Diagnosis of Snakebite and the Importance of Immunological Tests in Venom Research

Abstract
:1. Introduction
2. Background




3. Biodetection Methods Considered for Use in Venom Research

4. Major Roles of Enzyme-linked Immunosorbent Assay (ELISA) or Enzyme Immunoassay (EIA) in Venom Research
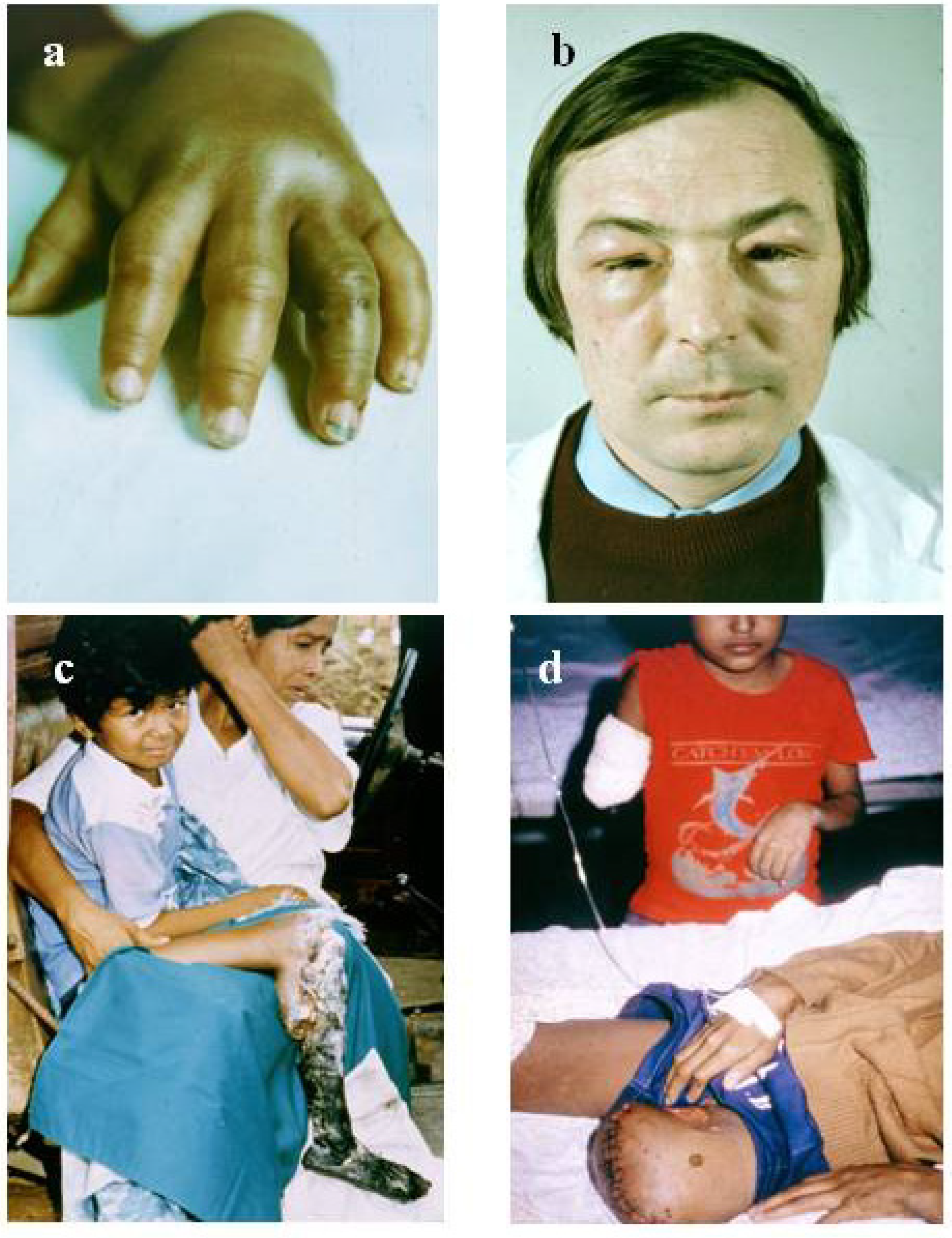
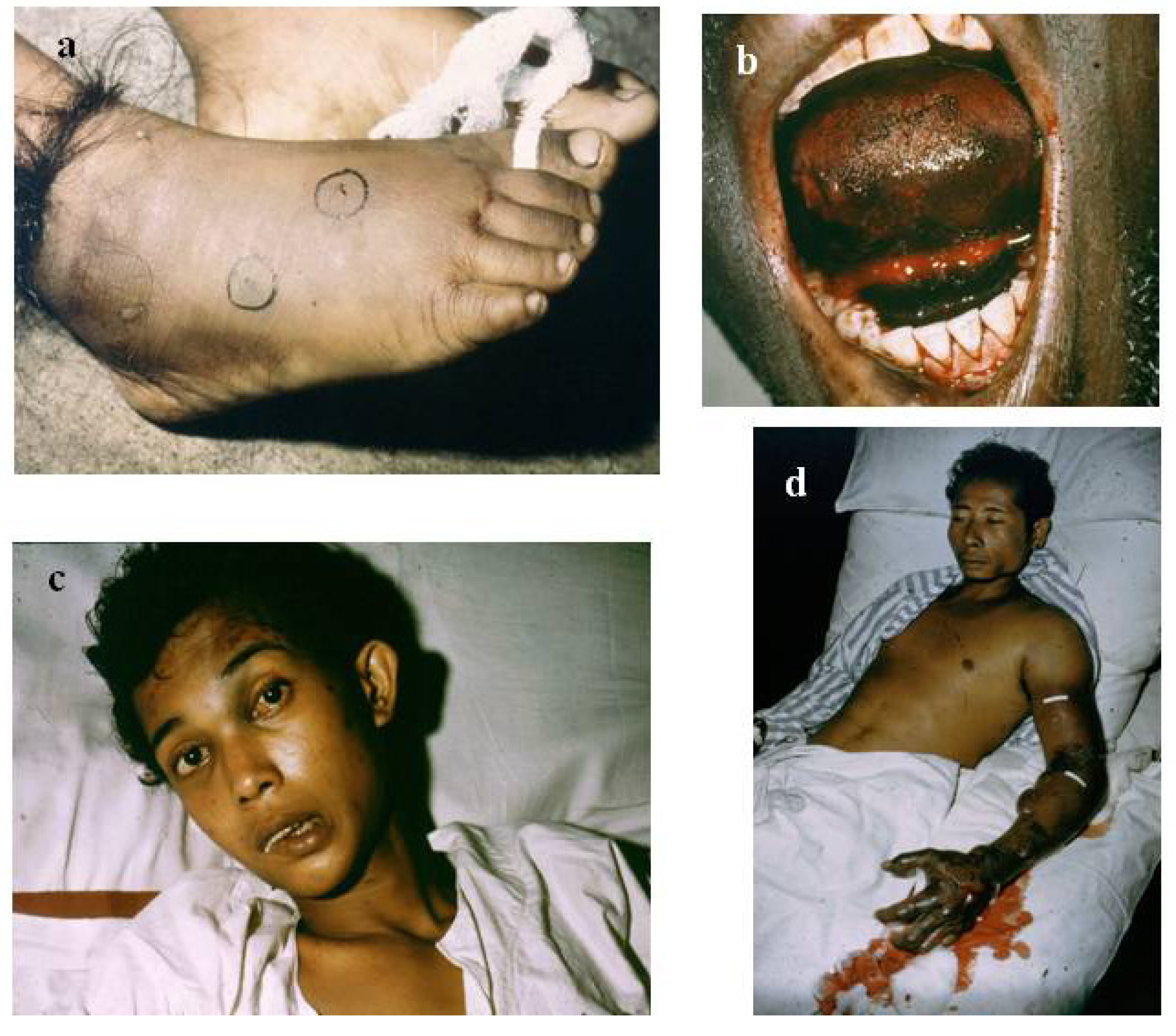
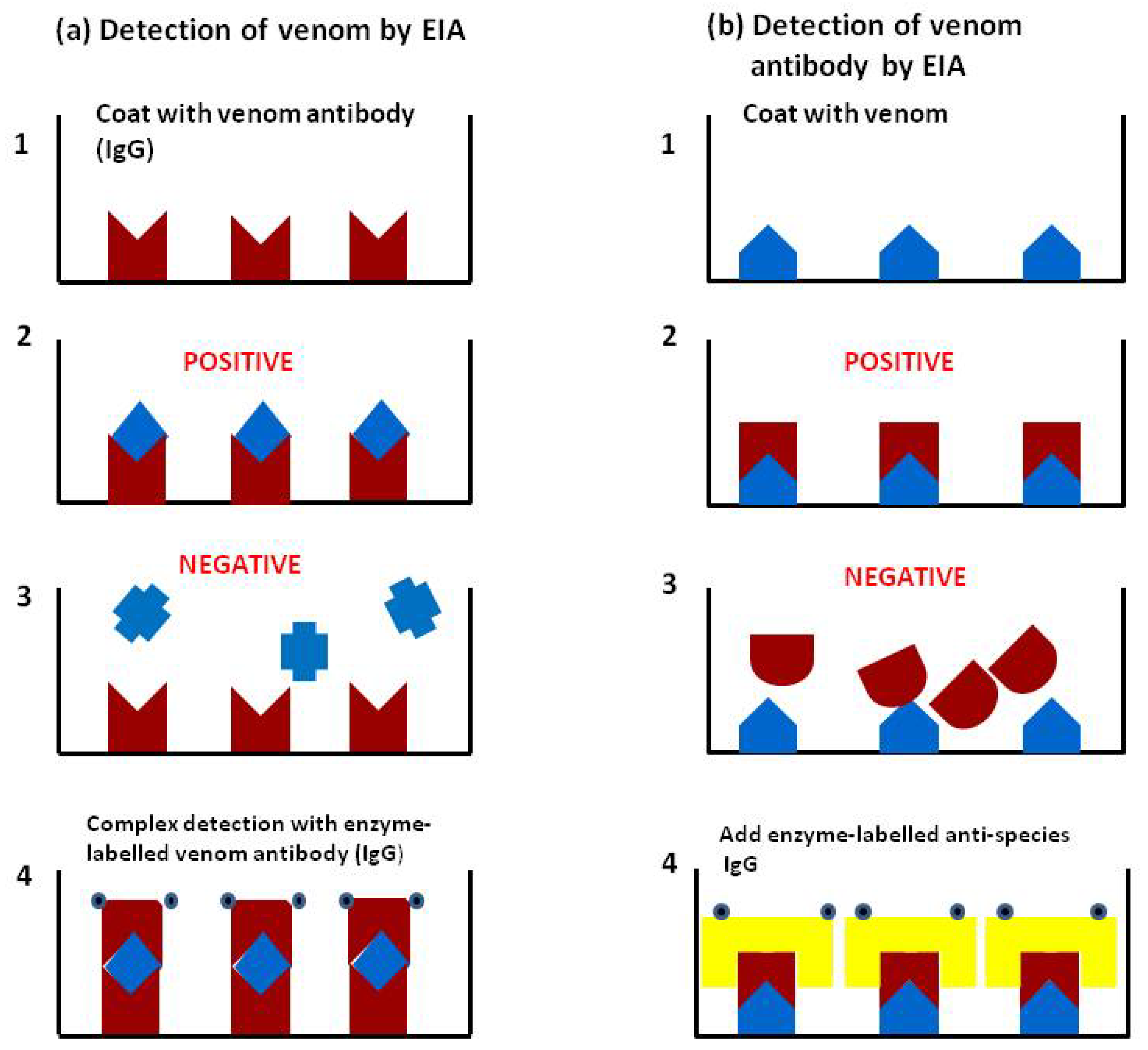
4.1. Accurate Retrospective Diagnosis: Which Snake is Responsible for the Accident?
4.1.1. Validation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Snakes brought | Samples EIA positive | Venom levels (range ng/mL) |
|---|---|---|---|
| Bothrops atrox | 26 | 26 (100%) | 3-466 |
| B.bilineatus | 25 | 24 (96%) | 22-421 |
| Porthidium hyoprora | 1 | 1 | 350 |
| B.microphthalmus | 1 | No assay developed | 5 * |
| B.taeniatus | 4 | 3 (75%) | 23-225 |
| Non venomous | 2 | 0 | - |
| Total venomous | 57 | 54 (95%) |
4.1.2. Application

4.1.3. Rapid EIA
4.2. The Assessment of New and Existing Antivenoms; How Good is the Antivenom?
4.2.1. Preclinical Assessment of Antivenom Using in Vivo and in Vitro Tests
4.2.2. Clinical Assessment (Resolution of Clinical Signs in Patients)
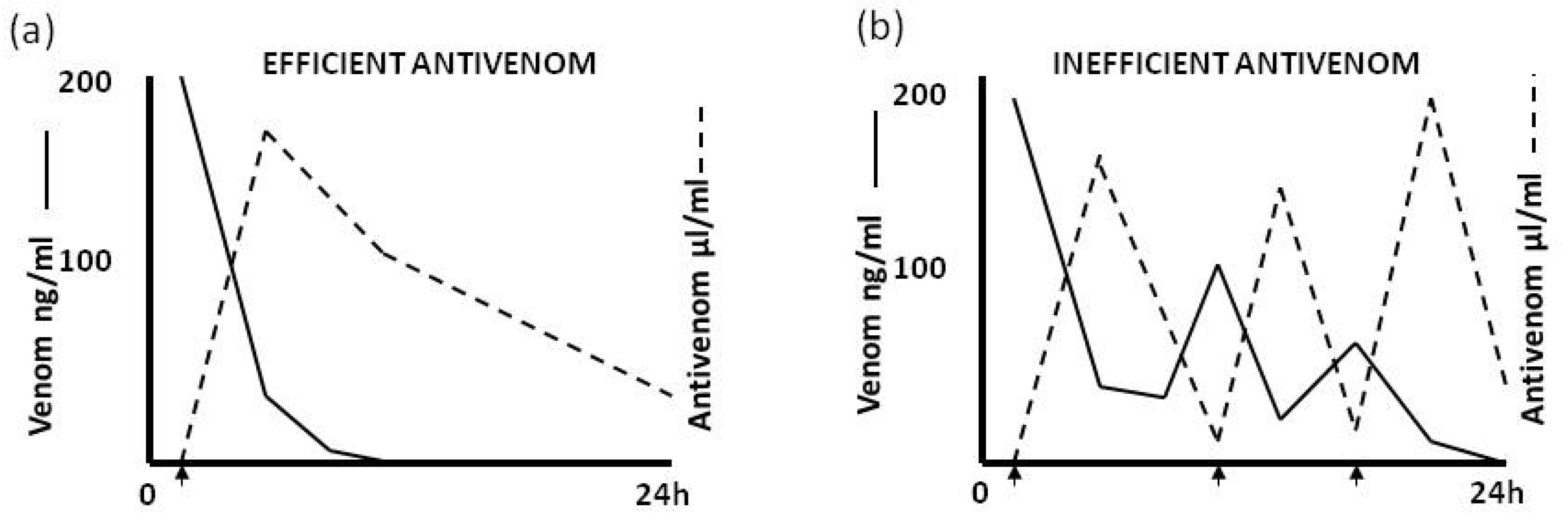
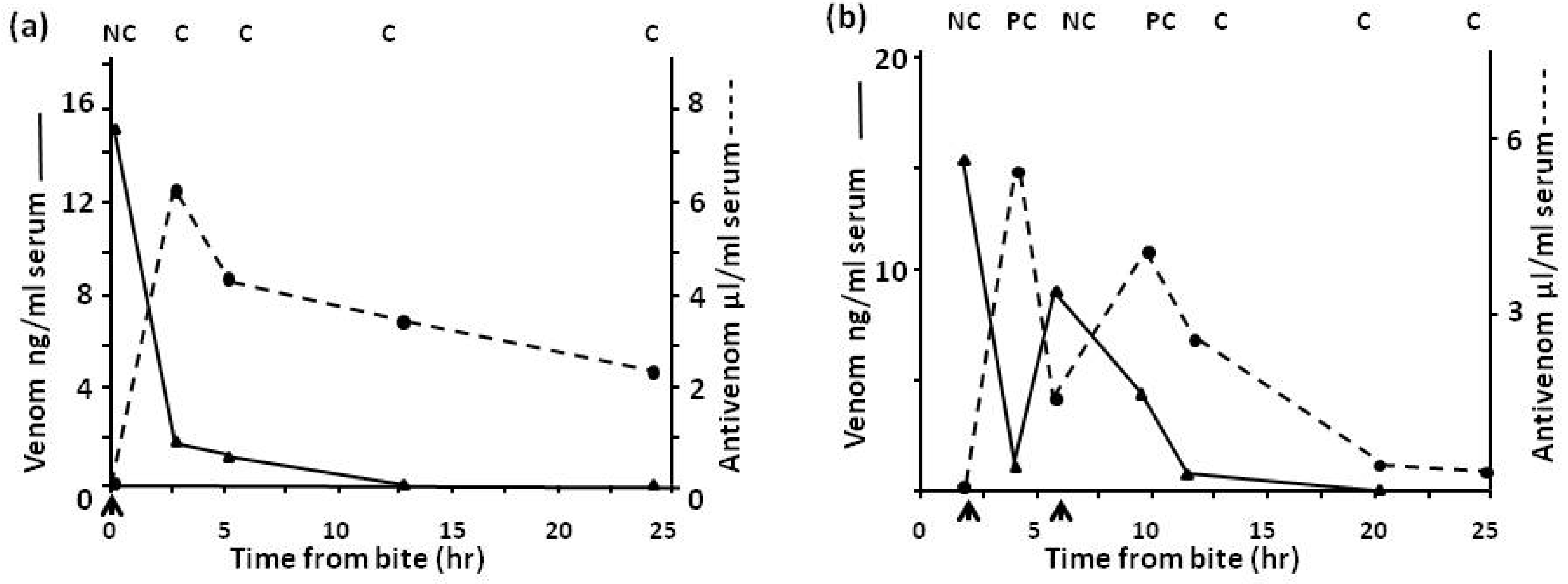
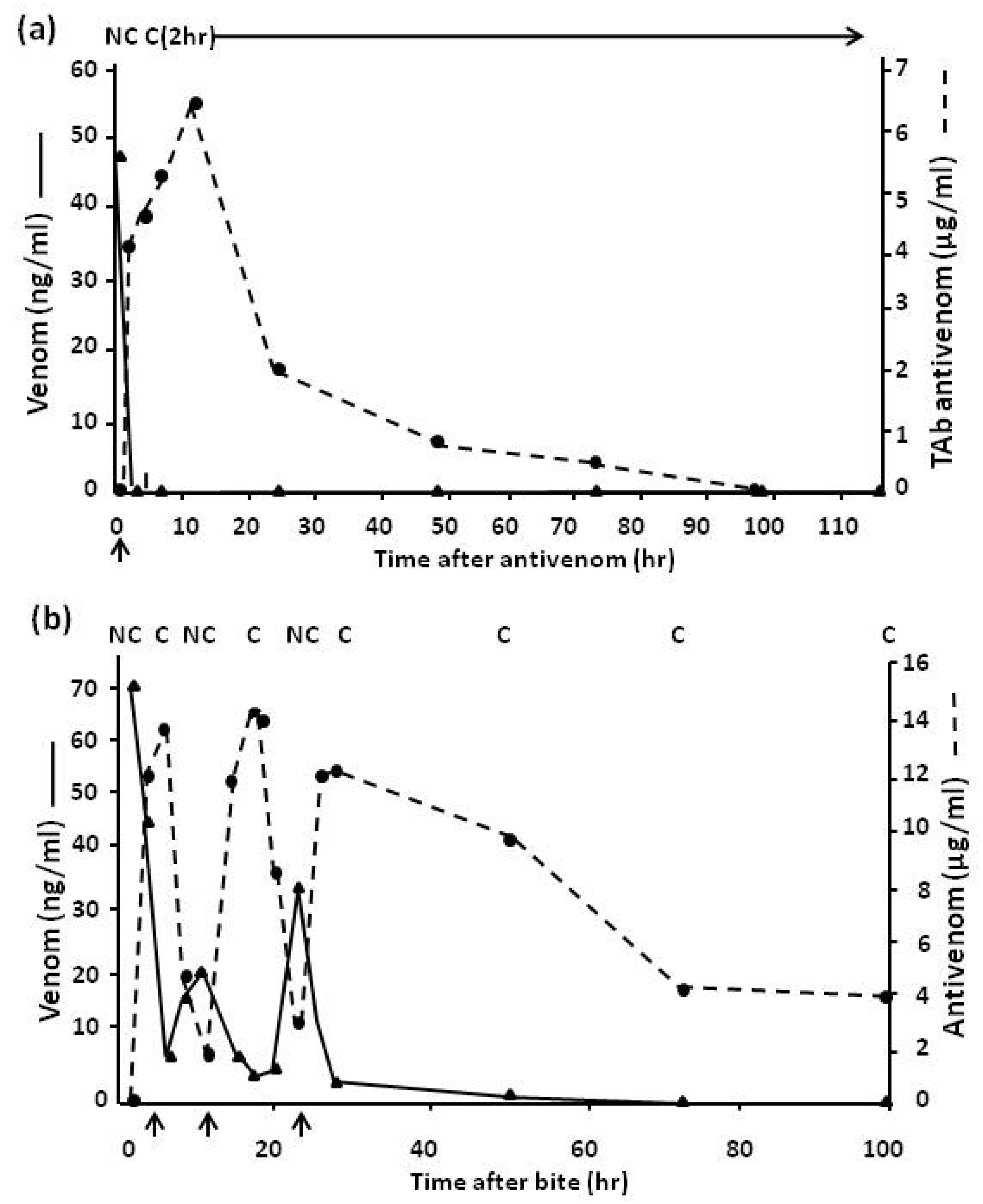
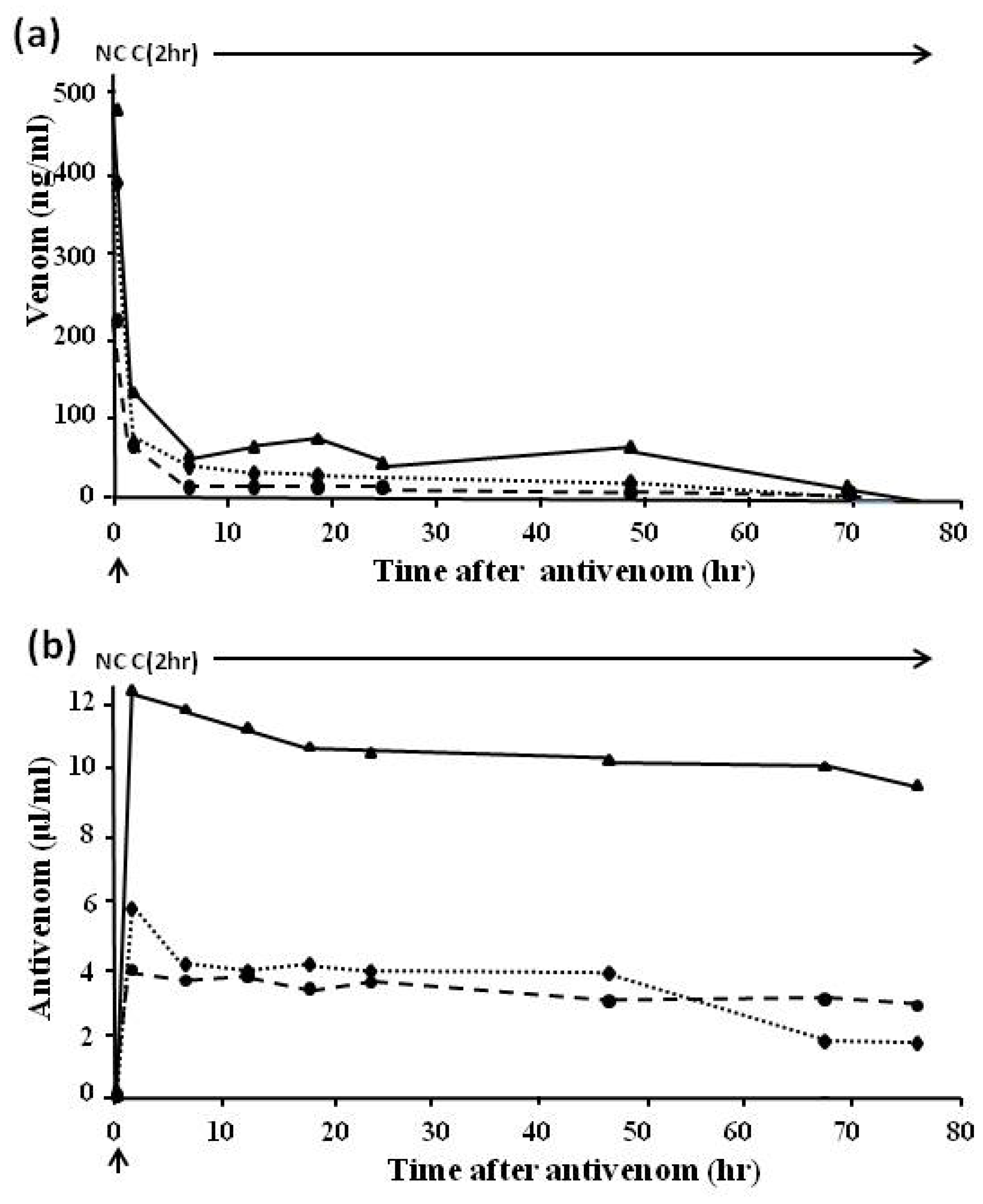
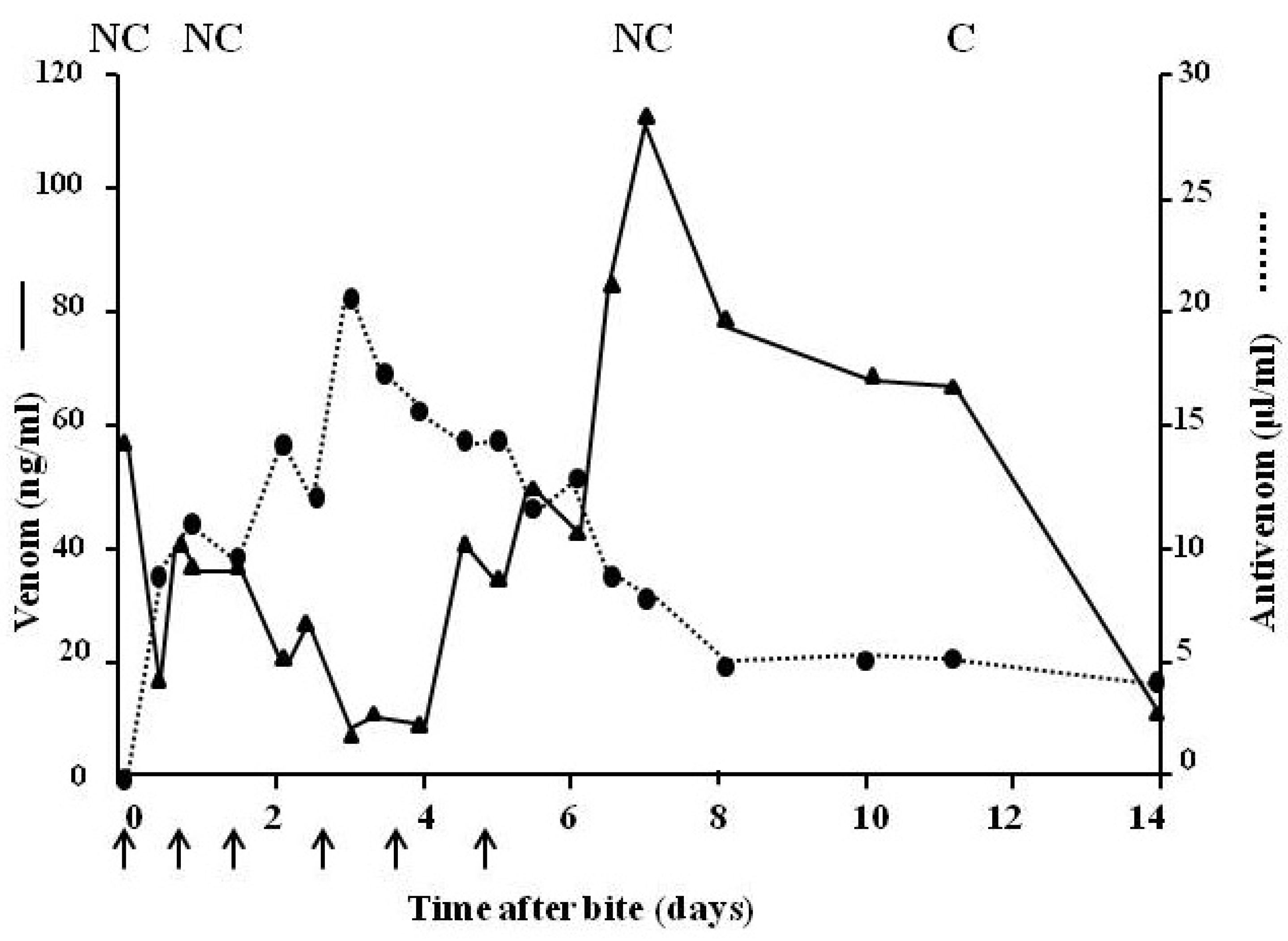
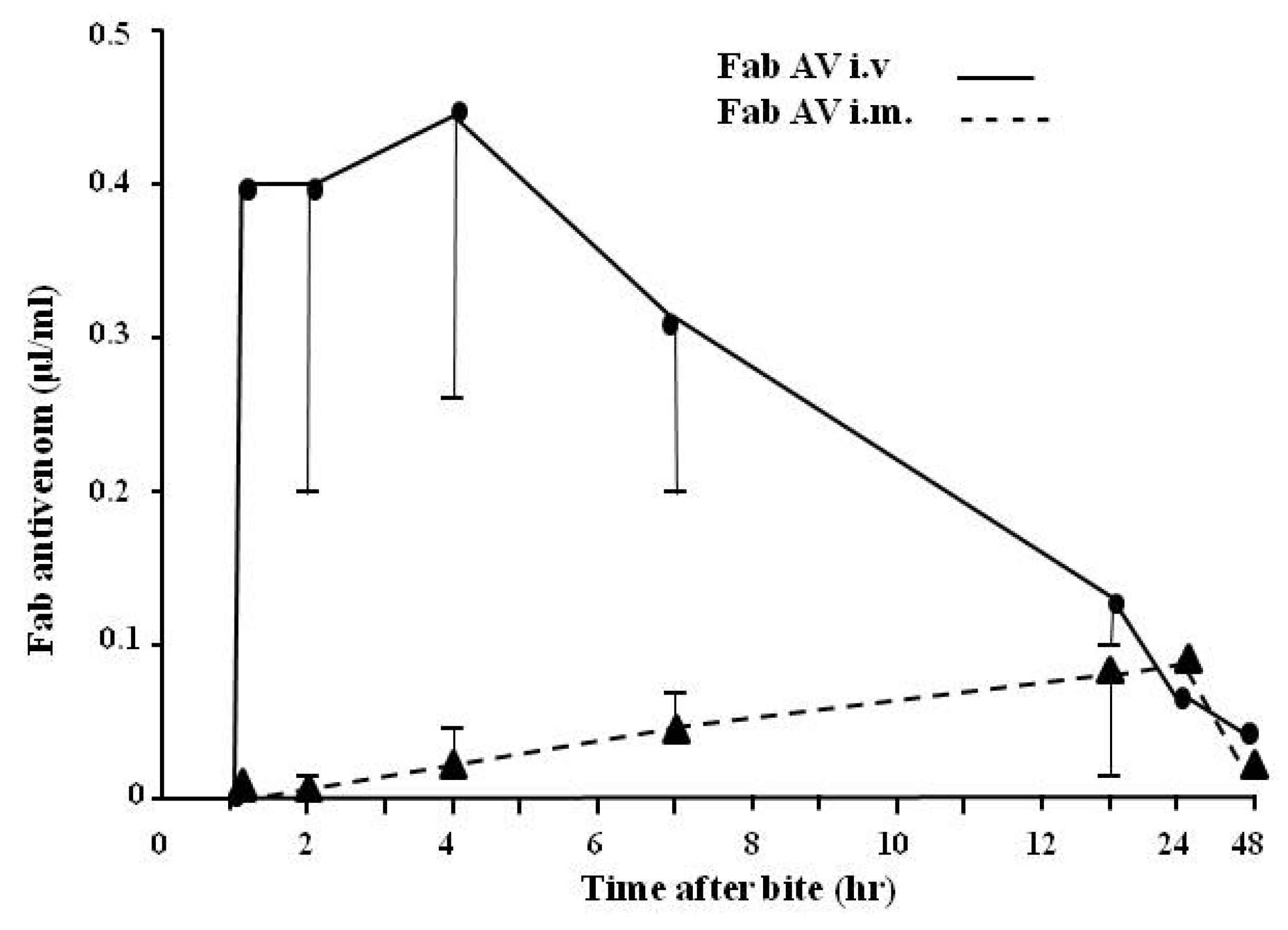
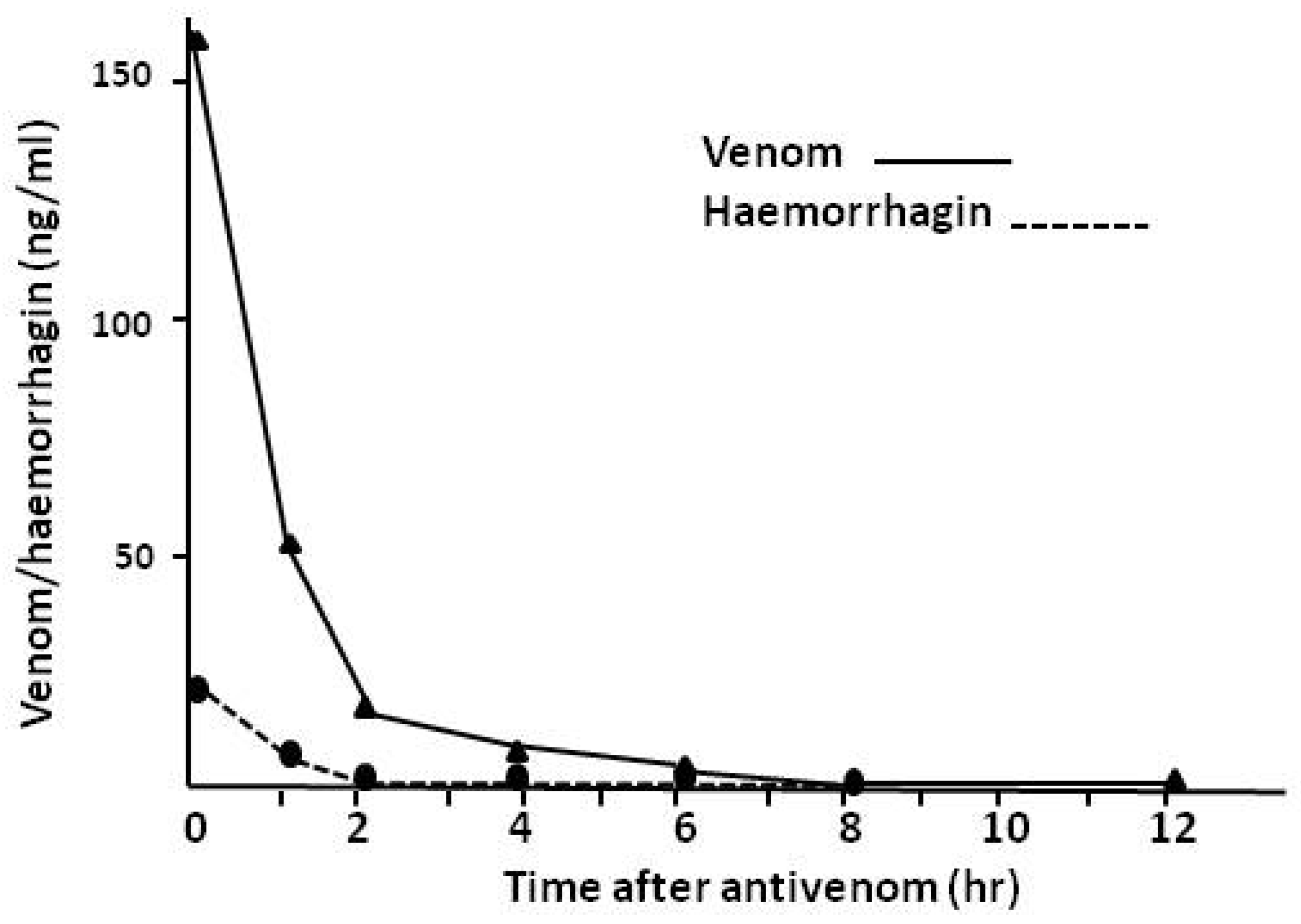
4.2.3. Pharmacokinetic Studies (Rate of Venom Elimination from the Circulation)






