Requirement of Simultaneous Assessment of Crystal- and Supernatant-Related Entomotoxic Activities of Bacillus thuringiensis Strains for Biocontrol-Product Development

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
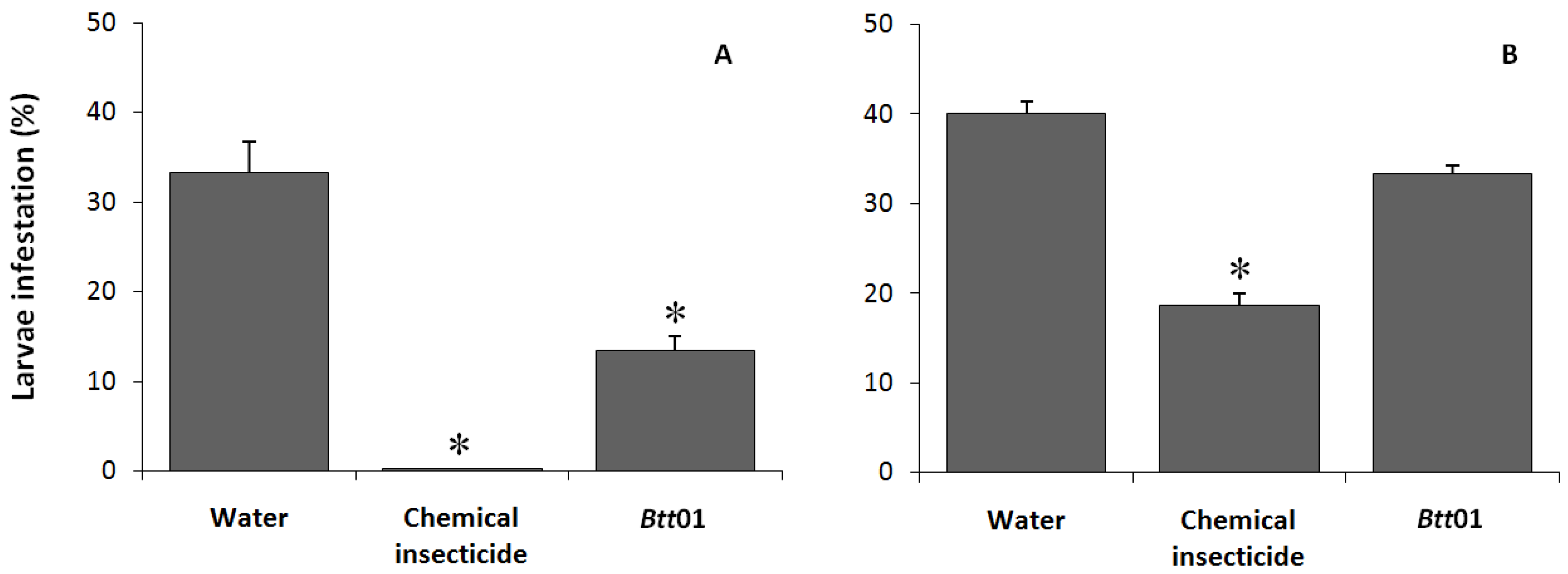
2.1. Entomotoxicity of Btt01 δ-Endotoxins in Laboratory and Field Bioassays

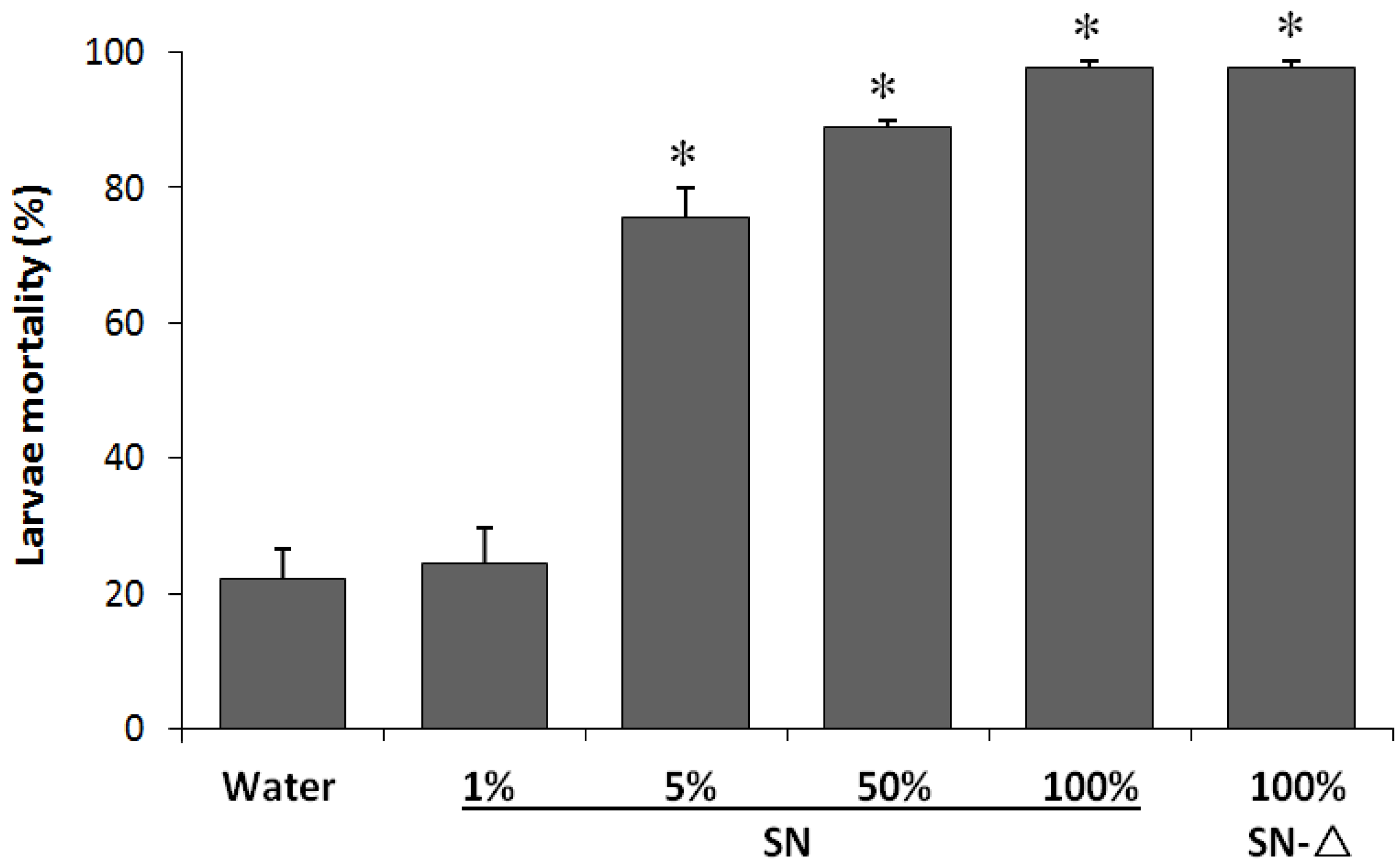
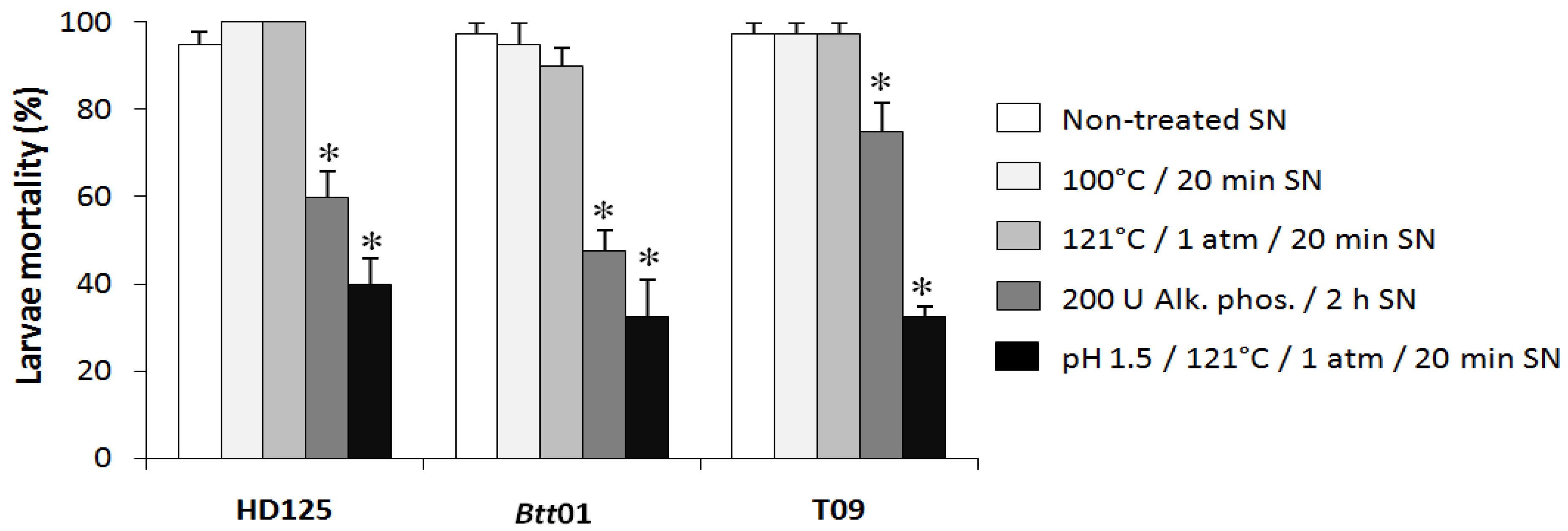
2.2. Entomotoxicity of Btt01 Supernatant



3. Discussion
4. Experimental Section
4.1. Bacillus Thuringiensis Strain and Culture Conditions
4.2. Feeding Bioassays with Endospores of Btt01 against Spodoptera frugiperda
4.3. Entomotoxic Activity of Btt01 Endospores in the Field
4.4. Feeding Bioassays with Supernatant of Btt01 against S. frugiperda
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aronson, A.I.; Beckman, W.; Dunn, P. Bacillus thuringiensis and related insect pathogens. Microbiol. Rev. 1986, 50, 1–24. [Google Scholar]
- Crickmore, N.; Zeigler, D.R.; Feitelson, J.; Schnepf, E.; van Rie, J.; Lereclus, D.; Baum, J.; Dean, D.H. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 807–813. [Google Scholar]
- Raymond, B.; Johnston, P.R.; Nielsen-LeRoux, C.; Lereclus, D.; Crickmore, N. Bacillus thuringiensis: An impotent pathogen? Trends Microbiol. 2010, 18, 189–194. [Google Scholar] [CrossRef]
- Schnepf, E.; Crickmore, N.; van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar]
- Zhao, J.Z.; Jun, C.; Collins, H.L.; Bates, S.L.; Roush, R.T.; Earle, E.D.; Shelton, A.M. Concurrent use of transgenic plants expressing a single and two Bacillus thuringiensis genes speeds insect adaptation to pyramided plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8426–8430. [Google Scholar]
- Kupke, T.; Lechner, M.; Kaim, G.; Gotz, F. Improved purification and biochemical properties of phosphatidylinositol-specific phospholipase C from Bacillus thuringiensis. Eur. J. Biochem. 1989, 185, 151–155. [Google Scholar] [CrossRef]
- Arora, N.; Ahmad, T.; Rajagopal, R.; Bhatnagar, R.K.A. Constitutively expressed 36 kDa exochitinase from Bacillus thuringiensis HD-1. Biochem. Biophys. Res. Commun. 2003, 307, 620–625. [Google Scholar] [CrossRef]
