
Low Toxicity of Deoxynivalenol-3-Glucoside in Microbial Cells

Abstract
:1. Introduction

2. Results
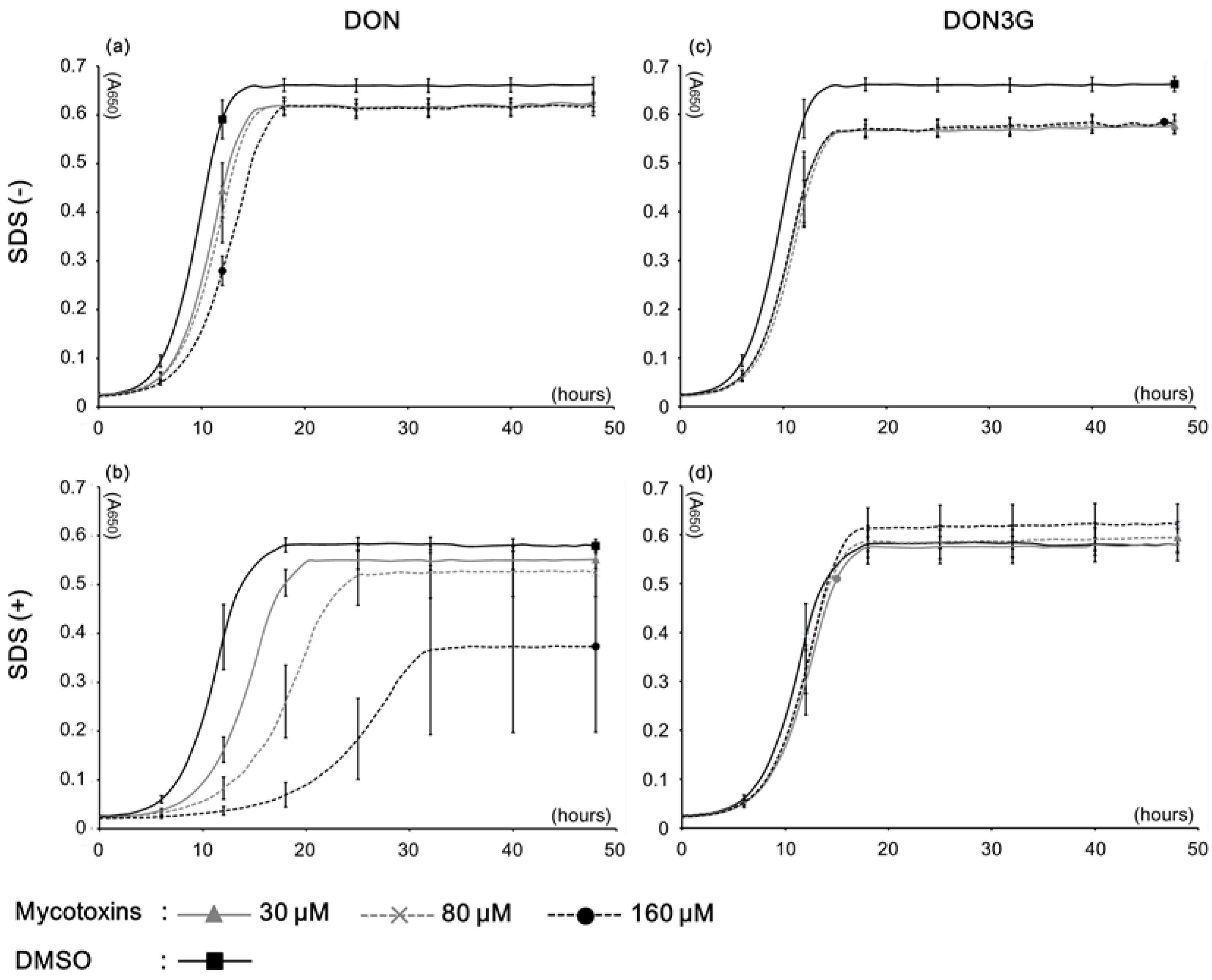
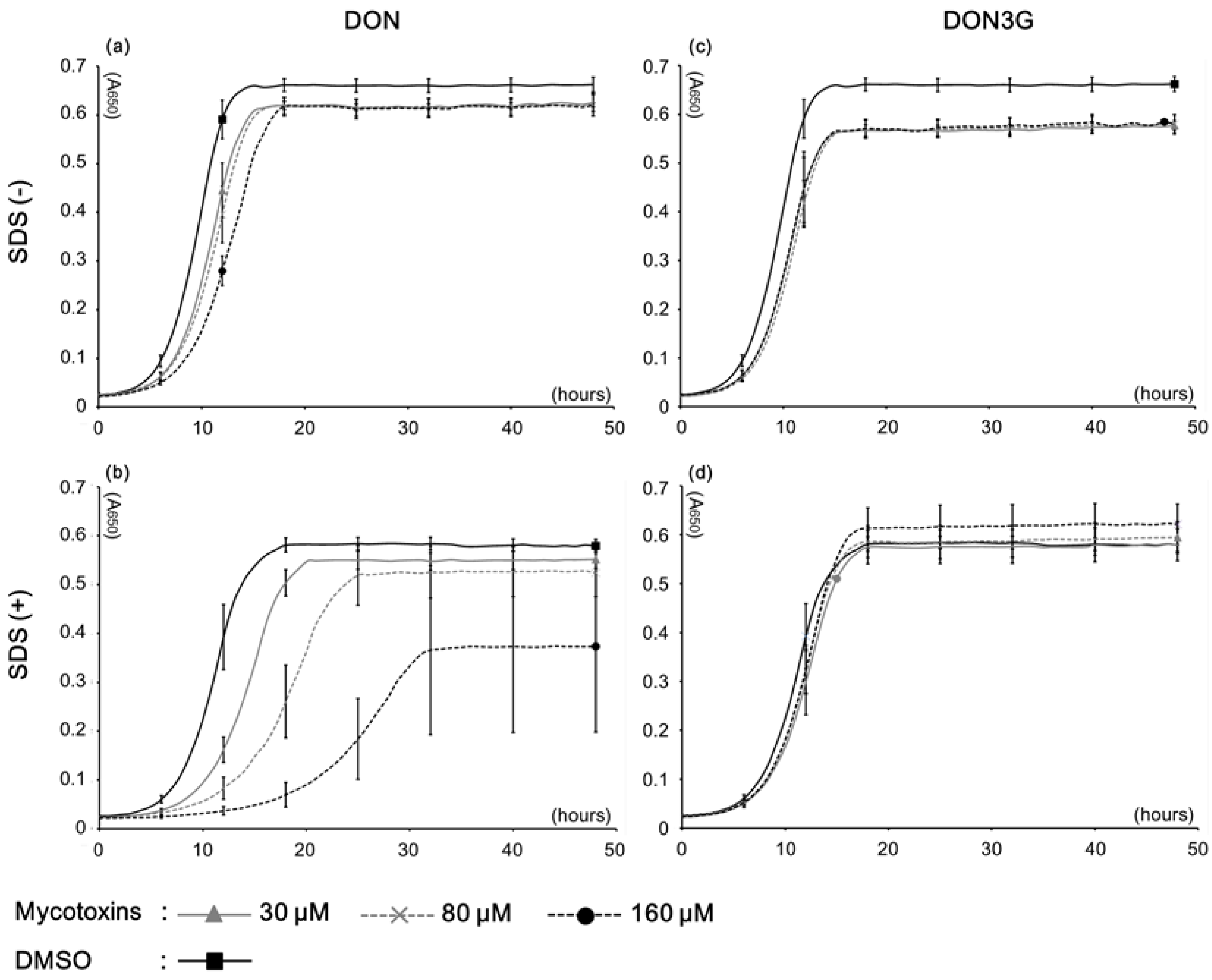
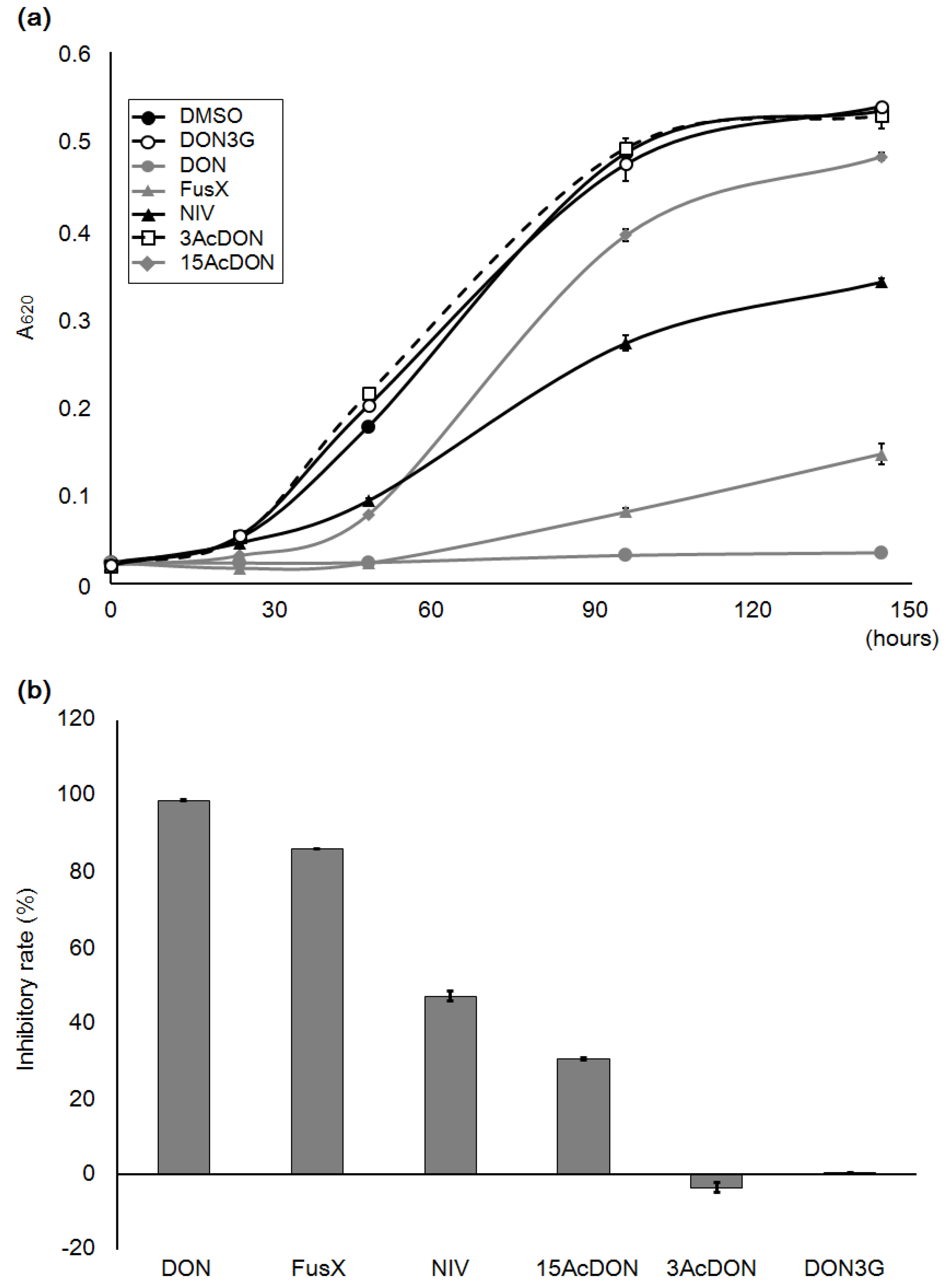
2.1. Yeast Growth with DON3G
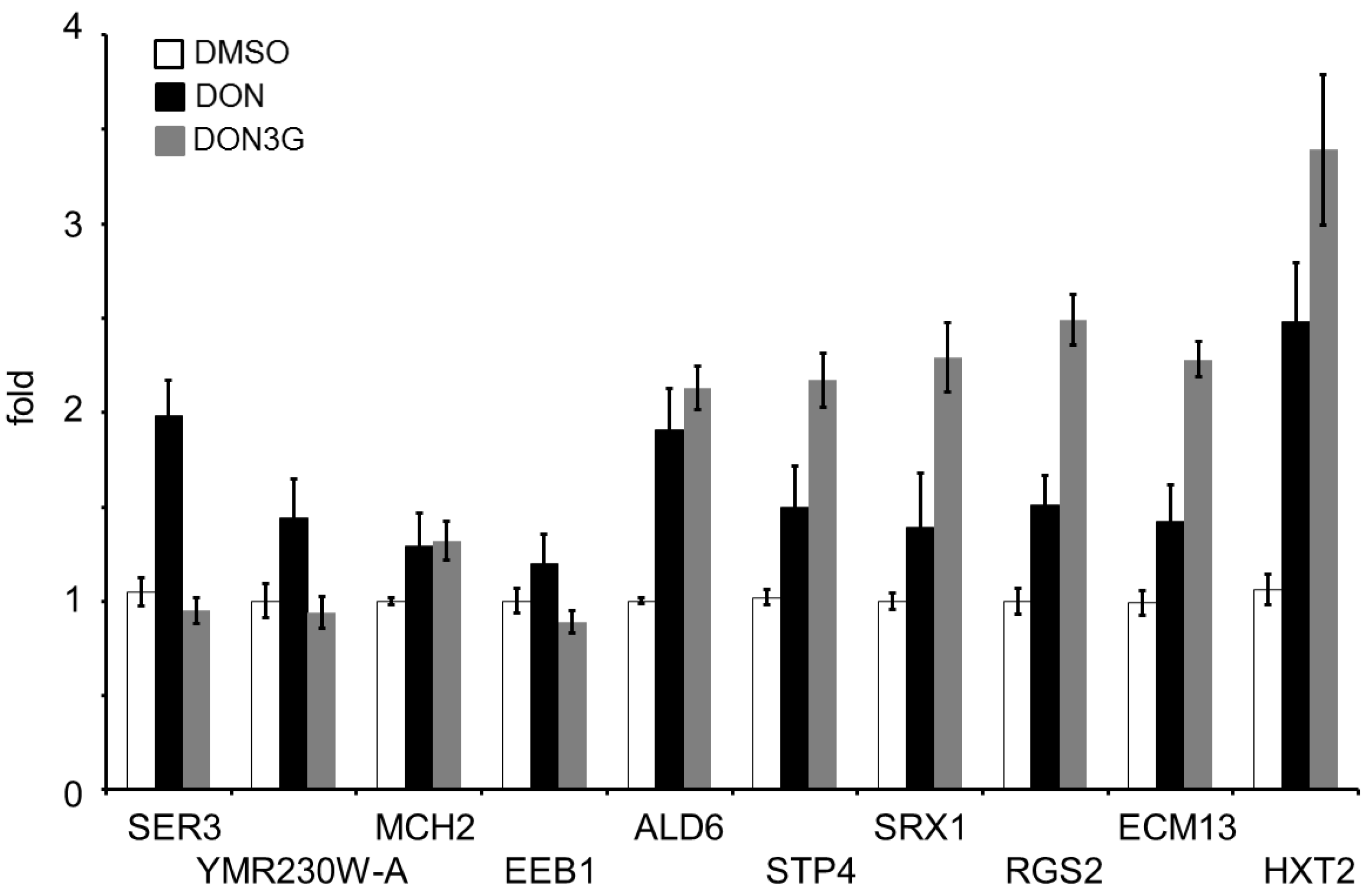
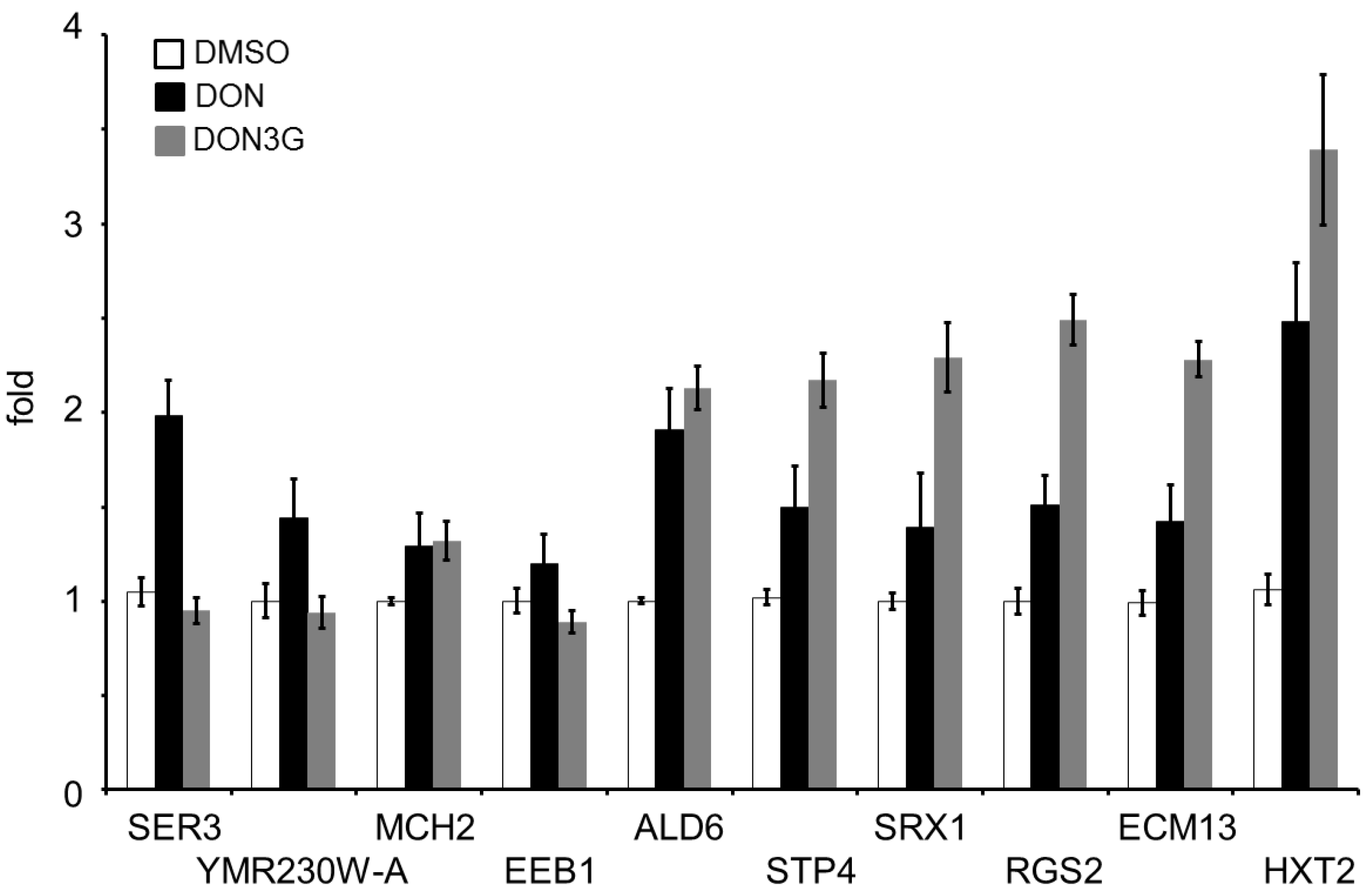
2.2. DON3G Influences a Small Number of Genes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium condition | Mycotoxin | Concentrations (μM) | IC20 * (μM) | ||
|---|---|---|---|---|---|
| 30 | 80 | 160 | |||
| SDS (−) | DON | 13.3 ± 0.64 ** | 14.4 ± 0.15 | 18.9 ± 0.08 | >160 |
| DON3G | 19.7 ± 0.98 | 20.2 ± 0.84 | 19.3 ± 2.09 | - *** | |
| SDS (+) | DON | 21.1 ± 1.58 | 37.4 ± 8.72 | 71.7 ± 13.54 | 30.4 |
| DON3G | 9.7 ± 3.84 | 7.7 ± 1.03 | 2.7 ± 2.45 | - | |
| Systematic No. | Gene Symbol | Fold Change | Gene Title | Description |
|---|---|---|---|---|
| YER081W | SER3 | 0.31 | 3-phosphoglycerate dehydrogenase | Serine and glycine biosynthesis |
| YMR230W-A | - | 0.35 | Putative protein of unknown function | - |
| YKL221W | MCH2 | 0.41 | Protein with similarity to mammalian monocarboxylate permeases | Monocarboxylate transport |
| YPL095C | EEB1 | 0.47 | Acyl-coenzymeA: ethanol O-acyltransferase | Fatty acid ethyl ester biosynthesis |