4.3. Assessment of First Aid Measures
4.3.1. Possible early First Aid Treatment with Antivenom in the Field

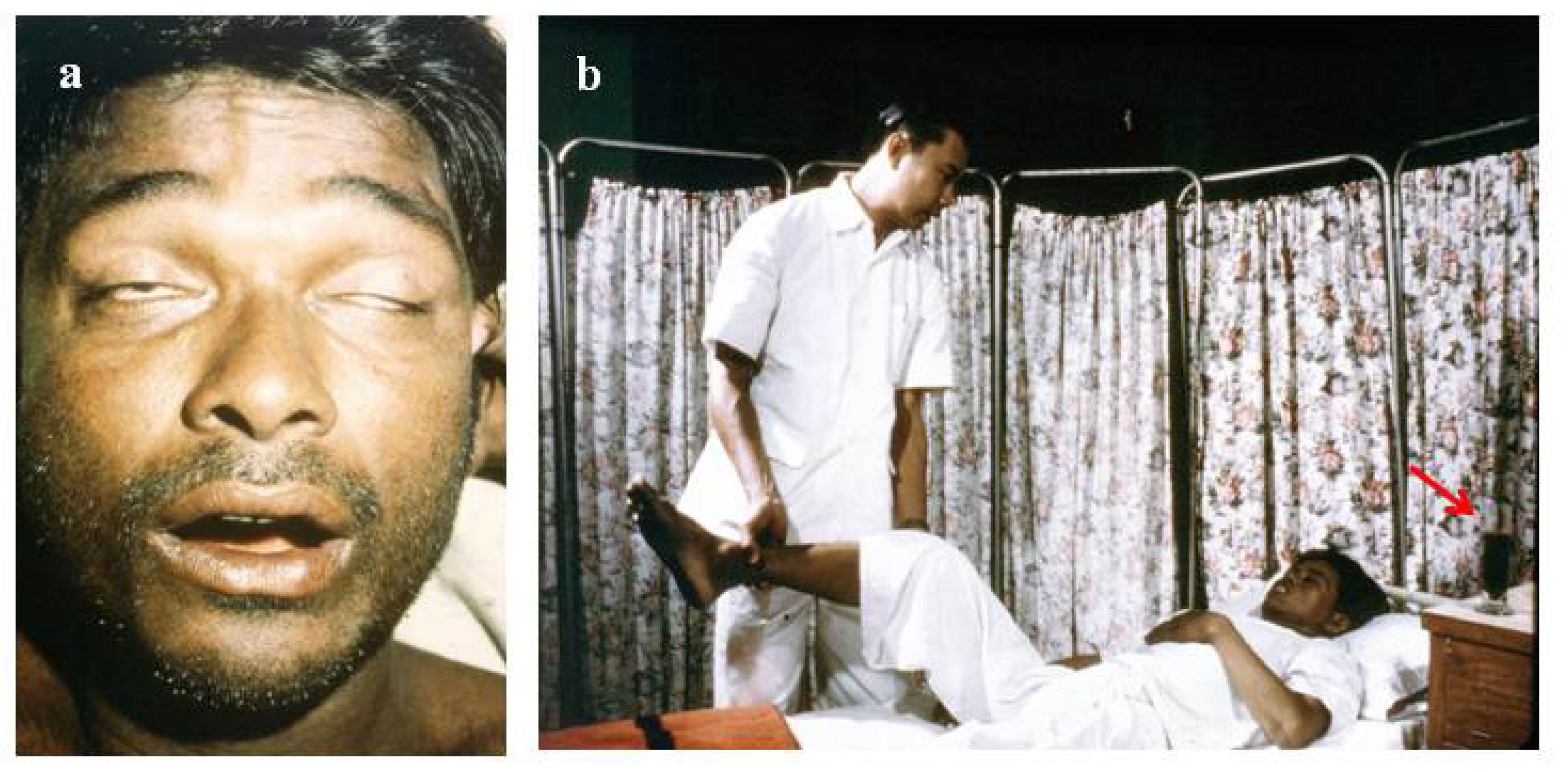
4.3.2. The Use of Tourniquets

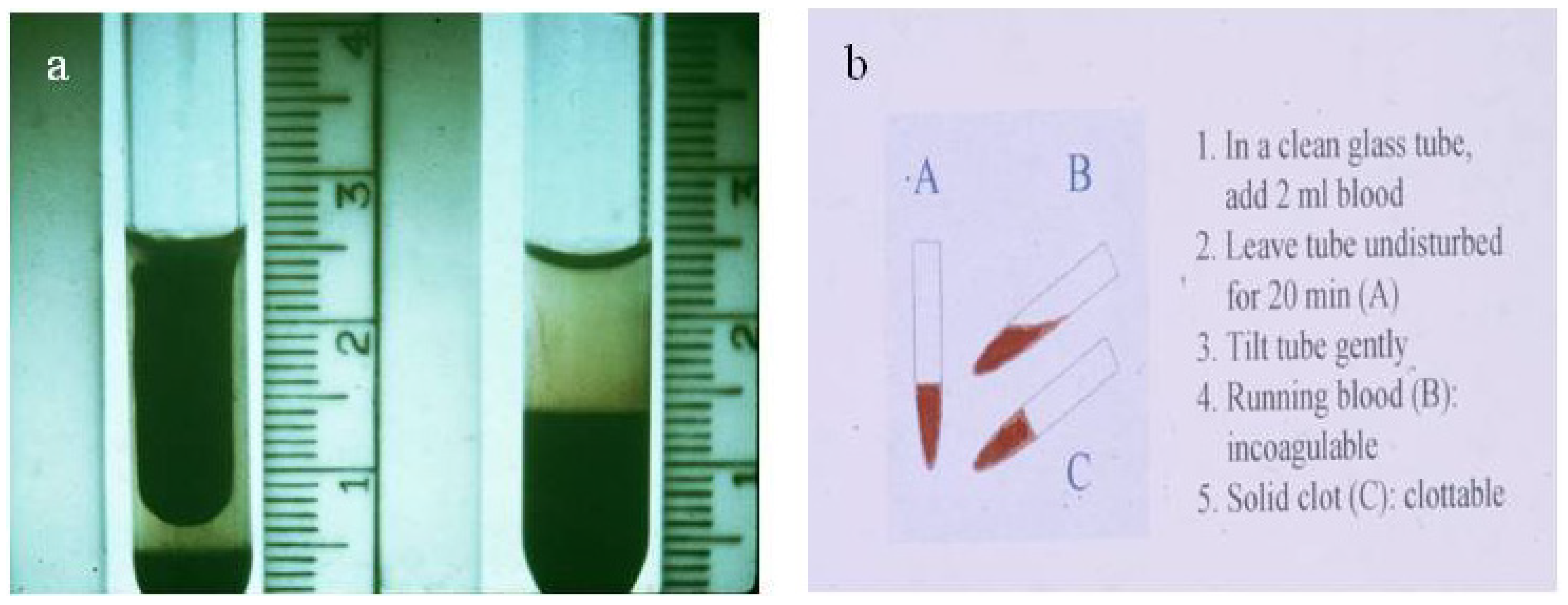
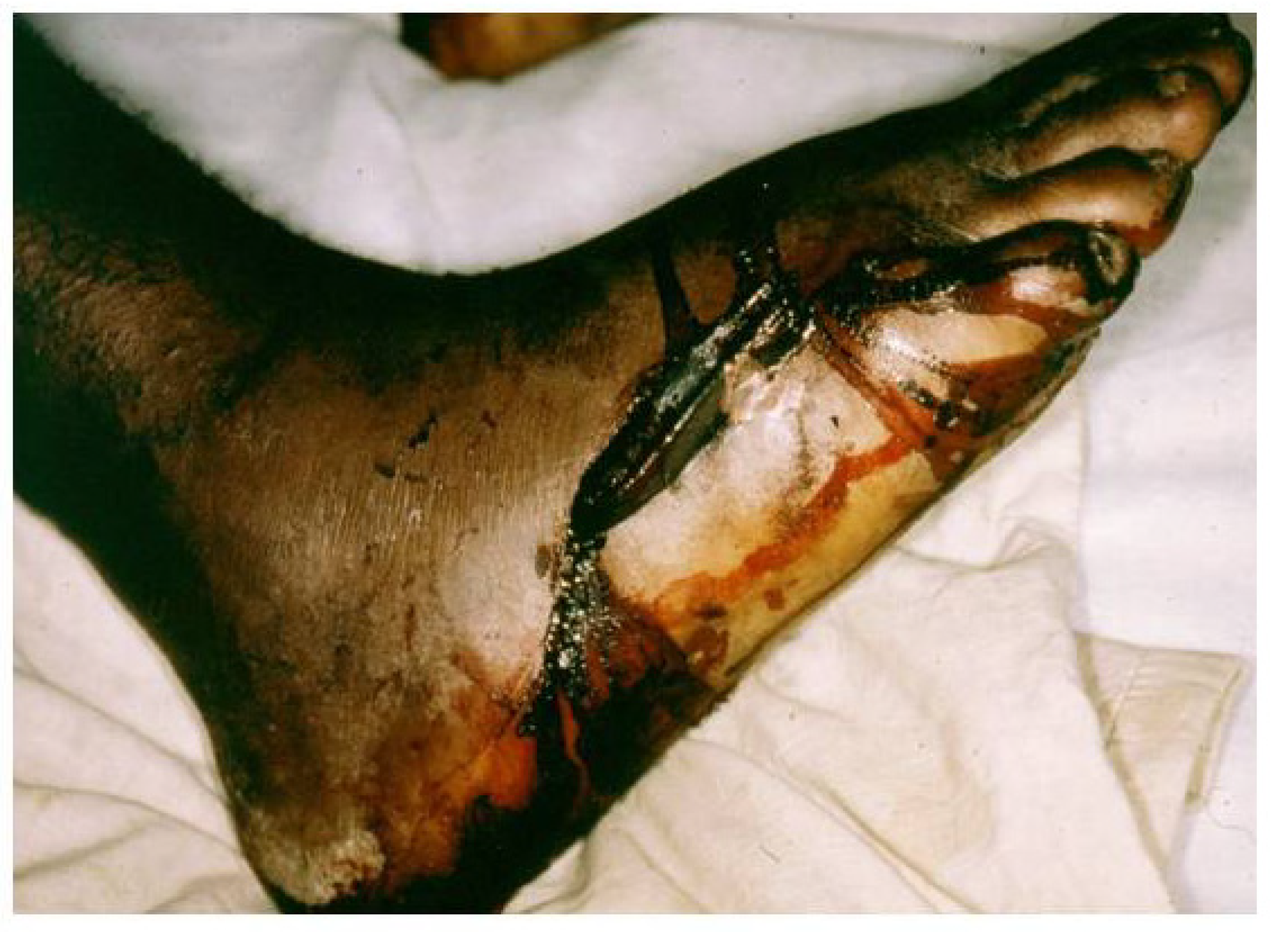
4.3.3. The Use of Local Remedies in Treatment of Snake Bite

| Sample | Venom antigen concentration(mean ± SD (n = 5)) |
|---|---|
| Serum | 11.6 ± 5.6 ng/mL |
| Bite site | 21.6 ± 6.7 ng/mL |
| Stone | 0.3 ± 0.4 ng/mL |
4.4. Epidemiological Studies

4.5. Detection of Individual Venom Components

5. Conclusions
Acknowledgments
Author Contributions
Conflict of Interest
References
- Theakston, R.D.G.; Lloyd-Jones, M.J.; Reid, H.A. Micro-ELISA for detecting and assaying snake venom and venom-antibody. Lancet 1977, 2, 639–641. [Google Scholar] [CrossRef]
- Sutherland, S.K.; Coulter, A.R.; Broad, A.J. Human snakebite victims: The successful detection of circulating snake venom by radioimmunoassay. Med. J. Aust. 1975, 1, 27–29. [Google Scholar]
- Sutherland, S.K.; Coulter, A.R. Snake bite: Detection of venom by radioimmunoassay. Med. J. Aust. 1977, 2, 683–684. [Google Scholar]
- Coulter, A.R.; Harris, R.D.; Sutherland, S.K. Enzyme immunoassay for the rapid clinical identification of snake venom. Med. J. Aust. 1980, 1, 433–435. [Google Scholar]
- O’Leary, M.A.; Isbister, G.K.; Schneider, J.J.; Brown, S.G.A.; Currie, B.J. Enzyme immunoassaysin brown snake (Pseudonaja spp) envenoming: Detecting venom, antivenom and venom-antivenom complexes. Toxicon 2006, 48, 4–11. [Google Scholar] [CrossRef]