- Donovan, W.P.; Engleman, J.T.; Donovan, J.C.; Baum, J.A.; Bunkers, G.J.; Chi, D.J.; Clinton, W.P.; English, L.; Heck, G.R.; Ilagan, O.M.; et al. Discovery and characterization of Sip1A: A novel secreted protein from Bacillus thuringiensis with activity against coleopteran larvae. Appl. Microbiol. Biotechnol. 2006, 72, 713–719. [Google Scholar] [CrossRef]
- Estruch, J.J.; Warren, G.W.; Mullins, M.A.; Nye, G.J.; Craig, J.A.; Koziel, M.G. Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proc. Natl. Acad. Sci. USA 1996, 93, 5389–5394. [Google Scholar] [CrossRef]
- Farkas, J.; Sebesta, K.; Horkas, K.; Samek, Z.; Dolijs, J.; Storm, F. The structure of exotoxin of Bacillus thuringiensis var. gelechaiae. Collect. Czech. Chem. Commun. 1969, 34, 1118–1120. [Google Scholar] [CrossRef]
- Levinson, B.L.; Kasyan, K.J.; Chiu, S.S.; Currier, T.C.; Gonzalez, J.M., Jr. Identification of β-exotoxin production, plasmids encoding β-exotoxin, and a new exotoxin in Bacillus thuringiensis by using high-performance liquid chromatography. J. Bacteriol. 1990, 172, 3172–3179. [Google Scholar]
- Beebee, T.; Korner, A.; Bond, R.P. Differential inhibition of mammalian ribonucleic acid polymerases by an exotoxin from Bacillus thuringiensis: The direct observation of nucleoplasmic ribonucleic acid polymerase activity in intact nuclei. Biochem. J. 1972, 127, 619–634. [Google Scholar]
- Demarchi, M. Análise da Conjuntura Agropecuária, Safra 2011/12: Milho; Secretary of Agriculture and Food Supply—Department of Rural Economy: Paraná, Brazil, 2011. [Google Scholar]
- Cruz, I. A Lagarta-do-Cartucho na Cultura do Milho; 21 Technical Circular; Embrapa-CNPMS: Sete Lagoas-MG, Brazil, 1995. [Google Scholar]
- Loguercio, L.L.; Carneiro, N.P.; Carneiro, A.A. Milho Bt: Alternativa biotecnológica para controle biológico de insetos-praga. Biotecnolog. Ciênc. Desenvolv. 2002, 24, 46–52. [Google Scholar]
- Adamczyk, J.J., Jr.; Holloway, J.W.; Leonard, B.R.; Graves, J.B. Susceptibility of fall armyworm collected from different plant hosts to selected insecticides and transgenic Bt cotton. J. Cotton Sci. 1997, 1, 21–28. [Google Scholar]
- Hardke, J.T.; Temple, J.H.; Leonard, R.B.; Jackson, R.E. Laboratory toxicity and field efficacy of selected insecticides against fall armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 2011, 94, 272–278. [Google Scholar] [CrossRef]