| YPL061W | ALD6 | 2.06 | Cytosolic aldehyde dehydrogenase | Oxidative stress response |
| YDL048C | STP4 | 2.25 | Protein containing a Kruppel-type zinc-finger domain | DNA replication stress |
| YKL086W | SRX1 | 2.57 | Sulfiredoxin | Oxidative stress resistance DNA replication stress |
| YOR107W | RGS2 | 3.15 | Negative regulator of glucose-induced cAMP signaling | Glucose response |
| YBL043W | ECM13 | 3.39 | Non-essential protein of unknown function | UVA irradiation response |
| YMR011W | HXT2 | 5.66 | High-affinity glucose transporter | Induced by low levels of glucose Repressed by high levels of glucose |
2.3. Real-Time Polymerase Chain Reaction Analysis
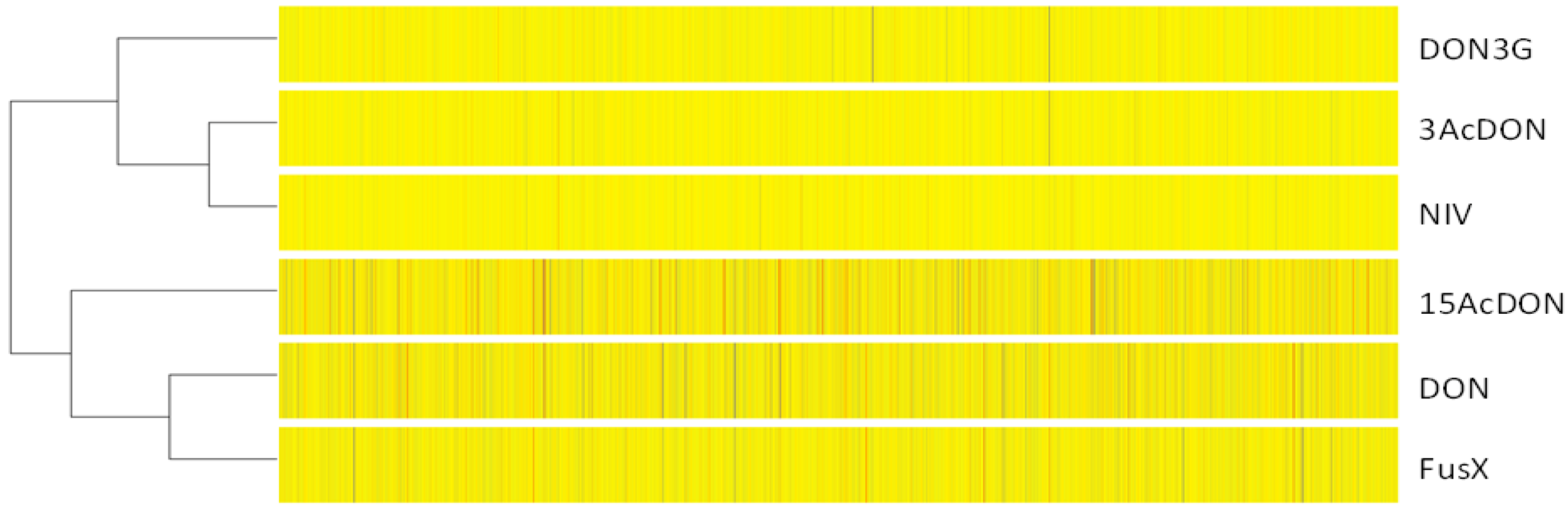
2.4. Clustering Analysis

2.5. Growth Test with Algal Cells

3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Biomaterials
4.3. Yeast Growth
4.4. DNA Microarray Analysis
4.5. Real-Time PCR Analysis
4.6. C. reinhardtii Growth
| Gene symbol | Direction | Sequence | Product size (bp) |
|---|---|---|---|
| SER3 | forward | TCACCAAAATGTACCAGGTGT | 227 |
| reverse | TGACAGTATGAGAATCGATTGCA | ||
| YMR230W-A | forward | GGATGTGTTACGATGCAGACA | 170 |
| reverse | ATATGGCGCGTTCTTGAAGG | ||
| MCH2 | forward | CGTGGGTTTTGCGTACTTTG | 194 |
| reverse | GTGACCTTTAGACTCTCCTAGGT | ||
| EEB1 | forward | TCCAGTTACAGGTGAAAACGT | 164 |