- Theakston, R.D.G. The application of imunoassay techniques, including enzyme-linked immunosorbent assay (ELISA), to snake venom research. Toxicon 1983, 21, 341–352. [Google Scholar] [CrossRef]
- Warrell, D.A. Redi award lecture: Clinical studies of snake-bite in four tropical continents. Toxicon 2013, 69, 3–13. [Google Scholar] [CrossRef]
- Warrell, D.A.; Davidson, NMcD.; Greenwood, B.M.; Ormerod, L.D.; Pope, H.M.; Watkins, B.J.; Prentice, C.R.M. Poisoning by bites of the saw-scaled or carpet viper (Echis carinatus) in Nigeria. Q. J. Med. 1977, 46, 33–62. [Google Scholar]
- Warrell, D.A.; Looareesuwan, S.; Theakston, R.D.G.; Phillips, R.E.; Chanthavanich, P.; Virivan, C.; Supanaranond, W.; Karbwang, J.; Ho, M.; Hutton, R.A.; et al. Randomized comparative trial of three monospecific antivenoms for bites by the Malayan pit viper (Calloselasma rhodostoma) in southern Thailand; clinical and laboratory correlations. Am. J. Trop. Med. Hyg. 1986, 35, 1235–1247. [Google Scholar]
- Sano-Martins, I.S.; Fan, H.W.; Castro, S.C.B.; Tomy, S.C.; França, F.O.S.; Jorge, M.T.; Kamiguti, A.S.; Warrell, D.A.; Theakston, R.D. Reliability of the simple 20 minute whole blood clotting test (WBCT20) as an indicator of low plasma fibrinogen concentration in patients envenomed by Bothrops snakes. Toxicon 1994, 32, 1045–1050. [Google Scholar] [CrossRef]
- Muelling, R.J.; Samson, R.F.; Beven, T. The precipitin test in elucidating the cause of death. Am. J. Clin. Path. 1957, 28, 489–494. [Google Scholar]