- Tholkappian, C.; Rajendran, S. Pesticide application and its adverse impact on health: Evidences from kerala. Int. J. Sci. Technol. 2011, 1, 56–59. [Google Scholar]
- Sadek, M.M. Antifeedant and toxic activity of Adhatoda vasica leaf extract against Spodoptera littoralis (Lep., Noctuidae). J. Appl. Entomol. 2003, 127, 396–404. [Google Scholar] [CrossRef]
- Barreto, M.R.; Guimaraes, C.T.; Teixeira, F.F.; Paiva, E.; Valicente, F.H. Effect of Baculovirus spodoptera isolates in Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) larvae and their characterization by RAPD. Neotrop. Entomol. 2005, 34, 67–75. [Google Scholar] [CrossRef]
- Delfín-González, H.; Bojórquez-Acevedo, M.; Manrique-Saide, P. Parasitoids of fall armyworm (Lepidoptera: Noctuidae) from a traditional maize crop in the mexican state of yucatan. Fla. Entomol. 2007, 90, 759–761. [Google Scholar] [CrossRef]
- Santos, W.L.; Freire, M.G.M.; Bogorni, P.C.; Vendramim, J.D.; Macedo, M.L.R. Effect of the Aqueous extracts of the seeds of Talisia esculenta and Sapindus saponaria on fall armyworm. Braz. Arch. Biol. Technol. 2008, 51, 373–383. [Google Scholar]
- Arango, J.A.; Romero, M.; Orduz, S. Diversity of Bacillus thuringiensis strains from Colombia with insecticidal activity against Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Appl. Microbiol. 2002, 92, 466–474. [Google Scholar]
- Monnerat, R.; Martins, E.; Queiroz, P.; Ordúz, S.; Jaramillo, G.; Benintende, G.; Cozzi, J.; Real, M.D.; Martinez-Ramirez, A.; Rausell, C.; et al. Genetic variability of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) populations from Latin America is associated with variations in susceptibility to Bacillus thuringiensis Cry toxins. Appl. Environ. Microbiol. 2005, 77, 7029–7035. [Google Scholar]
- Valicente, F.H.; Lana, U.G.P. Molecular characterization of the Bacillus thuringiensis (Berliner) strains 344 and 1644, efficient against fall armyworm Spodoptera frugiperda (J.E. Smith). Milho Sorgo 2008, 7, 195–209. [Google Scholar]
- Loguercio, L.L.; Santos, C.G.; Barreto, M.R.; Guimarães, C.T.; Paiva, E. Association of PCR and feeding bioassays as a large-scale method to screen tropical Bacillus thuringiensis isolates for a cry constitution with higher insecticidal effect against Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae. Lett. Appl. Microbiol. 2001, 32, 362–367. [Google Scholar] [CrossRef]
