
The Cytolytic Activity of Vaginolysin Strictly Depends on Cholesterol and Is Potentiated by Human CD59

Abstract
:1. Introduction
2. Results
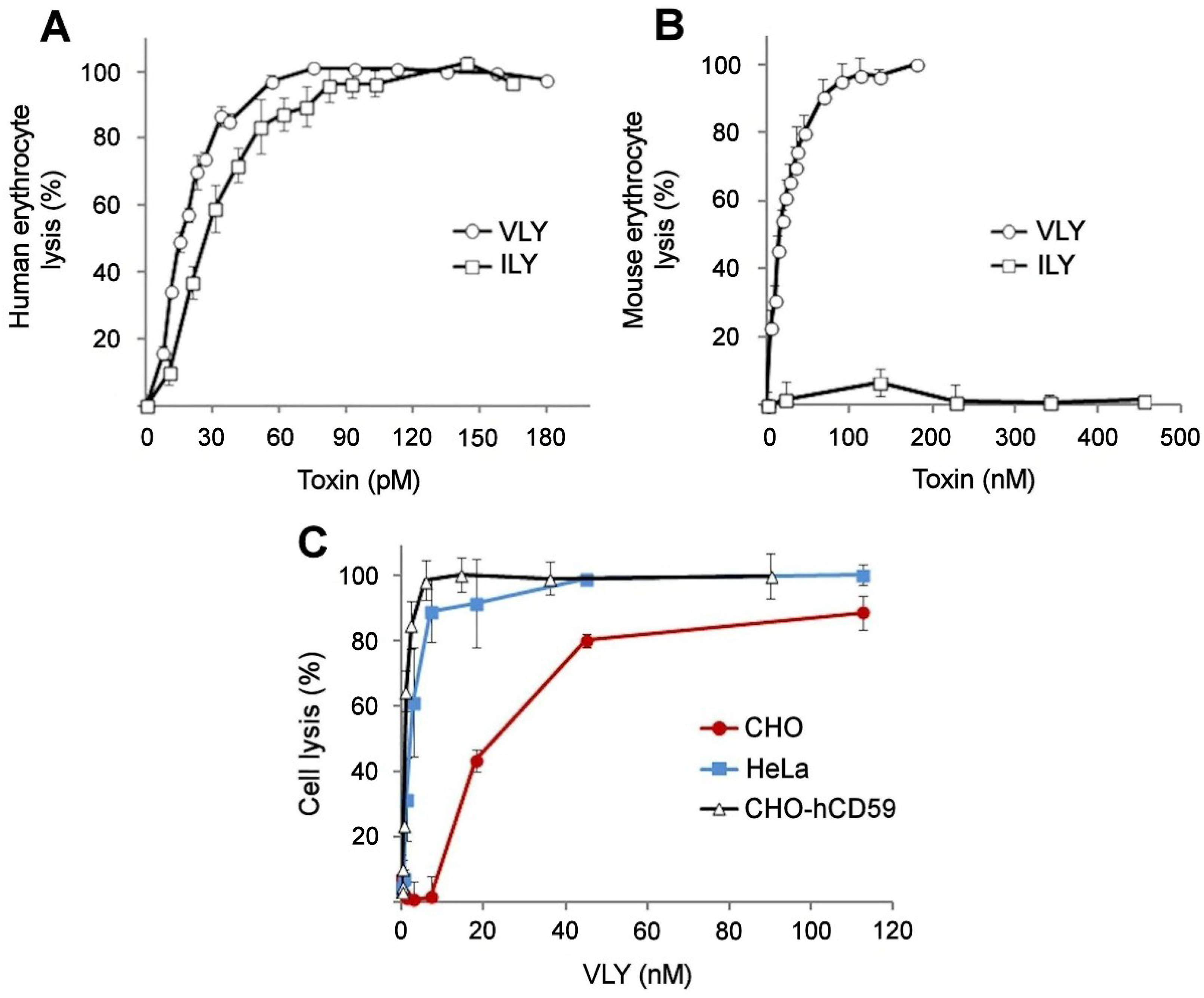
2.1. VLY Cytolytic Activity on Human and Non-Human Cells

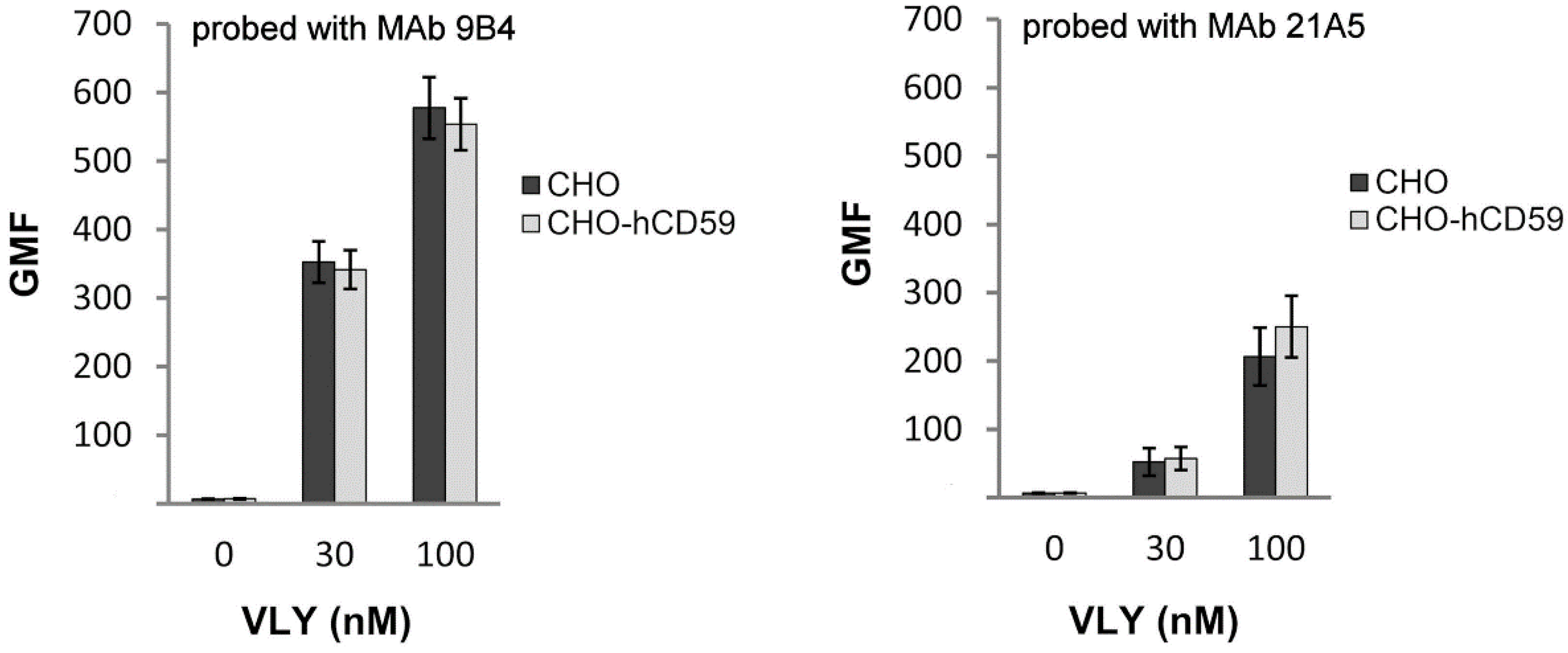
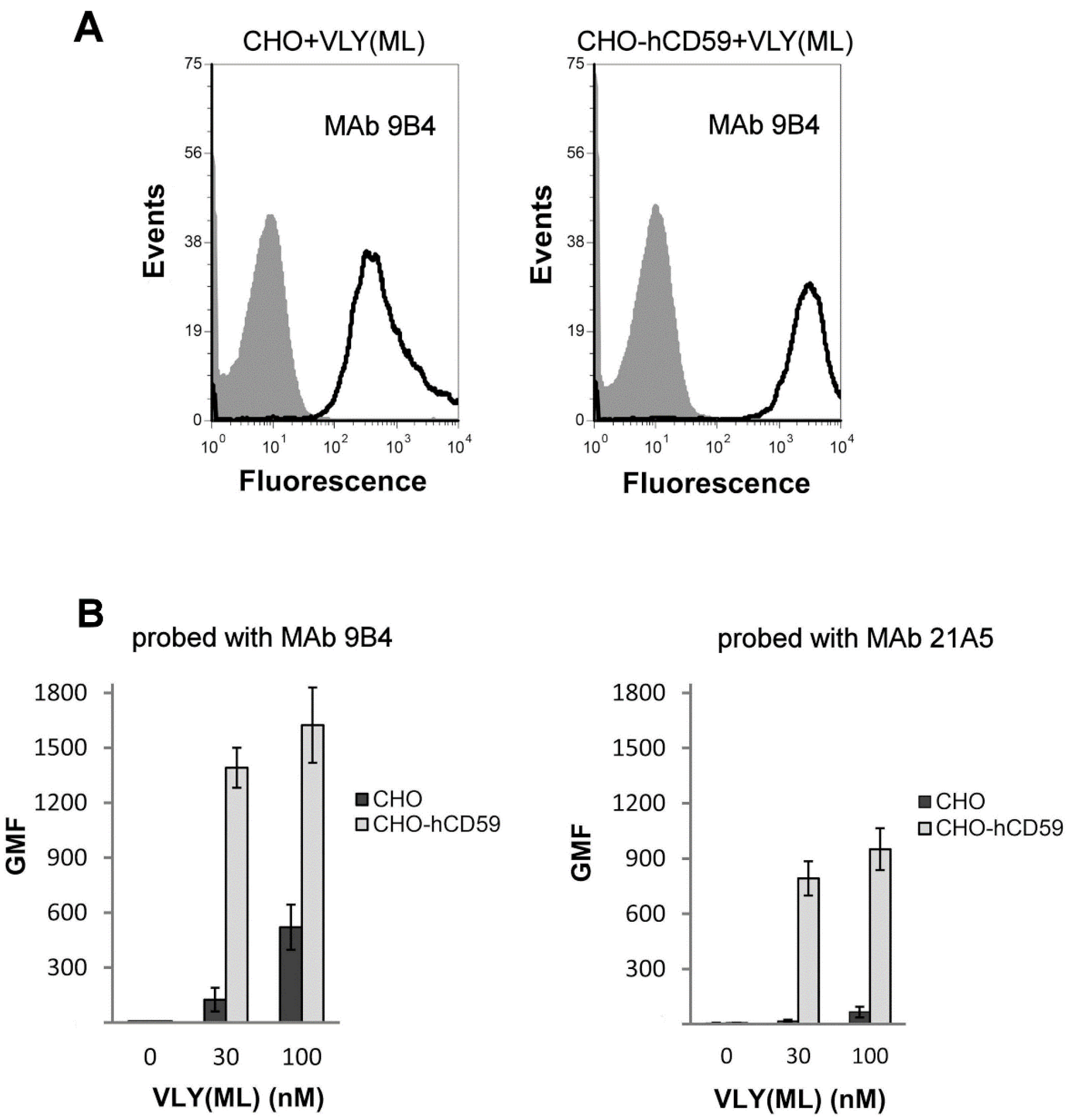
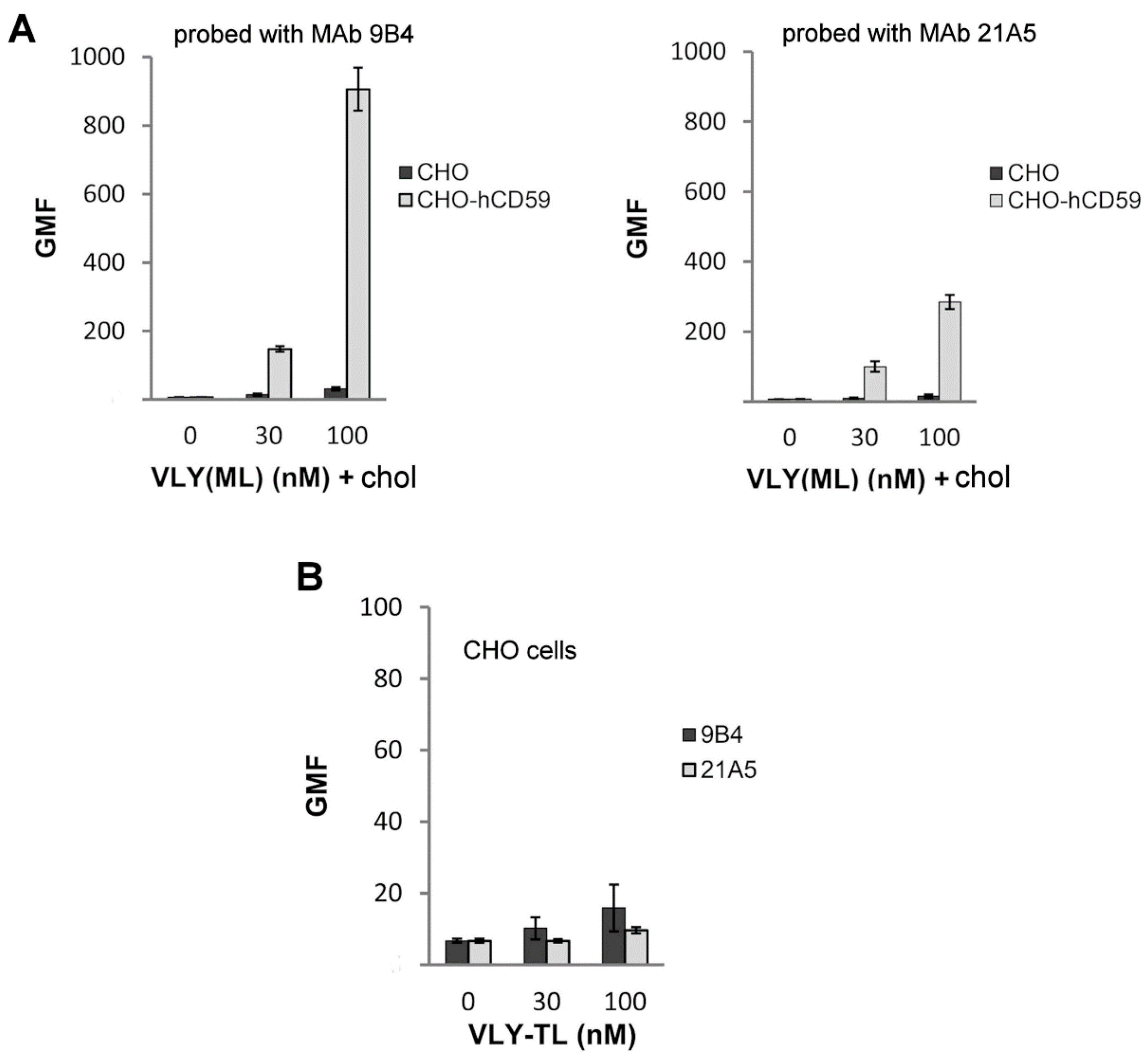
2.2. VLY Binding to CHO and CHO-hCD59 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VLY variants | Cholesterol binding (nmol cholesterol) a | Oligomer formation detected by denaturing agarose gel electrophoresis | HD50 (pM) b |
|---|---|---|---|
| VLY | 0.25 | + | 10 ± 1 |
| VLY(ML): G308A·G309A | 0.25 | - | >2000 |
| VLY-TL: T474G·L475G | >65 | - | >2000 |
| Y165A | 0.25 | + | 18 ± 1 |
| I306P | 0.25 | - | >1000 |
| A304P·I306P | 1.02 | - | >1000 |
| V305P | 0.25 | + | 17 ± 5 |
| MAbs | Native VLY | VLY(ML) | VLY-TL |
|---|---|---|---|
| 9B4 | 3 × 10−10 M | 2.2 × 10−10 M | 2.5 × 10−10 M |
| 21A5 | 2.7 × 10−10 M | 0.9 × 10−9 M | 0.7 × 10−9 M |



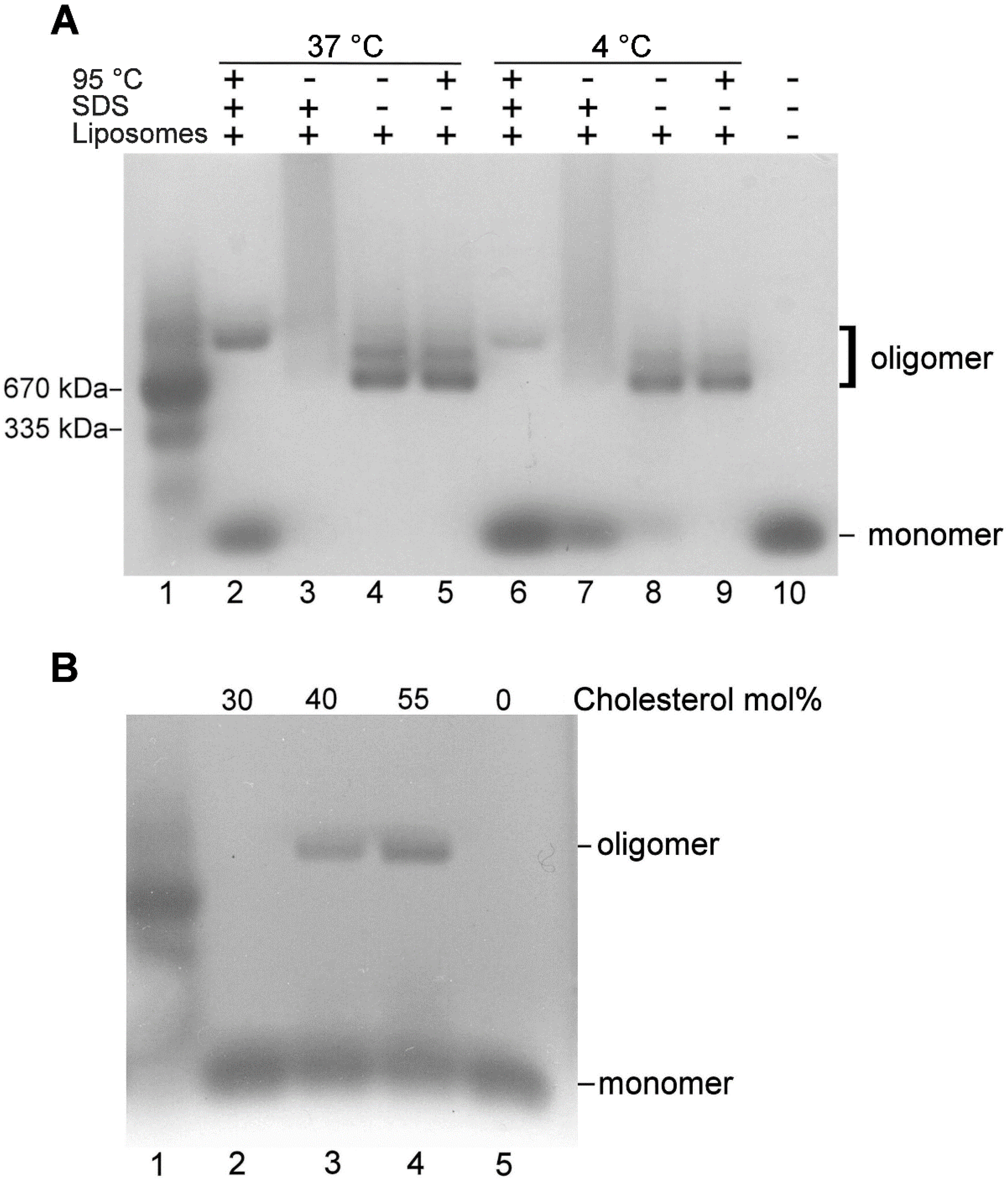
2.3. VLY Oligomer Formation

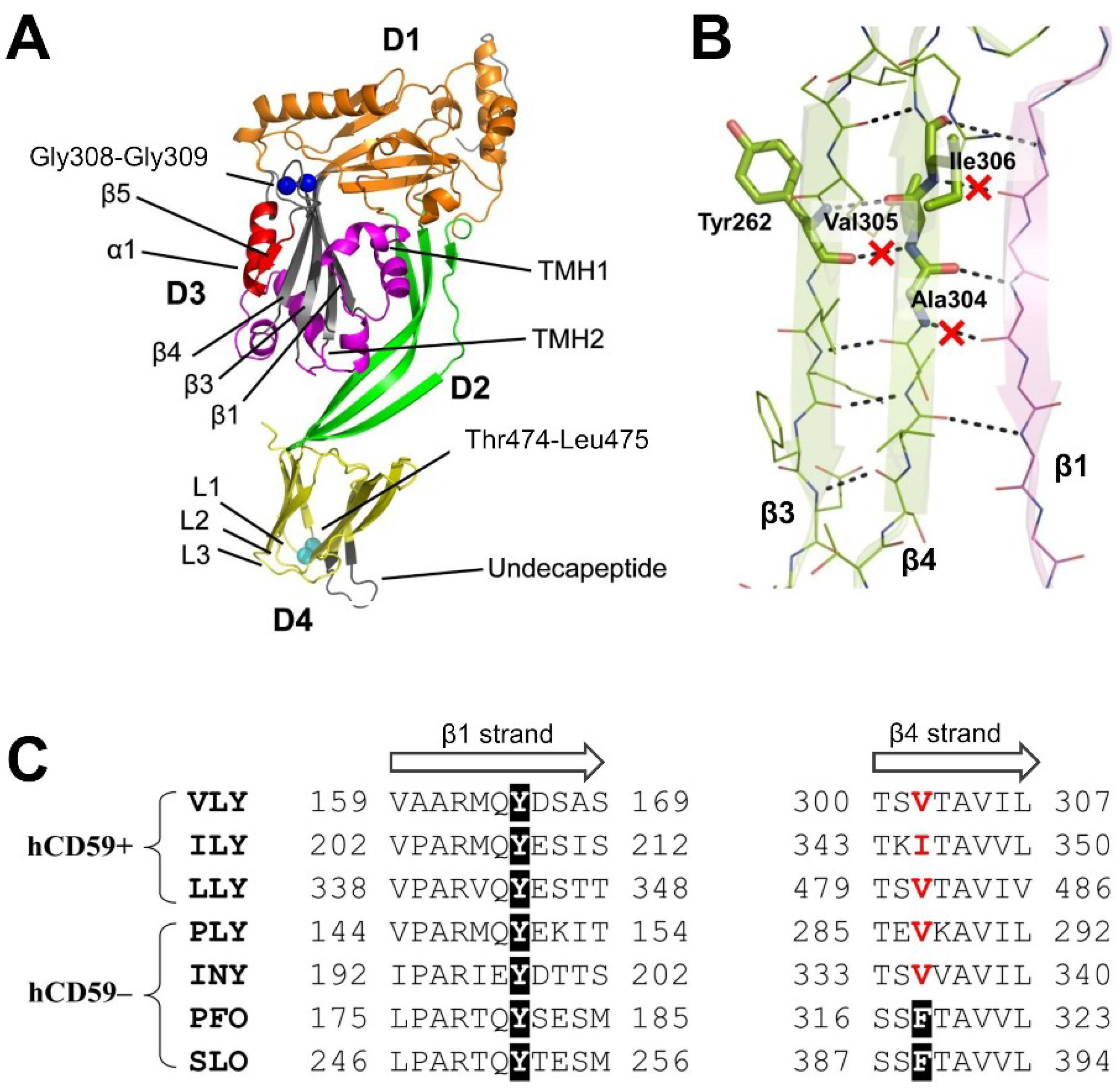
2.4. Analysis of VLY Monomer-Monomer Interface

3. Discussion
4. Experimental Section
4.1. Generation of Vaginolysin Mutants
4.2. Generation of Pneumolysin and Intermedilysin
4.3. Determination of VLY and ILY Hemolytic Activity
4.4. Determination of VLY Cytolytic Activity
4.5. Generation of CHO Cell Line Expressing hCD59
4.6. Flow Cytometry
4.7. Indirect ELISA
4.8. Preparation of Liposomes
4.9. Cholesterol Dot Blot Analysis