| reverse | ACTCATCAAAGCTGCCCAAG | ||
| ALD6 | forward | AGATGTTGAAGGCCGGTACC | 185 |
| reverse | TGACGGAAAGAAATGCAGGT | ||
| STP4 | forward | TTTGCATTTCGAGTACCCGC | 183 |
| reverse | TGTGTGTATGTATGAGTCGGTG | ||
| SRX1 | forward | CTGGGCGTGCGAGTCAAG | 176 |
| reverse | ATGTCGAGACTGCTGCCC | ||
| RGS2 | forward | AGGAATTCTCAACTCGGGGA | 173 |
| reverse | TCCACAGATGATGAAGAGGCT | ||
| ECM13 | forward | CGAGCAGACGGATGAACTTG | 214 |
| reverse | TACGGAACCATCGTCGACAT | ||
| HXT2 | forward | GGGTATGTCTTCATGGGCTGT | 180 |
| reverse | TATAATCTCTTATTCCTCGGAAACTC | ||
| PDR5 | forward | ACAGTGAGAGATGGAGAAATTATGG | 170 |
| reverse | GTCCATCTTGGTAAGTTTCTTTTCTT | ||
| ACT1 | forward | ATTGCCGAAAGAATGCAAAAGG | 220 |
| reverse | CGCACAAAAGCAGAGATTAGAAACA |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kunikane, S.; Nakayama, T. Recent advances in plant secondary product glycosyltransferase research. Seikagaku 2008, 80, 1033–1038. [Google Scholar] [PubMed]
- Schweiger, W.; Boddu, J.; Shin, S.; Poppenberger, B.; Berthiller, F.; Lemmens, M.; Muehlbauer, G.J.; Adam, G. Validation of a candidate deoxynivalenol-inactivating UDP-glucosyltransferase from barley by heterologous expression in yeast. Mol. Plant Microbe Interact. 