- Greenwood, B.M.; Warrell, D.A.; Davidson, N.McD.; Ormerod, L.D.; Reid, H.A. Immunodiagnosis of snake bite. Br. Med. J. 1974, 4, 743–745. [Google Scholar] [CrossRef]
- Tiru-Chelvam, R. Demonstration of sites of snake venom localisation by immunofluorescence techniques. J. Pathol. 1972, 107, 303–305. [Google Scholar]
- Boche, R.D.; Russell, F.E. Passive haemagglutination studies with snake venom and antivenin. Toxicon 1968, 6, 125–130. [Google Scholar] [CrossRef]
- Gao, R.; Zhang, Y.; Gopalakrishnakone, P. Single-bead-based immunofluorescence assay for snake venom detection. Biotechnol. Prog. 2008, 24, 245–249. [Google Scholar] [CrossRef]
- Tu, A.T.; Salafranca, E.S. Immunological properties and neutralization of sea snake venoms (II). Am. J. Trop. Med. Hyg. 1974, 23, 135–138. [Google Scholar]
- Gawade, S.P.; Gaitonde, B.B. Immunological studies on monovalent Enhydrina schistosa antivenin. Indian J. Med. Res. 1980, 72, 895–900. [Google Scholar]
- Coulter, A.R.; Sutherland, S.K.; Broad, A.J. Assay of snake venoms in tissue fluids. J. Immunol. Meth. 1974, 4, 297–300. [Google Scholar] [CrossRef]
- Coulter, A.R.; Fox, J.C.; Sutherland, S.K.; Waddell, C.J. A new solid-phase sandwich radioimmunoassay and its application to the detection of snake venom. J. Immunol. Meth. 1978, 23, 241–252. [Google Scholar] [CrossRef]
- Voller, A.; Bartlett, A.; Bidwell, D.E. Enzyme immunoassay for parasitic diseases. Trans. R. Soc. Trop. Med. Hyg. 1976, 70, 98–106. [Google Scholar] [CrossRef]