4.10. Denaturing Agarose Gel Electrophoresis
4.11. Construction and Assessment of the Homology-Based VLY Structural Model
4.12. Ethics Statement
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tweten, R.K. Cholesterol-dependent cytolysins, a family of versatile pore-forming toxins. Infect. Immun. 2005, 73, 6199–6209. [Google Scholar] [CrossRef] [PubMed]
- Los, F.C.O.; Randis, T.M.; Aroian, R.V.; Ratner, A.J. Role of pore-forming toxins in bacterial infectious diseases. Microbiol. Mol. Biol. Rev. 2013, 44, 173–207. [Google Scholar] [CrossRef]
- Ramachandran, R.; Tweten, R.K.; Johnson, A.E. Membrane-dependent conformational changes initiate cholesterol-dependent cytolysin oligomerization and intersubunit β-strand alignment. Nat. Struct. Mol. Biol. 2004, 11, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.; Chattopadhyay, K. Single point mutation in Vibrio cholera cytolysin compromises the membrane pore-formation mechanism of the toxin. FEBS J. 2012, 279, 4039–4051. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.; Harris, R.; Freytag, C.; Kehoe, M.; Tranum-Jensen, J.; Bhakdi, S. Assembly mechanism of the oligomeric streptolysin O pore: The early membrane lesion is lined by a free edge of the lipid membrane and is extended gradually during oligomerization. EMBO J. 1998, 17, 1598–1605. [Google Scholar] [CrossRef] [PubMed]
- Sonnen, A.F.; Plitzko, J.M.; Gilbert, R.J. Incomplete pneumolysin oligomers form membrane pores. Open Biol. 2014, 4, 140044. [Google Scholar] [CrossRef] [PubMed]