2010, 23, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Torres-Acosta, J.A.; Heinen, S.J.; McCormick, S.; Lemmens, M.; Paris, M.P.; Berthiller, F.; Adam, G.; Muehlbauer, G.J. Transgenic Arabidopsis thaliana expressing a barley UDP-glucosyltransferase exhibit resistance to the mycotoxin deoxynivalenol. J. Exp. Bot 2012, 63, 4731–4740. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthiller, F.; Dall’Asta, C.; Schuhmacher, R.; Lemmens, M.; Adam, G.; Krska, R. Masked mycotoxins: Determination of a deoxynivalenol glucoside in artificially and naturally contaminated wheat by liquid chromatography-tandem mass spectrometry. J. Agric. Food Chem. 2005, 53, 3421–3425. [Google Scholar] [CrossRef] [PubMed]
- Ovando-Martínez, M.; Ozsisli, B.; Anderson, J.; Whitney, K.; Ohm, J.B.; Simsek, S. Analysis of deoxynivalenol and deoxynivalenol-3-glucoside in hard red spring wheat inoculated with Fusarium graminearum. Toxins 2013, 5, 2522–2532. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Dall’asta, C.; Corradini, R.; Marchelli, R.; Sulyok, M.; Krska, R.; Adam, G.; Schuhmacher, R. Occurrence of deoxynivalenol and its 3-β-D-glucoside in wheat and maize. Food Addit. Contam. Part A 2009, 26, 507–511. [Google Scholar] [CrossRef]