- Selvanayagam, Z.E.; Gnanavendhan, S.G.; Ganesh, K.A.; Rajagopal, D.; Rao, P.V. ELISA for the detection of venoms from four medically important snakes of India. Toxicon 1999, 37, 757–770. [Google Scholar] [CrossRef]
- Theakston, R.D.G.; Pugh, R.N.H.; Reid, H.A. Enzyme-linked immunosorbent assay of venom antibodies in human victims of snake bite. J. Trop. Med. Hyg. 1981, 84, 109–112. [Google Scholar]
- Theakston, R.D.G. Fieldwork in the tropics: duplicate frozen samples. Lancet 1997, 350, 68. [Google Scholar] [CrossRef]
- Ho, M.; Warrell, M.J.; Warrell, D.A.; Bidwell, D.; Voller, A. A critical reappraisal of the use of enzyme-linked immunosorbent assay in the study of snake bite. Toxicon 1986, 24, 211–221. [Google Scholar] [CrossRef]
- Kulawickrama, S.; O’Leary, M.A.; Hodgson, W.C.; Brown, S.G.A.; Jacoby, T.; Davern, K.; Isbister, G.K. Development of a sensitive enzyme immunoassay for measuring taipan venom in serum. Toxicon 2010, 55, 1510–1518. [Google Scholar] [CrossRef]
- Dong, L.V.; Eng, K.H.; Quyen, L.K.; Gopalakrishnakone, P. Optical immunoassay for snake venom detection. Biosens. Bioelectron. 2004, 19, 1285–1294. [Google Scholar] [CrossRef]
- Suntrarachun, S.; Pakmanee, N.; Tirawatnapong, T.; Chanhome, L.; Sitprija, V. Development of a polymerase chain reaction to distinguish monocellate (Naja kaouthia) bites from other common Thai snake species, using both venom extracts and bite-site swabs. Toxicon 2001, 39, 1087–1090. [Google Scholar] [CrossRef]
- Pook, C.E.; McEwing, R. Mitochondrial DNA sequences from dried snake venom: A DNA barcoding approach to the identification of venom samples. Toxicon 2005, 46, 711–714. [Google Scholar] [CrossRef]