- Bavdek, A.; Kostanjšek, R.; Antonini, V.; Lakey, J.H.; Dalla Serra, M.; Gilbert, R.J.; Anderluh, G. pH dependence of listeriolysin O aggregation and pore-forming ability. FEBS J. 2012, 279, 126–141. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.J.; Tweten, R.K.; Johnson, A.E.; Heuck, A.P. Cholesterol exposure at the membrane surface is necessary and sufficient to trigger perfringolysin O binding. Biochemistry 2009, 48, 3977–3987. [Google Scholar] [CrossRef] [PubMed]
- Farrand, A.J.; LaChapelle, S.; Hotze, E.M.; Johnson, A.E.; Tweten, R.K. Only two amino acids are essential for cytolytic toxin recognition of cholesterol at the membrane surface. Proc. Natl. Acad. Sci. USA 2010, 107, 4341–4346. [Google Scholar] [CrossRef] [PubMed]
- Heuck, A.P.; Hotze, E.M.; Tweten, R.K.; Johnson, A.E. Mechanism of membrane insertion of a multimeric beta-barrel protein: Perfringolysin O creates a pore using ordered and coupled conformational changes. Mol. Cell 2000, 6, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Hotze, E.M.; Wilson-Kubalek, E.; Farrand, A.J.; Bentsen, L.; Parker, M.W.; Johnson, A.E.; Tweten, R.K. Monomer-monomer interactions propagate structural transitions necessary for pore formation by the cholesterol-dependent cytolysins. J. Biol. Chem. 2012, 287, 24534–24543. [Google Scholar] [CrossRef] [PubMed]
- Giddings, K.S.; Zhao, J.; Sims, P.J.; Tweten, R.K. Human CD59 is a receptor for the cholesterol-dependent cytolysin intermedilysin. Nat. Struct. Mol. Biol. 2004, 11, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Soltani, C.E.; Hotze, E.M.; Johnson, A.E.; Tweten, R.K. Specific protein-membrane contacts are required for prepore and pore assembly by a cholesterol-dependent cytolysin. J. Biol. Chem. 2007, 282, 15709–15716. [Google Scholar] [CrossRef] [PubMed]