- Kostelanska, M.; Hajslova, J.; Zachariasova, M.; Malachova, A.; Kalachova, K.; Poustka, J.; Fiala, J.; Scott, P.M.; Berthiller, F.; Krska, R. Occurrence of deoxynivalenol and its major conjugate, deoxynivalenol-3-glucoside, in beer and some brewing intermediates. J. Agric. Food Chem. 2009, 57, 3187–3194. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Nagl, V.; Schwartz, H.; Krska, R.; Moll, W.D.; Knasmüller, S.; Ritzmann, M.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in rats. Toxicol. Lett. 2012, 213, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.W.; Duncan, G.; Richardson, A.J. The human fecal microbiota metabolizes deoxynivalenol and deoxynivalenol-3-glucoside and may be responsible for urinary deepoxy-deoxynivalenol. Appl. Environ. Microbiol. 2013, 79, 1821–1825. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Sirisattha, S.; Mori, K.; Iwahashi, Y. Mycotoxin toxicity in Saccharomyces cerevisiae differs depending on gene mutations. Food Sci. Technol. Res. 2009, 15, 453–458. [Google Scholar] [CrossRef]
- Ghaffari, M.R.; Mardi, M.; Ehya, F.; Farsad, L.K.; Hosseini, S.; Ghareyazie, B. Mapping and expression analysis of a Fusarium head blight resistance gene candidate pleiotropic drug resistance 5 (PDR5) in wheat. Iran. J. Biotechnol. 2010, 8, 112–116. [Google Scholar]