- Valicente, F.H.; Barreto, M.R. Bacillus thuringiensis survey in Brazil: Geographical distribution and insecticidal activity against Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae). Neotrop. Entomol. 2003, 32, 639–644. [Google Scholar]
- Argôlo-Filho, R.C.; Gomes, R.A., Jr.; Barreto, M.R.; Lana, U.G.P.; Valicente, F.H.; Loguercio, L.L. Growth variation among Bacillus thuringiensis strains can affect screening procedures for supernatant-secreted toxins against insect pests. Pest Manag. Sci. 2011, 67, 1184–1192. [Google Scholar]
- Silva-Werneck, J.O.; De-Souza, M.T.; Dias, J.M.C.S.; Ribeiro, B.M. Characterization of Bacillus thuringiensis subsp. kurstaki strain S93 effective against the fall armyworm (Spodoptera frugiperda). Can. J. Microbiol. 1999, 45, 464–471. [Google Scholar] [CrossRef]
- He, J.; Luo, X.; Chen, S.; Cao, L.; Sun, M.; Yu, Z. Determination of spore concentration in Bacillus thuringiensis through the analysis of dipicolinate by capillary zone electrophoresis. J. Chromatogr. A 2003, 994, 207–212. [Google Scholar] [CrossRef]
- Barreto, M.R.; Loguercio, L.L.; Valicente, F.H.; Paiva, E. Insecticidal activity of culture supernatants from Bacillus thuringiensis Berliner strains against Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) larvae. An. Soc. Entomol. Bras. 1999, 28, 675–685. [Google Scholar] [CrossRef]
- Loguercio, L.L.; Barreto, M.R.; Rocha, T.L.; Santos, C.G.; Teixeira, F.F.; Paiva, E. Combined analysis of supernatant-based feeding bioassays and PCR as a first-tier screening strategy for Vip-derived activities in Bacillus thuringiensis strains effective against tropical fall armyworm. J. Appl. Microbiol. 2002, 93, 269–277. [Google Scholar] [CrossRef]
- Bond, R.P.M.; Boyce, C.B.C.; French, S.J. A purification and some properties of an insecticidal exotoxin from Bacillus thuringiensis Berliner. Biochem. J. 1969, 114, 477–488. [Google Scholar]
- Hernández, C.S.; Ferré, J.; Larget-Thiéry, I. Update on the detection of β-exotoxin in Bacillus thuringiensis strains by HPLC analysis. J. Appl. Microbiol. 2001, 90, 643–647. [Google Scholar] [CrossRef]