- Rollins, S.A.; Sims, P.J. The complement-inhibitory activity of CD59 resides in its capacity to block incorporation of C9 into membrane C5b-9. J. Immunol 1990, 144, 3478–3483. [Google Scholar] [PubMed]
- Kim, D.D.; Song, W.C. Membrane complement regulatory proteins. Clin. Immunol. 2006, 118, 127–136. [Google Scholar] [CrossRef] [PubMed]
- LaChapelle, S.; Tweten, R.K.; Hotze, E.M. Intermedilysin-receptor interactions during assembly of the pore complex. J. Biol. Chem. 2009, 284, 12719–12726. [Google Scholar] [CrossRef] [PubMed]
- Nagamune, H.; Ohnishi, C.; Katsuura, A.; Fushitani, K.; Whiley, R.A.; Tsuji, A.; Matsuda, Y. Intermedilysin, a novel cytotoxin specific for human cells secreted by Streptococcus intermedius UNS46 isolated from a human liver abscess. Infect. Immun. 1996, 64, 3093–3100. [Google Scholar] [PubMed]
- Gelber, S.E.; Aguilar, J.L.; Lewis, K.L.; Ratner, A.J. Functional and phylogenetic characterization of vaginolysin, the human-specific cytolysin from Gardnerella vaginalis. J. Bacteriol. 2008, 190, 3896–3903. [Google Scholar] [CrossRef] [PubMed]
- Wickham, S.E.; Hotze, E.M.; Farrand, A.J.; Polekhina, G.; Nero, T.L.; Tomlinson, S.; Parker, M.W.; Tweten, R.K. Mapping the intermedilysin-human CD59 receptor interface reveals a deep correspondence with the binding site on CD59 for complement binding proteins C8α and C9. J. Biol. Chem. 2011, 286, 20952–20962. [Google Scholar] [CrossRef] [PubMed]