- Sirisattha, S.; Momose, Y.; Kitagawa, E.; Iwahashi, H. Toxicity of anionic detergents determined by Saccharomyces cerevisiae microarray analysis. Water Res. 2004, 38, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Iwahashi, Y. Comprehensive gene expression analysis of type B trichothecenes. J. Agric. Food Chem. 2012, 60, 9519–9527. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.J.; McCormick, S.P.; Ziegenhorn, S.L. Phytotoxicity of selected trichothecenes using Chlamydomonas reinhardtii as a model systemt. Nat. Toxins 1999, 7, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Iwahashi, Y. Phytotoxicity evaluation of type B trichothecenes using a Chlamydomonas reinhardtii model system. Toxins 2014, 6, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Savard, M. Deoxynivalenol fatty acid and glucoside conjugates. J. Agric. Food Chem. 1991, 39, 570–574. [Google Scholar] [CrossRef]
- Poppenberger, B.; Berthiller, F.; Lucyshyn, D.; Sieberer, T.; Schuhmacher, R.; Krska, R.; Kuchler, K.; Glössl, J.; Luschnig, C.; Adam, G. Detoxification of the Fusarium mycotoxin deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 47905–47914. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Iwahashi, Y. Gene expression profile of MAP kinase PTC1 mutant exposed to Aflatoxin B1: Dysfunctions of gene expression in glucose utilization and sphingolipid metabolism. Chem-Bio Informat. J. 2009, 9, 94–107. [Google Scholar] [CrossRef]
