4778–4784. [Google Scholar]
- Harvill, E.T.; Lee, G.; Grippe, V.K.; Merkel, T.J. Complement depletion renders c57bl/6 mice sensitive to the Bacillus anthracis Sterne strain. Infect. Immun. 2005, 73, 4420–4422. [Google Scholar] [CrossRef]
- James, J.A.; Harley, J.B. B-cell epitope spreading in autoimmunity. Immunol. Rev. 1998, 164, 185–200. [Google Scholar]
- James, J.A.; Harley, J.B. A model of peptide-induced lupus autoimmune B cell epitope spreading is strain specific and is not H-2 restricted in mice. J. Immunol. 1998, 160, 502–508. [Google Scholar]
- Milich, D.R.; Chisari, F.V. Genetic regulation of the immune response to hepatitis B surface antigen (HBsAg). I. H-2 restriction of the murine humoral immune response to the a and d determinants of HBsAg. J. Immunol. 1982, 129, 320–325. [Google Scholar]
- Else, K.; Wakelin, D. Genetic variation in the humoral immune responses of mice to the nematode Trichuris muris. Parasite Immunol. 1989, 11, 77–90. [Google Scholar] [CrossRef]
- Reason, D.; Liberato, J.; Sun, J.; Keitel, W.; Zhou, J. Frequency and domain specificity of toxin-neutralizing paratopes in the human antibody response to Anthrax Vaccine Adsorbed. Infect. Immun. 2009, 77, 2030–2035. [Google Scholar] [CrossRef]
- Chen, Z.; Moayeri, M.; Purcell, R. Monoclonal antibody therapies against anthrax. Toxins 2011, 3, 1004–1019. [Google Scholar] [CrossRef]
- Nguyen, M.L.; Terzyan, S.; Ballard, J.D.; James, J.A.; Farris, A.D. The major neutralizing antibody responses to recombinant anthrax lethal and edema factors are directed to non-cross-reactive epitopes. Infect. Immun. 2009, 77, 4714–4723. [Google Scholar] [CrossRef]
- Dumas, E.K.; Nguyen, M.L.; Cox, P.M.; Rodgers, H.; Peterson, J.L.; James, J.A.; Farris, A.D. Stochastic humoral immunity to Bacillus anthracis protective antigen: Identification of anti-peptide IgG correlating with seroconversion to lethal toxin neutralization. 2011; Manuscript Submitted. [Google Scholar]
- Abboud, N.; Casadevall, A. Immunogenicity of Bacillus anthracis protective antigen domains and efficacy of elicited antibody responses depend on host genetic background. Clin. Vaccine Immunol. 2008, 15, 1115–1123. [Google Scholar] [CrossRef]
- Abboud, N.; de Jesus, M.; Nakouzi, A.; Cordero, R.J.; Pujato, M.; Fiser, A.; Rivera, J.; Casadevall, A. Identification of linear epitopes in Bacillus anthracis protective antigen bound by neutralizing antibodies. J. Biol. Chem. 2009, 284, 25077–25086. [Google Scholar]
- Clement, K.H.; Rudge, T.L., Jr.; Mayfield, H.J.; Carlton, L.A.; Hester, A.; Niemuth, N.A.; Sabourin, C.L.; Brys, A.M.; Quinn, C.P. Vaccination of rhesus macaques with the anthrax vaccine adsorbed vaccine produces a serum antibody response that effectively neutralizes receptor-bound protective antigen in vitro. Clin. Vaccine Immunol. 2010, 17, 1753–1762. [Google Scholar] [CrossRef]
- Rivera, J.; Nakouzi, A.; Abboud, N.; Revskaya, E.; Goldman, D.; Collier, R.J.; Dadachova, E.; Casadevall, A. A monoclonal antibody to Bacillus anthracis protective antigen defines a neutralizing epitope in domain 1. Infect. Immun. 2006, 74, 4149–4156. [Google Scholar] [CrossRef]
- Oscherwitz, J.; Yu, F.; Cease, K.B. A synthetic peptide vaccine directed against the 2β2-2β3 loop of domain 2 of protective antigen protects rabbits from inhalation anthrax. J. Immunol. 2010, 185, 3661–3668. [Google Scholar] [CrossRef]
- Kaur, M.; Chug, H.; Singh, H.; Chandra, S.; Mishra, M.; Sharma, M.; Bhatnagar, R. Identification and characterization of immunodominant B-cell epitope of the c-terminus of protective antigen of Bacillus anthracis. Mol. Immunol. 2009, 46, 2107–2115. [Google Scholar] [CrossRef]
- Voth, D.E.; Hamm, E.E.; Nguyen, L.G.; Tucker, A.E.; Salles, I.I.; Ortiz-Leduc, W.; Ballard, J.D. Bacillus anthracis oedema toxin as a cause of tissue necrosis and cell type-specific cytotoxicity. Cell. Microbiol. 2005, 7, 1139–1149. [Google Scholar] [CrossRef]
- McClain, M.T.; Rapp, E.C.; Harley, J.B.; James, J.A. Infectious mononucleosis patients temporarily recognize a unique, cross-reactive epitope of Epstein-Barr virus nuclear antigen-1. J. Med. Virol. 2003, 70, 253–257. [Google Scholar] [CrossRef]
- James, J.A.; Scofield, R.H.; Harley, J.B. Basic amino acids predominate in the sequential autoantigenic determinants of the small nuclear 70k ribonucleoprotein. Scand. J. Immunol. 1994, 39, 557–566. [Google Scholar] [CrossRef]
- James, J.A.; Harley, J.B. Peptide autoantigenicity of the small nuclear ribonucleoprotein c. Clin. Exp. Rheumatol. 1995, 13, 299–305. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Garman, L.; Dumas, E.K.; Kurella, S.; Hunt, J.J.; Crowe, S.R.; Nguyen, M.L.; Cox, P.M.; James, J.A.; Farris, A.D. MHC Class II and Non-MHC Class II Genes Differentially Influence Humoral Immunity to Bacillus anthracis Lethal Factor and Protective Antigen. Toxins 2012, 4, 1451-1467. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins4121451
Garman L, Dumas EK, Kurella S, Hunt JJ, Crowe SR, Nguyen ML, Cox PM, James JA, Farris AD. MHC Class II and Non-MHC Class II Genes Differentially Influence Humoral Immunity to Bacillus anthracis Lethal Factor and Protective Antigen. Toxins. 2012; 4(12):1451-1467. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins4121451
Chicago/Turabian StyleGarman, Lori, Eric K. Dumas, Sridevi Kurella, Jonathan J. Hunt, Sherry R. Crowe, Melissa L. Nguyen, Philip M. Cox, Judith A. James, and A. Darise Farris. 2012. "MHC Class II and Non-MHC Class II Genes Differentially Influence Humoral Immunity to Bacillus anthracis Lethal Factor and Protective Antigen" Toxins 4, no. 12: 1451-1467. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins4121451