- Menard, J.P.; Fenollar, F.; Henry, M.; Bretelle, F.; Raoult, D. Molecular quantification of Gardnerella vaginalis and Atopobium vaginae loads to predict bacterial vaginosis. Clin. Infect. Dis. 2008, 47, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Zvirbliene, A.; Pleckaityte, M.; Lasickiene, R.; Kucinskaite-Kodze, I.; Zvirblis, G. Production and characterization of monoclonal antibodies against vaginolysin: Mapping of a region critical for its cytolytic activity. Toxicon 2010, 56, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Pleckaityte, M.; Janulaitiene, M.; Lasickiene, R.; Zvirbliene, A. Genetic and biochemical diversity of Gardnerella vaginalis strains isolated from women with bacterial vaginosis. FEMS Immunol. Med. Microbiol. 2012, 65, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Tabata, A.; Ohkura, K.; Ohkubo, Y.; Tomoyasu, T.; Ohkuni, H.; Whiley, R.A.; Nagamune, H. The diversity of receptor recognition in cholesterol dependent cytolysins. Microbiol. Immunol. 2014, 58, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Budvytyte, R.; Pleckaityte, M.; Zvirbliene, A.; Vanderah, D.J.; Valincius, G. Reconstitution of cholesterol-dependent vaginolysin into tethered phospholipid bilayers: Implications for bioanalysis. PLoS One 2013, 8, e82536. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, N.M.; Lewis, W.G.; Lewis, A.L. Clinical features of bacterial vaginosis in a murine model of vaginal infection with Gardnerella vaginalis. PLoS One 2013, 8, e59539. [Google Scholar] [CrossRef] [PubMed]