- Yiannikouris, A.; André, G.; Poughon, L.; François, J.; Dussap, C.G.; Jeminet, G.; Bertin, G.; Jouany, J.P. Chemical and conformational study of the interactions involved in mycotoxin complexation with β-D-glucans. Biomacromolecules 2006, 7, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Fukuzawa, H.; Kubo, T.; Yamano, T. Genome of a green alga, Chlamydomonas reinhardtii, lights up key functions of plant and animal cells. (In Japanese)Tanpakushitsu Kakusan Koso 2008, 53, 1133–1143. [Google Scholar] [PubMed]
- Kam, V.; Moseyko, N.; Nemson, J.; Feldman, L.J. Gravitaxis in Chlamydomonas reinhardtii: Characterization using video microscopy and computer analysis. Int. J. Plant. Sci. 1999, 160, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; McCormick, S.P.; Appell, M. Structure-activity relationships of trichothecene toxins in an Arabidopsis thaliana leaf assay. J. Agric. Food Chem. 2007, 55, 6487–6492. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Schuhmacher, R.; Adam, G.; Krska, R. Formation, determination and significance of masked and other conjugated mycotoxins. Anal. Bioanal. Chem. 2009, 395, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Ohmichi, K.; Sakamoto, S.; Sago, Y.; Kushiro, M.; Nagashima, H.; Yoshida, M.; Nakajima, T. Detection of a new Fusarium masked mycotoxin in wheat grain by high-resolution LC-Orbitrap MS. Food Addit. Contam. Part A 2011, 28, 1447–1456. [Google Scholar] [CrossRef]