- Fox, J.W.; University of Virginia, Charlottsville, VA, USA. Personal communication, 2013.
- Larrick, J.W.; Yost, J.A.; Kaplan, J. Snake bite among the Waorani Indians of eastern Ecuador. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 542–543. [Google Scholar] [CrossRef]
- Larrick, J.W.; Yost, J.A.; Kaplan, J. Patterns of health and disease among the Waorani Indians of eastern Ecuador. Med. Anthropol. 1979, 3, 147–189. [Google Scholar] [CrossRef]
- Pugh, R.N.H.; Theakston, R.D.G. A clinical study of viper bite poisoning. Ann. Trop. Med. Parasitol. 1987, 81, 135–149. [Google Scholar]
- Phillips, R.E.; Theakston, R.D.G.; Warrell, D.A.; Galagedera, Y.; Abeysekera, D.T.D.J.; Dissanayake, P.; Hutton, R.A.; Aloysius, D.J. Paralysis, rhabdomyolysis and haemolysis caused by bites of Russell’s viper (Vipera russelli pulchella) in Sri Lanka: Failure of Haffkine antivenom. Q. J. Med. 1988, 68, 691–716. [Google Scholar]
- Virivan, C.; Veeravat, U.; Warrell, M.J.; Theakston, R.D.G.; Warrell, D.A. ELISA confirmation of acute and past envenoming by the monocellate Thai cobra (Naja kaouthia). Am. J. Trop. Med. Hyg. 1986, 35, 173–181. [Google Scholar]
- Theakston, R.D.G. Snake venoms in science and clinical medicine. 2. Applied immunology in snake venom research. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 741–744. [Google Scholar] [CrossRef]
- Faiz, A.; Sir Salimullah Medical College, Dhaka, Bangladesh. Personal communication, 2014.