- Dowd, K.J.; Tweten, R.K. The cholesterol-dependent cytolysin signature motif: A critical element in the allosteric pathway that couples membrane binding to pore assembly. PLoS Pathog. 2012, 8, e1002787. [Google Scholar] [CrossRef] [PubMed]
- Shepard, L.A.; Shatursky, O.; Johnson, A.E.; Tweten, R.K. The mechanism of pore assembly for a cholesterol-dependent cytolysin: Formation of a large prepore complex precedes the insertion of the transmembrane beta-hairpins. Biochemistry 2000, 39, 10284–10293. [Google Scholar] [CrossRef] [PubMed]
- Hotze, E.M.; Tweten, R.K. Membrane assembly of the cholesterol-dependent cytolysin pore complex. Biochim. Biophys. Acta 2012, 1818, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Polekhina, G.; Giddings, K.S.; Tweten, R.K.; Parker, M.W. Insights into the action of the superfamily of cholesterol-dependent cytolysins from studies of intermedilysin. Proc. Natl. Acad. Sci. USA 2005, 102, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Farrand, S.; Hotze, E.; Friese, P.; Hollingshead, S.K.; Smith, D.F.; Cummings, R.D.; Dale, G.L.; Tweten, R.K. Characterization of a streptococcal cholesterol-dependent cytolysin with a Lewis y and b specific lectin domain. Biochemistry 2008, 47, 7097–7107. [Google Scholar] [CrossRef] [PubMed]
- Waheed, A.A.; Shimada, Y.; Heijnen, H.F.; Nakamura, M.; Inomata, M.; Hayashi, M.; Iwashita, S.; Slot, J.W.; Ohno-Iwashita, Y. Selective binding of perfringolysin O derivative to cholesterol-rich membrane microdomains (rafts). Proc. Natl. Acad. Sci. USA 2001, 98, 4926–4931. [Google Scholar] [CrossRef] [PubMed]