- Innes, T.C.M.; Bouwer, G. An improved bioassay for the detection of Bacillus thuringiensis β-exotoxin. J. Invertebr. Pathol. 2009, 101, 137–139. [Google Scholar]
- Masson, L.; Erlandson, M.; Puzstai-Carey, M.; Brousseau, R.; Juárez-Pérez, V.; Frutos, R. A holistic approach for determining the entomopathogenic potential of Bacillus thuringiensis strains. Appl. Environ. Microbiol. 1998, 64, 4782–4788. [Google Scholar]
- Rice, W.C. Specific primers for the detection of Vip3A insecticidal gene within a Bacillus thuringiensis collection. Lett. Appl. Microbiol. 1999, 28, 378–382. [Google Scholar] [CrossRef]
- Slininger, P.J.; Behle, R.W.; Jackson, M.A.; Schisler, D.A. Discovery and development of biological agents to control crop pests. Neotrop. Entomol. 2003, 32, 183–195. [Google Scholar] [CrossRef]
- Hoddle, M.S. The strength of biological control in the battle against invasive pests: A reply. Conserv. Biol. 2004, 18, 61–64. [Google Scholar] [CrossRef]
- Louda, S.M.; Stiling, P. The doubled-edged sword of biological control in conservation and restoration. Conserv. Biol. 2004, 18, 50–53. [Google Scholar]
- Höfte, H.; Whiteley, H.R. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 1989, 53, 242–255. [Google Scholar]
- Bravo, A.; Sarabia, S.; Lopez, L.; Ontiveros, H.; Abarca, C.; Ortiz, A.; Ortiz, M.; Lina, L.; Villa-lobos, F.J.; Guadalupe, P.; et al. Characterization of cry genes in mexican Bacillus thuringiensis strain collection. Appl. Environ. Microbiol. 1998, 64, 4965–4972. [Google Scholar]
- Luo, K.; Banks, D.; Adang, M.J. Toxicity, binding, and permeability analyses of four Bacillus thuringiensis Cry1 δ-endotoxins using brush border membrane vesicles of Spodoptera exigua and Spodoptera frugiperda. Appl. Environ. Microbiol. 1999, 65, 457–464. [Google Scholar]
- Valicente, F.H.; Barreto, M.R.; Vasconcelos, M.J.V.; Figueiredo, J.E.F.; Paive, E. PCR identification of Cry I genes in Bacillus thuringiensis strains that are efficient against fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae). An. Soc. Entomol. Bras. 2000, 29, 147–153. [Google Scholar] [CrossRef]
- Cornell Guide for Integrated Field Crop Management 2012. Available online: http://www.ipmguidelines.org/FieldCrops/ (accessed on 10 October 2012).