- Harris, J.B.; University of Newcastle, Newcastle upon Tyne, UK. Personal communication, 2014.
- Meyer, W.P.; Habib, H.G.; Onayade, A.A.; Yakubu, A.; Smith, D.C.; Nasidi, A.; Daudu, I.J.; Warrell, D.A.; Theakston, R.D.G. First clinical experiences with a new ovine Fab Echis ocellatus snakebite antivenom in Nigeria: randomised comparative trial with Institute Pasteur serum (Ipser) Africa antivenom. Am. J. Trop. Med. Hyg. 1997, 56, 291–300. [Google Scholar]
- Trevett, A.J.; Lalloo, D.G.; Nwokolo, N.C.; Theakston, R.D.G.; Naraqi, S.; Warrell, D.A. An analysis of the sensitivity and cost effectiveness of venom detection kits in the management of snake bite victims in Papua New Guinea. Toxicon 1995, 33, 703–705. [Google Scholar] [CrossRef]
- Steuten, J.; Winkel, K.; Carroll, T.; Williamson, N.A.; Ignjatovic, V.; Fung, K.; Purcell, A.W.; Fry, B.G. The molecular basis of cross reactivity in the Australian Snake Venom Detection Kit (SVDK). Toxicon 2007, 50, 1041–1052. [Google Scholar] [CrossRef]
- Jelinek, G.A.; Tweed, C.; Lynch, D.; Celenza, T.; Bush, B.; Michalopoulos, N. Cross reactivity between venomous, mildly venomous, and non-venomous snake venoms with the Commonwealth Serum Laboratories Venom Detection Kit. Emerg. Med. Australas. 2004, 16, 459–464. [Google Scholar] [CrossRef]
- Isbister, G.K.; Brown, S.G.; Page, C.B.; McCoubrie, D.L.; Greene, S.L.; Buckley, N.A. Snakebite in Australia: A practical approach to diagnosis and treatment. Med. J. Aust. 2013, 199, 763–768. [Google Scholar] [CrossRef]
- World Health Organization. Progress in the characterization of venoms and standardization of antivenoms; WHO Offset Publication No.58; World Health Organization: Geneva, Switzerland, 1981. [Google Scholar]
- World Health Organization. 59th Meeting of the WHO Expert Committee on Biological Standardization, Geneva, Switzerland, 13–17 October 2008.
- Theakston, R.D.G.; Warrell, D.A.; Griffiths, E. Report of a WHO workshop on the standardization and control of antivenoms. Toxicon 2003, 41, 541–557. [Google Scholar] [CrossRef]
- Laing, G.D.; Theakston, R.D.G.; Leite, R.P.; Dias da Silva, W.D.; Warrell, D.A.; B.I.A.S.G. Comparison of the potency of three Brazilian Bothrops antivenoms using in vivo rodent and in vitro assays. Toxicon 1992, 30, 1219–1225. [Google Scholar] [CrossRef]
- Laing, G.D.; Lee, L.; Smith, D.C.; Landon, J.; Theakston, R.D.G. Experimental assessment of a new, low-cost antivenom for treatment of carpet viper (Echis ocellatus) envenoming. Toxicon 1995, 33, 307–313. [Google Scholar] [CrossRef]
- Laing, G.D.; Renjifo, J.M.; Ruiz, F.; Harrison, R.A.; Nasidi, A.; Gutierrez, J-M.; Warrell, D.A.; Theakston, R.D.G. A new Pan African polyspecific antivenom developed in response to the antivenom crisis in Africa. Toxicon 2003, 42, 35–41. [Google Scholar] [CrossRef]
- Laing, G.D.; Yarleque, A.; Marcelo, A.; Rodriguez, E.; Warrell, D.A.; Theakston, R.D.G. Preclinical testing of three South American antivenoms against the venoms of five medically- important Peruvian snake venoms. Toxicon 2004, 44, 103–106. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Rojas, E.; Queseda, L.; Leon, G.; Nunez, J.; Laing, G.D.; Sasa, M.; Renjifo, J.M.; Nasidi, A.; Warrell, D.A.; et al. Pan-African polyspecific antivenom produced by caprylic acid purification of horse IgG: An alternative to the antivenom crisis in Africa. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 468–475. [Google Scholar] [CrossRef]
- Segura, A.; Villalta, M.; Herrera, M.; Leon, G.; Harrison, R.; Durfa, N.; Nasidi, A.; Calvete, J.J.; Theakston, R.D.G.; Warrell, D.A.; et al. Preclinical assessment of the efficacy of a new antivenom (EchiTab-plus-ICP®) for the treatment of viperid snakebite envenoming in sub-Saharan Africa. Toxicon 2010, 55, 369–374. [Google Scholar]
- Theakston, R.D.G.; Laing, G.D.; Fielding, C.M.; Freire Lascano, A.; Touzet, J-M.; Vallejo, F.; Guderian, R.H.; Nelson, S.J.; Wuster, W.; Richards, A.M.; et al. Treatment of snake bites by Bothrops species and Lachesis muta in Ecuador: Laboratory screening of candidate antivenoms. Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 550–554. [Google Scholar] [CrossRef]
- Theakston, R.D.G.; Reid, H.A. Development of simple standard assay procedures for the characterization of snake venoms. Bull. WHO 1983, 61, 949–956. [Google Scholar]
- Sells, P.G. Animal experimentation in snake venom research and in vitro alternatives. Toxicon 2003, 42, 115–133. [Google Scholar] [CrossRef]
- Theakston, R.D.G.; Reid, H.A. Enzyme-linked immunosorbent assay (ELISA) in assessing antivenom potency. Toxicon 1979, 17, 511–515. [Google Scholar] [CrossRef]
- Abubakar, I.S.; Abubakar, S.B.; Habib, A.G.; Nasidi, A.; Durfa, N.; Yusuf, P.O.; Larnyang, S.; Garnvwa, J.; Sokomba, E.; Salako, L.; et al. Randomised controlled double-blind non-inferiority trial of two antivenoms for saw-scaled or carpet viper (Echis ocellatus) envenoming in Nigeria. PLoS Negl. Trop. Dis. 2010, 4, 1–10. [Google Scholar]
- Smalligan, R.; Cole, J.; Brito, N.; Laing, G.D.; Mertz, B.L.; Manock, S.; Maudlin, J.; Quist, B.; Holland, G.; Nelson, S.; et al. Crotaline snake bite in the Ecuadorian Amazon: Randomised double blind comparative trial of three South American polyspecific antivenoms. Br. Med. J. 2004, 329, 1129–1133. [Google Scholar] [CrossRef]
- Warrell, D.A.; Hudson, B.J.; Lalloo, D.G.; Trevett, A.J.; Whitehead, P.; Bamler, P.R.; Ranaivoson, M.; Wiyono, A.; Richie, T.L.; Fryauff, D.J.; et al. The emerging syndrome of envenoming by the New Guinea small-eyed snake Micropechis ikaheka. Q. J. Med. 1996, 89, 523–530. [Google Scholar] [CrossRef]
- Lalloo, D.G.; Trevett, A.J.; Black, J.; Mapao, J.; Saweri, A.; Naraqi, S.; Owens, D.; Kamiguti, A.S.; Hutton, R.A.; Theakston, R.D.G.; et al. Neurotoxicity, anticoagulant activity and evidence of rhabdomyolysis in patients bitten by death adders (Acanthophis sp.) in southern Papua New Guinea. Q. J. Med. 1996, 89, 25–35. [Google Scholar] [CrossRef]