- Gekara, N.O.; Jacobs, T.; Chakraborty, T.; Weiss, S. The cholesterol-dependent cytolysin listeriolysin O aggregates rafts via oligomerization. Cell Microbiol. 2005, 7, 1345–1356. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.D.; Sanders, M.E.; Tullos, N.A.; Stray, S.J.; Norcross, E.W.; McDaniel, L.S.; Marquart, M.E. The cholesterol-dependent cytolysin pneumolysin from Streptococcus pneumoniae binds to lipid raft microdomains in human corneal epithelial cells. PLoS One 2013, 8, e61300. [Google Scholar] [CrossRef] [PubMed]
- Nelson, L.D.; Chiantia, S.; London, E. Perfringolysin O association with ordered lipid domains: Implications for transmembrane protein raft affinity. Biophys. J. 2010, 99, 3255–3263. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.B.; Moe, P.C.; Wang, D.; Rossi, K.; Trigatti, B.L.; Heuck, A.P. Modifications in perfringolysin O domain 4 alter the cholesterol concentration threshold required for binding. Biochemistry 2012, 51, 3373–3382. [Google Scholar] [CrossRef] [PubMed]
- Kimberley, F.C.; Sivasankar, B.; Morgan, P.B. Alternative roles for CD59. Mol. Immunol. 2007, 44, 73–81. [Google Scholar] [CrossRef] [PubMed]
- LaRocca, T.J.; Stivison, E.A.; Hod, E.A.; Spitalnik, S.L.; Cowan, P.J.; Randis, T.M.; Ratner, A.J. Human-specific bacterial pore-forming toxins induce programmed necrosis in erythrocytes. MBio 2014, 5, e01251-14. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Abagyan, R.; Dong, S.; Gilbert, A.; Nussenzweig, V.; Tomlinson, S. Mapping of the active site of CD59. J. Exp. Med. 1997, 185, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Cizas, P.; Budvytyte, R.; Morkuniene, R.; Moldovan, R.; Broccio, M.; Lösche, M.; Niaura, G.; Valincius, G.; Borutaite, V. Size-dependent neurotoxicity of beta-amyloid oligomers. Arch. Biochem. Biophys. 2010, 496, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acid Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [PubMed]
- Rossjohn, J.; Feil, S.C.; McKinstry, W.J.; Tweten, R.K.; Parker, M.W. Structure of a cholesterol-binding, thiol-activated cytolysin and a model of its membrane form. Cell 1997, 89, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Sippl, M.J. Recognition of errors in three-dimensional structures of proteins. Proteins 1993, 17, 355–362. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zilnyte, M.; Venclovas, Č.; Zvirbliene, A.; Pleckaityte, M. The Cytolytic Activity of Vaginolysin Strictly Depends on Cholesterol and Is Potentiated by Human CD59. Toxins 2015, 7, 110-128. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7010110
Zilnyte M, Venclovas Č, Zvirbliene A, Pleckaityte M. The Cytolytic Activity of Vaginolysin Strictly Depends on Cholesterol and Is Potentiated by Human CD59. Toxins. 2015; 7(1):110-128. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7010110
Chicago/Turabian StyleZilnyte, Milda, Česlovas Venclovas, Aurelija Zvirbliene, and Milda Pleckaityte. 2015. "The Cytolytic Activity of Vaginolysin Strictly Depends on Cholesterol and Is Potentiated by Human CD59" Toxins 7, no. 1: 110-128. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7010110