- Keller, M.; Sneh, B.; Strizhov, N.; Prudovsky, E.; Regev, A.; Koncz, C.; Schell, J.; Zilberstein, A. Digestion of delta-endotoxin by gut proteases may explain reduced sensitivity of advanced instar larvae of Spodoptera littoralis to Cry1C. Insect Biochem. Mol. Biol. 1996, 26, 365–373. [Google Scholar] [CrossRef]
- Kostichka, K.; Warren, G.W.; Mullins, M.; Mullins, A.D.; Palekar, N.V.; Craig, J.A.; Koziel, M.G.; Estruch, J.J. Cloning of a cryV-type insecticidal protein gene from Bacillus thuringiensis: The cryV-encoded protein is expressed early in stationary phase. J. Bacteriol. 1996, 178, 2141–2144. [Google Scholar]
- Tabashnik, B.E. Evolution of resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 1994, 39, 47–79. [Google Scholar] [CrossRef]
- Espinasse, S.; Gohar, M.; Chaufaux, J.; Buisson, C.; Perchat, S.; Sanchis, V. Correspondence of high levels of beta-exotoxin I and the Presence of cry1B in Bacillus thuringiensis. Appl. Environ. Microbiol. 2002, 68, 4182–4186. [Google Scholar] [CrossRef]
- WHO—World Health Organization. Microbial Pest Control Agent: Bacillus thuringiensis; World Health Organization: Geneva, Switzerland, 1999. [Google Scholar]
- Carlberg, G.; Kihamia, C.M.; Minjas, J. Microbial control of flies in latrines in Dares Salaam with a Bacillus thuringiensis (serotype 1) preparation, Muscabac. World J. Appl. Microbiol. Biotechnol. 1985, 1, 33–44. [Google Scholar] [CrossRef]
- Capalbo, D.M.F.; Valicente, F.H.; Moraes, I.O.; Pelizer, L.H. Solid-state fermentation of Bacillus thuringiensis tolworthi to control fall armyworm in maize. Electron. J. Biotechnol. 2001, 4, 112–115. [Google Scholar]
- Nakano, O.; Silveira-Neto, S.; Zucchi, R.A. Entomologia Econômica; Luiz de Queiroz College of Agriculture (ESALQ-USP): Piracicaba, Brazil, 1981. [Google Scholar]
- Argôlo-Filho, R.C.; Loguercio, L.L. Bacillus thuringiensis is an environmental pathogen and host-specificity has developed as an adaptation to human-generated ecological niches. Insects 2014, 5, 62–91. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Argôlo-Filho, R.C.; Costa, R.L.; Pinheiro, D.H.; Corrêa, F.M.; Valicente, F.H.; Pomella, A.W.V.; Loguercio, L.L. Requirement of Simultaneous Assessment of Crystal- and Supernatant-Related Entomotoxic Activities of Bacillus thuringiensis Strains for Biocontrol-Product Development. Toxins 2014, 6, 1598-1614. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins6051598
Argôlo-Filho RC, Costa RL, Pinheiro DH, Corrêa FM, Valicente FH, Pomella AWV, Loguercio LL. Requirement of Simultaneous Assessment of Crystal- and Supernatant-Related Entomotoxic Activities of Bacillus thuringiensis Strains for Biocontrol-Product Development. Toxins. 2014; 6(5):1598-1614. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins6051598
Chicago/Turabian StyleArgôlo-Filho, Ronaldo Costa, Robson Luz Costa, Daniele Heloisa Pinheiro, Fábio Mathias Corrêa, Fernando Hercos Valicente, Alan William Vilela Pomella, and Leandro Lopes Loguercio. 2014. "Requirement of Simultaneous Assessment of Crystal- and Supernatant-Related Entomotoxic Activities of Bacillus thuringiensis Strains for Biocontrol-Product Development" Toxins 6, no. 5: 1598-1614. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins6051598