- Riviere, G.; Choumet, V.; Audebert, F.; Sabouraud, A.; Debray, M.; Scherrmann, J.M.; Bon, C. Effect of antivenom on venom pharmacokinetics in experimentally envenomed rabbits: Toward an optimization of antivenom therapy. J. Pharmacol. Exp. Ther. 1997, 281, 1–8. [Google Scholar]
- Maung-Maung-Thwin; Khin-Mee-Mee; Mi-Mi-Kyin; Thein-Than. Kinetics of envenomation with Russell’s viper (Vipera russelli) venom and of antivenom use in mice. Toxicon 1988, 26, 373–378. [Google Scholar] [CrossRef]
- Mello, S.M.; Linardi, A.; Rennó, A.L.; Tarsitano, C.A.B.; Pereira, E.M.; Hyslop, S. Renal kinetics of Bothrops alternatus (Urutu) snake venom in rats. Toxicon 2010, 55, 470–480. [Google Scholar] [CrossRef]
- Ismail, M.; Aly, M.H.M.; Abd-Elsalami, M.A.; Morad, A.M. A three-compartment open pharmacokinetic model can explain variable toxicities of cobra venoms and their alpha toxins. Toxicon 1996, 34, 1011–1026. [Google Scholar]
- O’Leary, M.A.; Isbister, G.K. Detection of venom-antivenom (VAV) immunocomplexes in vitro as a measure of antivenom efficacy. Toxicon 2014, 77, 125–132. [Google Scholar] [CrossRef]
- Rojas, A.; Vargas, M.; Ramírez, N.; Estrada, R.; Segura, Á.; Herrera, M.; Villata, M.; Gómez, A.I.; Gutiérrez, J.M.; León, G. Role of the animal model on the pharmacokinetics of equine-derived antivenoms. Toxicon 2014, 70, 9–14. [Google Scholar]
- Cardoso, J.L.C.; Fan, H.W.; França, F.O.S.; Jorge, M.T.; Leite, R.P.; Nishioka, S.A.; Avila, A.; Sano‑Martins, I.S.; Tomy, S.C.; Santoro, M.L.; et al. Randomized comparative trial of three antivenoms in the treatment of envenoming by lance-headed vipers (Bothrops jararaca) in São Paulo, Brazil. Q. J. Med. 1993, 86, 315–325. [Google Scholar]
- Theakston, R.D.G.; Fan, H.W.; Warrell, D.A.; Dias da Silva, W.D.; Ward, S.A.; Higashi, H.G.; B.I.A.S.G. Use of enzyme immunoassays to compare the effect and assess the dosage regimens of three Brazilian Bothrops antivenoms. Am. J. Trop. Med. Hyg. 1992, 47, 593–604. [Google Scholar]
- Jorge, M.T.; Cardoso, J.L.C.; Castro, S.C.B.; Ribeiro, L.; França, F.O.S.; Sbroglio de Almeida, M.E.; Kamiguti, A.S.; Sano-Martins, I.S.; Santoro, M.L.; Moncau, J.E.C.; et al. A randomised blinded comparison of two doses of antivenom in the treatment of Bothrops envenoming in São Paulo, Brazil. Trans. R. Soc. Trop. Med. Hyg. 1990, 89, 111–114. [Google Scholar]
- Lalloo, D.G.; Trevett, A.J.; Korinhona, A.; Nwokolo, N.C.; Laurenson, I.F.; Paul, M.; Black, J.; Naraqi, S.; Mavo, B.; Saweri, A.; et al. nakebites by the Papuan Taipan (Oxyuranus scutellatus canni); paralysis, haemostatic and electrocardiographic abnormalities and effects of antivenom. Am. J. Trop. Med. Hyg. 1995, 52, 525–531. [Google Scholar]
- Ho, M.; Silamut, K.; White, N.J.; Karbwang, J.; Looareesuwan, S.; Phillips, R.E.; Warrell, D.A. Pharmacokinetics of three commercial antivenoms in patients envenomed by the Malayan pit viper, Calloselasma rhodostoma, in Thailand. Am. J. Trop. Med. Hyg. 1990, 42, 260–266. [Google Scholar]
- Gillissen, A.; Theakston, R.D.; Barth, J.; May, B.; Krieg, M.; Warrell, D.A. Neurotoxicity, haemostatic disturbances and haemolytic anaemia after a bite by a Tunisian saw-scaled or carpet viper (Echis “pyramidum”-complex): Failure of antivenom treatment. Toxicon 1994, 32, 937–944. [Google Scholar] [CrossRef]
- Krifi, M.N.; Savin, S.; Debray, M.; Bon, C.; El Ayeb, M.; Choumet, V. Pharmacokinetic studies of scorpion venom before and after antivenom immunotherapy. Toxicon 2005, 45, 187–198. [Google Scholar] [CrossRef]
- Chippaux, J.P.; Ramos-Cerrillo, B.; Stock, R.P. Study of the efficacy of the black stone on envenomation by snake bite in the murine model. Toxicon 2007, 49, 717–720. [Google Scholar] [CrossRef]
- Sutherland, S.K.; Coulter, A.R.; Harris, R.D. Rationalisation of first-aid measures for elapid snakebite. Lancet 1979, I, 183–185. [Google Scholar] [CrossRef]
- Pugh, R.N.H.; Theakston, R.D.G. The incidence and mortality of snake bite in savanna Nigeria. Lancet 1980, 2, 1181–1183. [Google Scholar] [CrossRef]
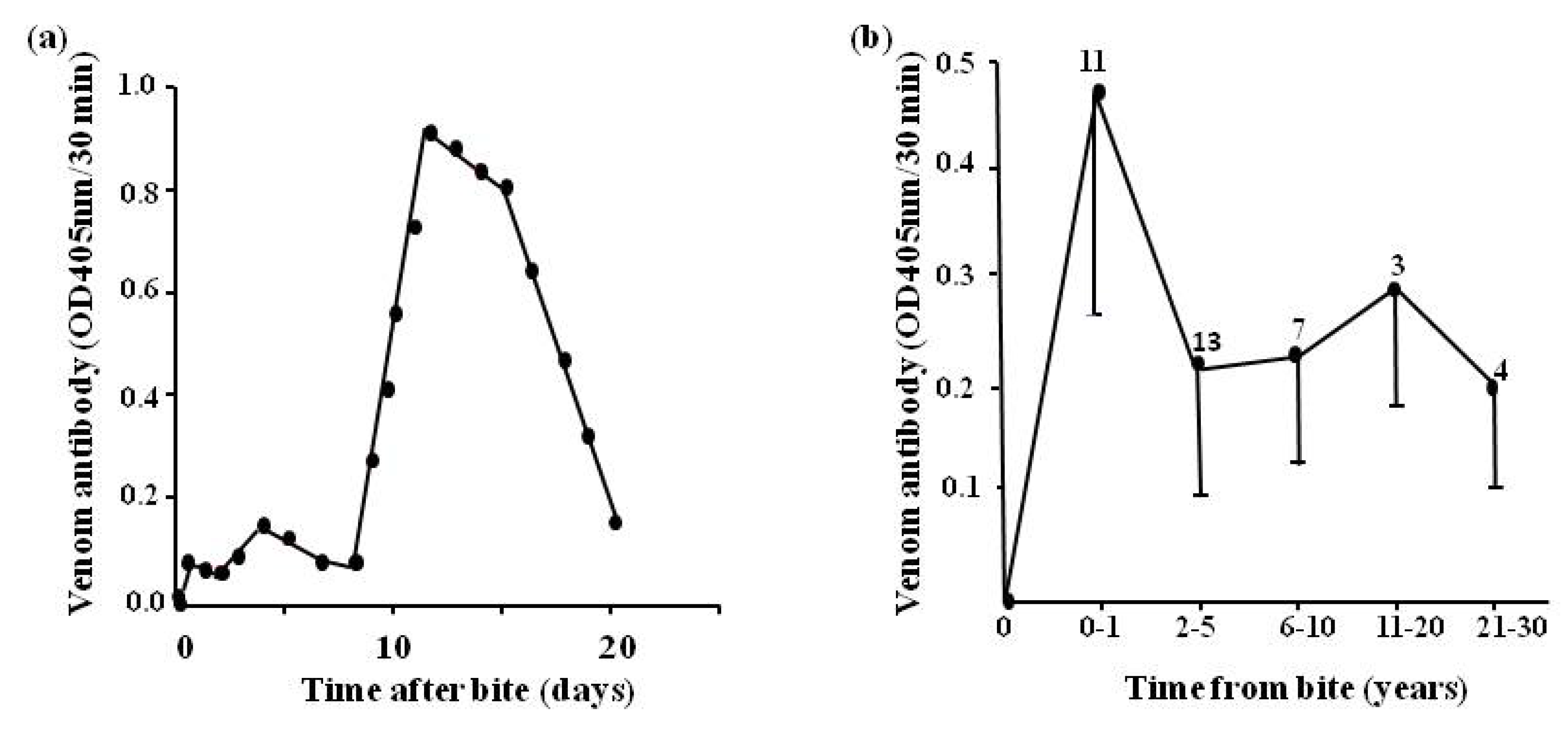
- Theakston, R.D.G.; Reid, H.A.; Larrick, J.W.; Yost, J.A. Snake venom antibodies in Ecuadorian Indians. J. Trop. Med. Hyg. 1981, 84, 199–202. [Google Scholar]
- Warrell, D.A.; Arnett, C. The importance of bites by the saw-scaled or carpet viper (Echis carinatus). Acta Trop. 1976, 33, 307–341. [Google Scholar]
- Warrell, D.A.; University of Oxford, Oxford, UK. Personal communication, 1979.
- Chippaux, J.P.; Theakston, R.D.G. Epidemiological studies of snake bite in French Guiana. Ann. Trop. Med. Parasitol. 1987, 81, 301–304. [Google Scholar]
- Theakston, R.D.G.; Reid, H.A.; Iddon, D.; Larrick, J.W. Protective effect of snake venom antibodies in sera of previous snake bite victims. Ann. Trop. Med. Parasitol. 1983, 77, 311–314. [Google Scholar]
- Paine, M.J.I; Desmond, H.P.; Theakston, R.D.G.; Crampton, J.M. Purification, cloning and molecular characterization of a high molecular weight hemorrhagic metaloprotease from Bothrops jararaca venom—Insights into the Disintegrin gene family. J. Biol. Chem. 1992, 267, 22869–22876. [Google Scholar]
- Hawgood, B.J. Hugh Alistair Reid OBE, MD: Investigation and treatment of snake bite. Toxicon 1998, 36, 431–446. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Theakston, R.D.G.; Laing, G.D. Diagnosis of Snakebite and the Importance of Immunological Tests in Venom Research. Toxins 2014, 6, 1667-1695. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins6051667
Theakston RDG, Laing GD. Diagnosis of Snakebite and the Importance of Immunological Tests in Venom Research. Toxins. 2014; 6(5):1667-1695. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins6051667
Chicago/Turabian StyleTheakston, R. David G., and Gavin D. Laing. 2014. "Diagnosis of Snakebite and the Importance of Immunological Tests in Venom Research" Toxins 6, no. 5: 1667-1695. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins6051667