- Maragos, C.M.; Kurtzman, C.; Busman, M.; Price, N.; McCormick, S. Development and evaluation of monoclonal antibodies for the glucoside of T-2 toxin (t2-glc). Toxins 2013, 5, 1299–1313. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.H. A comprehensive guide to biology and laboratory use. In The Chlamydomonas Sourcebook; Academic Press: San Diego, CA, USA, 1989; pp. 1503–1504. [Google Scholar]
- Iwahashi, H.; Odani, M.; Ishidou, E.; Kitagawa, E. Adaptation of Saccharomyces cerevisiae to high hydrostatic pressure causing growth inhibition. FEBS Lett. 2005, 579, 2847–2852. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, T.; Iwahashi, Y. Low Toxicity of Deoxynivalenol-3-Glucoside in Microbial Cells. Toxins 2015, 7, 187-200. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7010187
Suzuki T, Iwahashi Y. Low Toxicity of Deoxynivalenol-3-Glucoside in Microbial Cells. Toxins. 2015; 7(1):187-200. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7010187
Chicago/Turabian StyleSuzuki, Tadahiro, and Yumiko Iwahashi. 2015. "Low Toxicity of Deoxynivalenol-3-Glucoside in Microbial Cells" Toxins 7, no. 1: 187-200. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7010187